
Abstract
Genetically modified lymphoblastoid cell lines (LCL) have been shown to be an attractive alternative source of antigen-presenting cells for cancer vaccination in vitro. We tested their application in patients with pancreatic cancer in a phase I clinical trial. As a model tumor antigen, we selected the point-mutated (codon 12) Ki-Ras p21 oncogene (muRas) frequently (∼85%) present in pancreatic adenocarcinoma. Autologous LCLs were established in vitro by spontaneous outgrowth from peripheral blood lymphocytes of seven pancreatic carcinoma patients and were genetically modified with an episomal Epstein-Barr virus (EBV)–based expression vector to express muRas (muRas-LCL). Weekly vaccinations with subcutaneous injection of 5×106 muRas-LCL were done. In six of seven patients, therapeutic vaccination elicited a T-cell response with an increase in the frequency of muRas-specific precursor cytotoxic T lymphocytes in the peripheral blood and positive delayed-type hypersensitivity reactions at the injection site. Besides local reactions and flu-like symptoms, there were no signs of toxicity and no acute EBV infection, onset of EBV-associated lymphoma, or other severe complications. A clinical response (stable disease) was observed for a short time period (2–4 months) in four of seven patients (57%), mostly in earlier tumor stages. Our results indicate that LCL presenting genetically modified antigen represent a valuable and easily available tool for in vivo autologous tumor vaccination. LCL can be transfected with any known tumor antigen and therefore should be further clinically investigated.
Introduction
A promising vaccine strategy is the immunization with antigen-presenting cells (APCs), such as dendritic cells (DCs) pulsed ex vivo with peptides from tumor antigens (Melief, 2008) or genetically modified to express tumor antigens (Ribas, 2005), because such APCs are potent inducers of anti-tumor immunity (Palucka et al., 2011). However, these cells are available in limited numbers in cancer patients (Thurner et al., 1999), and it is not entirely clear which type and activation status of the DC (e.g. CD34- or monocyte-derived DC, cytokines used for DC generation) is best suited for vaccination (Tacken et al., 2007). In addition, it has been reported that multiple applications of DCs may decrease anti-tumor immunity (Ribas et al., 2000).
Lymphoblastoid cell lines (SP-LCL) can be considered an attractive alternative source of antigen-presenting cells for cancer vaccination (Pecher and Finn, 1996; Kubuschok et al., 2000; Kubuschok et al., 2002), but so far have not been applied to human subjects (Moore et al., 1957). Like dendritic cells, SP-LCLs are “professional” APCs and have a high immunostimulatory capacity (Bangham and McMichael, 1986; Martin et al., 1993; Sun et al., 1999). They can be easily obtained by spontaneous outgrowth from Epstein-Barr virus (EBV)–transformed B cells in cyclosporine-containing medium from EBV-positive patients (Bird et al., 1981) without adding exogenous EBV virus. Because of their rapid and long-lasting growth in vitro, they are a potentially unlimited source of APCs. Therefore, we established genetically modified SP-LCL to express the model tumor antigen-mutated ras (muRas-LCL) (Kubuschok et al., 2000) and investigated their practicability as a cancer vaccine in vitro in pancreatic cancer patients who expressed this antigen in their primary tumors (Kubuschok et al., 2002).
We demonstrated that such genetically modified SP-LCL can efficiently present the tumor antigen to the immune system and induce antigen-specific cytotoxic T cells, which are able to kill tumor antigen positive tumor cells. We selected the point-mutated Ki-Ras p21 oncogene as a model tumor antigen, because many human cancers harbor point mutations in the ras gene at codon 12, and these cancers are difficult to treat by current chemoimmunotherapy regimens (Bardelli and Siena, 2010). In pancreatic adenocarcinoma, mutated ras (muRas) at codon 12 occurs in 80–90% of cases. In contrast to other mutated tumor-specific proteins (p.e. p53), mutations at codon 12 are almost exclusively restricted to four common amino acid substitutions, namely the replacement of the normal glycine (Gly) residue by either an aspartic acid (Asp), valine (Val), cysteine (Cys), or arginine (Arg), and can therefore be covered by four vaccine formulations. We also established a method suitable for immunomonitoring of T-cell responses against mutated p21 ras protein and showed that CD8+ as well as CD4+ T-cell immunity to muRas is pre-existent in a considerable proportion of patients with pancreatic carcinoma. This demonstrates the presence of a naturally occurring T-cell response in pancreatic cancer, which can be exploited for immunostimulatory approaches (Kubuschok et al., 2006).
On the basis of this experimental background we now present the in vivo results of a phase I study in seven patients with locally advanced pancreatic carcinoma vaccinated with muRas-transfected LCL. The objectives of this study were (a) to determine the safety and toxicity of the vaccine; (b) to determine the cellular immune response against mutated ras before and after vaccination (number of CD4+/CD8+ T cells and their cytotoxic activity); and (c) to determine the immunological response against EBV before and after vaccination. (d) We also sought to gain preliminary evidence for a potential clinical response.
Materials and Methods
Patients
This study was approved by the local ethical review board (Ethikkommission der Ärztekammer des Saarlandes) and performed according to the Declaration of Helsinki. The trial is registered at the German Registry for Somatic Gene-Transfer Trials (DeReG-Nr.1001). Patients were eligible for enrollment if they had relapsed or newly diagnosed locally advanced, non-resectable muRas-positive pancreatic cancer, a measurable tumor parameter (e.g., CA-19-9), seropositivity for EBV virus, a Karnofsky index >50%, an estimated life expectancy ≥4 months, signed informed consent, and were ≥6 weeks from chemotherapy, radiotherapy, or immunotherapy. Patients were excluded if they had a severe concomitant disease or organ dysfunction not caused by the pancreatic carcinoma.
Tumor tissues and peripheral blood mononuclear cells
Tumor tissues and peripheral blood mononuclear cells (PBMC) were obtained from 72 patients with operable adenocarcinoma of the pancreas at the University of Saarland Medical School (Homburg, Germany) between 1997 and 2000. Thirty-five patients were shown by PCR to bear one of the most common Ki-ras mutations at amino acid position 12 (valine, aspartic acid, or arginine replacing glycine) in a sample of their primary tumor. Peripheral blood was taken 7 days after surgical resection, and PBMC were isolated and human leukocyte antigen (HLA)-A2 typed as previously described (Kubuschok et al., 2006). PBMC samples were used for the production of the muRas vaccine and other PBMC samples for T-cell assays were collected before and after vaccinations.
Vaccine preparation
SP-LCL were established by spontaneous outgrowth of peripheral blood B lymphocytes from PBMC samples in the presence of 0.1 μg/ml cyclosporine A (Novartis, Basel, Switzerland), as previously described (Kubuschok et al., 2000). A DNA fragment encoding the amino acids 1-25 of wild-type Ki-ras, or Ki-ras mutated at codon 12, was cloned into the HindIII/BamH1 sites of the expression vector pCEP4 (Invitrogen, Leek, Netherlands), resulting in pCEP4-muRas/1-25Codon12. Plasmids pCEP4-muRas/1-25Val12, pCEP4-muRas/1-25Aspl12, or pCEP4-muRas/1-25Argl12 were transfected by electroporation into SP-LCLs established from pancreatic cancer patients, as previously described (Kubuschok et al., 2000). Recombinant DNA work was done under good medical practice (GMP) conditions with the official permission and according to the rules of the State Government of Saarland.
Phenotype of mu-Ras-LCL
SP-LCL before and after transfection were tested for the expression of MHC-class-I and II molecules (HLA-A2, HLA-DR) and the costimulatory molecules B7.1, ICAM-1, and LFA-3 by direct or indirect immunofluorescence and analyzed by FACScan (Becton-Dickinson, Heidelberg, Germany) equipped with Lysis II software, as described earlier (Kubuschok et al., 2000).
Cytokine production of mu-Ras-LCL
SP-LCL were transfected with plasmids pCEP4-blank (without insert) and pCEP4-muRas/1-25Val12, respectively, as described above. For real-time polymerase chain reaction (RT-PCR) and ELISA, continuously growing cell cultures with at least 60% vital cells were used for expression analysis. At least two different transfections were done with three different LCLs. Messenger RNA (mRNA) expression was determined by real-time reverse transcriptase PCR for the cytokines IL-6, -8, -10, TNF-α, MIP-1α, and MIP-1β according to the protocol described by Bettink et al. (2010). For this purpose, mRNA derived from 3×106 gene-modified LCL was isolated (RNeasy Mini Kit; Quiagen, Hilden, Germany) and transcribed into cDNA using SuperScript™ II Reverse Transcriptase (Invitrogen GmbH, Karlsruhe, Germany). Real-time PCR was done using TaqMan technology (StepOnePlus™ Real-Time PSR System; Applied Biosystems, Darmstadt, Germany). The following (5’-3’)-primers were used: IL-6 (forward—ATGCAATAACCACCCCTGAC, backward—GAGGTGCCCATGCTACATTT); IL-8 (forward—CTGGCCGTGGCTCTC TTG, backward—CCTTGGCAAAACTGCACCTT); IL-10 (forward—GGTGATGCCCCA AGCTGA, backward—TCCCCCAGGGAGTTC ACA); TNF-α (forward—ATGAGCACTG AAAGCATGATCC, backward—GAGGGCTGATTAGAGAGAGGT); MIP-1α (forward—AGCTGACTACTTTGAGACGAGCAG, backward—CGGCTTCGCTTGGTTAGGA); MIP-1β (forward—CTGCTCTCCAGCGCTCTCA, backward—GTAAGAAAAGCAGC AGGCGG). All data were normalized based on their expression of 18s-RNA as housekeeping standard. Protein expression was detected for the cytokines IL-6, -8, -10, and MIP-1α. For this purpose, the supernatant of the LCL cultures was analyzed by ELISA in strict adherence to the manufacturer's instructions (R&D Systems, Wiesbaden, Germany). Protein expression levels were normalized to the number of cells and the cell culture volume of each LCL.
Treatment schedule
For immunization, 5×106 muRas-LCL were subcutaneously injected alternately into the right and left thighs for a total of initially 4 weekly vaccinations. In the case of an increase in muRas-specific T-cell response, vaccinations were paused for 8 weeks before the next four immunizations. In nonresponding patients, vaccinations were continued up to the generation of a significant muRas-specific T-cell response, including at least four additional vaccinations.
Evaluation of specific T-cell response by interferon-γ, IL-4, and IL-10 ELISPOT
The frequency of precursor cytotoxic T cells (pCTL) was measured before and weekly after every immunization by IFN-γ-Elispot. PBMC were stimulated with the relevant HLA-A2*0201–restricted muRas peptides and viral control peptides (influenza matrix protein, BMLF1, and LMP2). A rise in pCTL frequency was defined as a minimum two-fold increase of pCTL frequency in comparison to the value before vaccination. As described earlier (Kubuschok et al., 2006), the interferon-γ, IL-4, and IL-10 ELISPOT were performed in multiscreen filtration plates (MAHAN 4550; Millipore, Bedford, MA), and T-cell subpopulations were isolated by CD4-/CD8-antibody-coated magnetic microbeads from PBMC (Dynal, Oslo, Norway). ELISPOT assays were performed according to manufacturer's instructions (Gen-Probe Diaclone SAS; Besancon, Cedex, France).
Evaluation of specific T-cell responses by intracellular interferon-γ FACS analysis
Antigen stimulations with muRas, wild-type ras, or virus peptides were performed in PBMC, which were analyzed by intracellular flow cytometry as described earlier (Kubuschok et al., 2006). As negative controls, PBMCs were incubated without peptide and with control peptide HIV Pol476–484.
Cytotoxic function of vaccine-generated muRas-specific T cells
PBMC were stimulated by autologous 100 Gy-irradiated muRas-LCL every week. After 4 weeks, standard chromium release assays were done as previously described (Kubuschok et al., 2006). Target cell lines Patu, 8988S, and SW480 were provided as described (Kubuschok et al., 2006) and were retested for muRas and HLA-A2 expression 2 weeks before the assays (Kubuschok et al., 2006).
Synthetic peptides for T-cell stimulation
The quality and sequences of peptides derived from mutated Ras (muRas [Val12] or [Asp12]5–21 or 5–14) and viral proteins (IMP58–66, HIV-Pol476–484, EBV-BMLF-1280–288, EBV-LMP2426–434) have been reported earlier (Kubuschok et al., 2006).
Determination of delayed-type hypersensitivity reaction
Delayed-type hypersensitivity (DTH) responses (erythema and induration) against muRas-LCL were measured 24, 48, and 72 hr after the vaccination and were considered positive if the diameter of the skin reaction was >5 mm after 48 h. Mock-vector-LCL in protein-free medium was injected at least 10 cm away from the muRas vaccine as a negative control. Punch biopsies (6 mm) were obtained from positive DTH sites and evaluated immunohistochemically for activated CD4+ and CD8+ T lymphocytes (CD4, CD8, and CD45RA) by alkaline phosphatase-anti-alkaline phosphatase (APAAP) technique according to the manufacturer's recommendations (Dako-Cytomation, Glostrup, Denmark). As a control for the proper function of the DTH response, we tested the delayed-type hypersensitivity reaction (DTH-reaction) for selected recall antigens (tuberculin, tetanus toxoid, diphtheria toxoid, streptococcal antigen, trichophyton antigen, candida antigen, and proteus antigen) with a commercially available test stamp (Immignost Multitest; Bioysn, Felbach, Germany) before the first vaccination and 2 weeks after the last vaccination.
Safety monitoring during treatment
To detect possible side effects we determined total blood count, differential count, plasma levels of electrolytes, protein, albumin, creatinine kinase, glucose, amylase, lipase, and liver and renal function weekly. To exclude acute EBV-infection, serum antibodies against EBV antigens and EBV DNA were determined weekly during vaccination. Adverse events reported were coded according to the World Health Organization's (WHO's) common toxicity criteria (Trotti et al., 2003).
Statistical analysis
Statistical analysis was done as described earlier (Kubuschok et al., 2002). All data for assessment of cytokine production by RT-PCR and ELISA were acquired in two independent experiments. Assay samples were run in duplicates. Variance was calculated using the F-test. Significance was calculated using Student's T-test.
Results
Patients' characteristics
Of 35 patients screened, we identified 12 HLA-A2 positive (indispensable for immunomonitoring) patients with a Ki-Ras codon 12 mutation. Three patients were excluded because of the distance between their homes and the treatment facility, and two patients did not give informed consent. Seven patients with relapsed or metastatic pancreatic adenocarcinoma were enrolled in this pilot study. These patients' characteristics are shown in Table 1. There were four males and three females with a mean age of 59.1 years (range, 48 to 66 years). Four patients had been treated by pancreaticoduodenectomy (Whipple or Traverso operation) and subsequent adjuvant chemotherapy (aCH), two patients were a priori inoperable and were directly subjected to palliative chemotherapy (pCH), and one patient underwent palliative chemotherapy when relapse occurred 14 months after pancreaticoduodenectomy (patient 2). All tumors were classified as adenocarcinomas, moderately differentiated in most cases (6/7), with the exception of one poorly differentiated carcinoma (pat 5). At the time of diagnosis, three patients had stage I disease (pT1-2, N0, M0), according to the WHO classification (Sobin et al., 2009), two patients suffered from locally advanced pancreatic cancer equivalent with stage II or III (pT1-3, N0-1, M0), and two patients had metastatic disease (Stage IV). The time to relapse after operation combined with adjuvant therapy, or after first-line chemotherapy, ranged from 4 to 26 months, demonstrating differences in the dynamics of disease progression (Table 1).
Traverso operation+radiotherapy.
Palliative gastrojejunostomy.
Time to progression (time from start of therapy until objective tumor progression) after 1st or 2nd therapy before vaccination in months, this time is identical with the time vaccination starts after the last therapy.
Gemcitabine/5-fluorouracil.
Ag, antigen; m, male; f, female; OP, Whipple-OP; Inop, inoperable, aCH, adjuvant chemotherapy (gemcitabine); noCH, no adjuvant chemotherapy; pCH, palliative chemotherapy (gemcitabine); n, not determined.
Immunological status before vaccination
In order to determine the general T-cell responsiveness in the treated patients, we tested for DTH-reactions for selected recall antigens. In all, except one patient, a normal reaction against recall antigens was found (Table 1). The muRas-specific CD8+ T-cell response was also investigated before the start of vaccination to collect information about the basic level of muRas-specific T-cell response with respect to later monitoring of the muRas T-cell response during therapy.
Phenotype, cytokine production, and function of muRas-LCL
In order to control the quality of the vaccine before application, we demonstrated the expression of muRas oligonucleotides in LCL transfected with pCEP-muRas/1-25 by immunofluorescence staining and Western blot using a monoclonal antibody against muRas. We also demonstrated the expression of MHC class I and II molecules (HLA-A,B,C and HLA-DR) as well as important immunoregulatory molecules (CD54, CD56, CD80, CD86) on the surface of these antigen-presenting LCL (Kubuschok et al., 2000). Moreover, we analyzed the cytokine profile secreted by LCL before and after transfection to explore whether the production of immunosuppressive or immunostimulatory cytokines is turned on in muRas-transfected LCL. According to previously published data about the cytokine profile of LCL, we focused on five cytokines, which are regularly expressed in LCL and may influence the immunostimulatory capacity of LCL (IL-8, IL-10, TNF-α, MIP-1α/CCL3, and MIP-1β/CCL4) (Miyauchi et al., 2011). We also investigated the secretion of IL-6, which is in addition to IL-8, upregulated by oncogenic ras signals in cancer cells (Ancrile et al., 2008), and therefore may also be upregulated by transfection of muRas oligonucleotides in LCL. To this end, the expression of these six cytokines (rather immunosuppressive: IL-6, IL-8, IL-10, MIP-1α, MIP-1β; rather immunostimulatory: TNF-α) was determined in different LCL by real-time PCR (data not shown).
For transfection experiments, we chose the three LCL (LCL #1, #2, and #3), which expressed these six cytokines most strongly. These three LCL were transfected as described above, so that three sublines were derived from each LCL: nontransfected (1), transfected with the control-plasmid pCEP4-blank (2), and transfected with pCEP4-muRas/1-25 (3). In these sublines, the mRNA transcription of the six cytokines was investigated by real-time PCR, and additionally protein expression was examined by ELISA in the LCL cell culture supernatant, for the cytokines with the strongest PCR signals (IL-6, IL-8, IL-10, MIP-1β). Overall, we noted that the cytokine levels varied considerably within a given LCL as well as between LCL from different subjects. This was the case for nontransfected LCL as well as for transfected LCL.
Concerning the impact of transfection, stably transfected LCL generally expressed or produced lower amounts of cytokines than nontransfected cell lines. As an example, the results of LCL #3 are shown (Fig. 1a and b). mRNA transcription as determined by RT-PCR, with a few exceptions, paralleled the cytokine secretion as determined by ELISA (Fig. 1a and b). Figure 1c shows the relative attenuation of expression of the four cytokines of the three LCL after transfection (all experiments were repeated at least twice). Notably, there was no significant difference between the decrease caused by transfection with pCEP4-blank and the one caused by pCEP4-muRas. This indicates that muRas-peptides encoded by pCEP4-muRas (p1-25) had no vector-independent impact on the expression of the cytokines under investigation, including the ras-regulated IL-6 and IL-8.
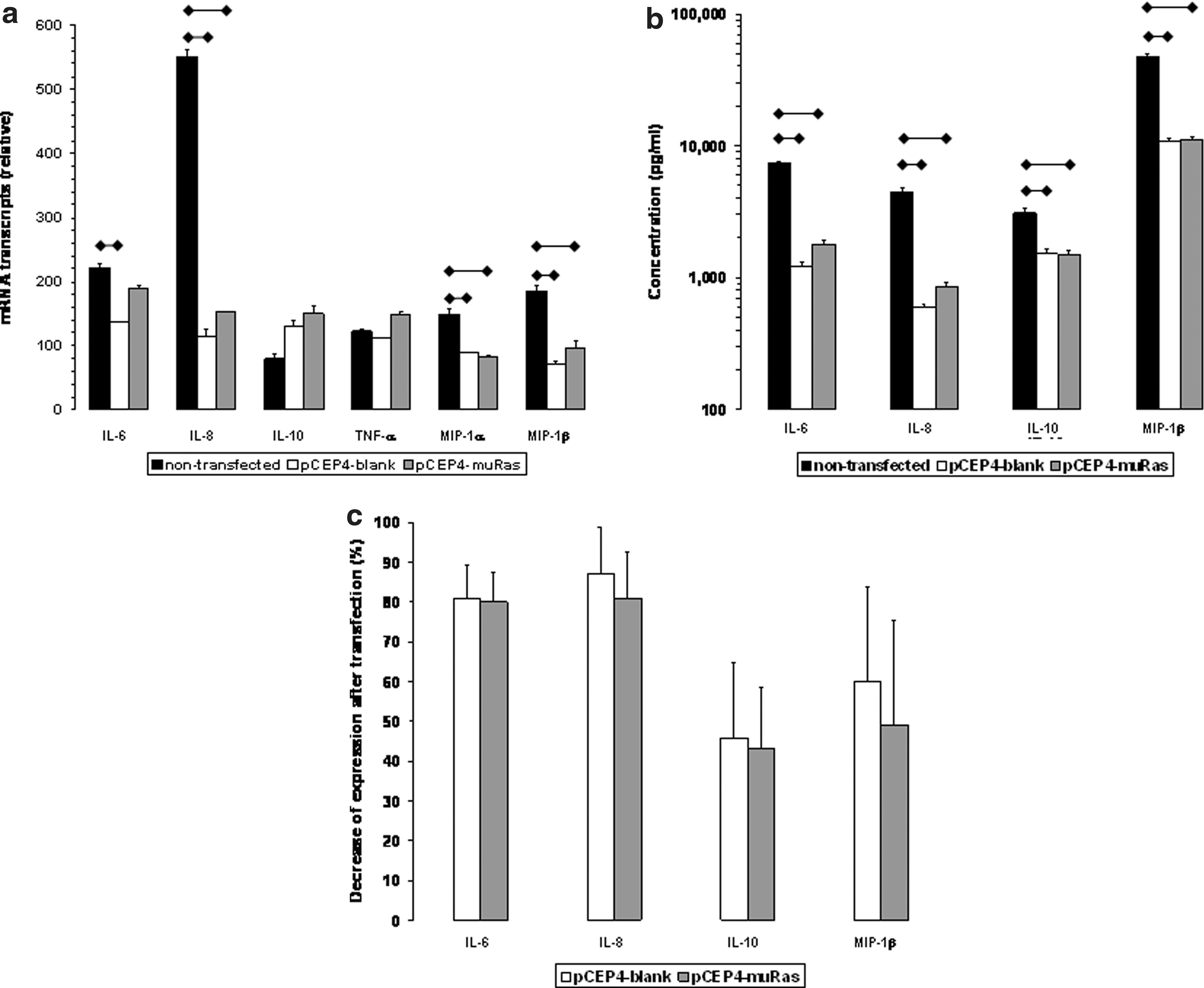
In addition, these results indicate that either genetic components of the vector other than muRas oligonucleotides may cause the decrease of expression or that components of the transfection procedure itself are responsible for downregulation of cytokines. As we had demonstrated earlier (Kubuschok et al., 2002), the in vitro capacity of muRas LCL to elicit a muRas-specific cytotoxic T-cell response against pancreatic tumor cells was strong. This supports the view that the immunostimulatory capacity of muRas LCL is not seriously affected by the lower expression of cytokines.
Peripheral blood T-cell response against muRas vaccine
Six of seven patients demonstrated an increase of muRas-specific CD8+/CD4+ T-lymphocytes in the peripheral blood after vaccination with muRas-LCL (Table 2) measured against the 17-mer peptide derived from muRas in ELISPOT assay. This result was confirmed by assessing the CD8+ T-cell response against the 9mer peptide 5-14 in IFN-γ as determined by flow cytometry (Table 2). In contrast, in patient #6 there was no increase of the muRas-specific T-lymphocytes during the vaccination (Fig. 2). Corresponding to this observation, no DTH response was observed at vaccine injections sites in this patient. This patient had no muRas-specific T cells before vaccination and also did not react to recall antigens before vaccination, indicating a status of immunological anergy. At the start of treatment this patient had metastatic pancreatic carcinoma with a high tumor burden as documented by peritoneal carcinosis and a high value of CA19-9 (22,800 U/ml; Fig. 2), which progressed during the vaccination period.
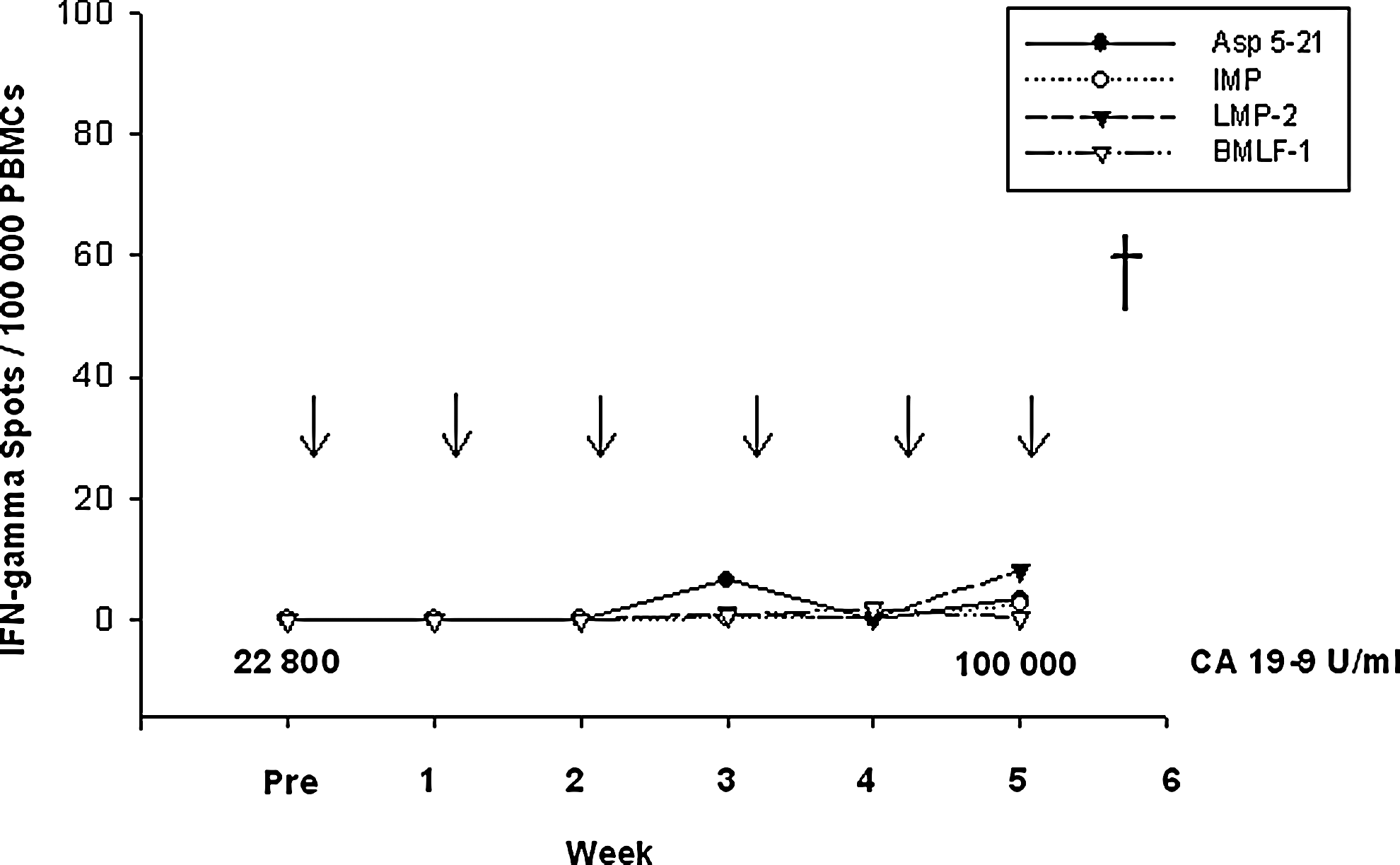
Nonresponse to vaccination with muRasAsp12-LCL. In patient #6, no T-cell response against muRasAsp12 5-21 or viral peptides (IMP, LMP-2, or BMLF-1) was detectable before or after five vaccinations. Pre: before vaccination; ↓: vaccination; †: death.
muRas target peptide 5-21 (17mer).
muRas target peptide 5-14 (9mer).
Stable or progressive disease according to Response Evaluation Criteria in Solid Tumors (RECIST).
In contrast to this patient, a significant increase in the frequency of antigen-experienced T cells, as well as a DTH reaction after vaccination, was observed in the other six patients vaccinated with muRas-LCL (Table 2). For example, patient #1 underwent R0-Whipple operation and adjuvant chemotherapy. Twenty-six months after this initial treatment, increasing levels of CA-19-9 (measurement error +/− 1 U/ml) were observed without radiological signs for local relapse or metastases in CT scans. There was no measurable T-cell response against muRas at this time point. A first set of four vaccinations was applied and elicited a muRas-specific T-cell response up to 45 IFN-γ–secreting cells per 100,000 PBMCs measured by ELISPOT assay (Fig. 3a). This maximal response was observed 1 week after the fourth vaccination. Simultaneously, CA-19-9 levels decreased and therapy was paused.

Two months later, CA-19-9 levels increased again and a second course of four vaccinations was applied, followed by another vaccination break. Again, a decline in CA-19-9 levels was observed. When CA-19-9 levels increased after another month, and three small liver metastases were detected by abdominal CT scan, a third set of vaccinations was performed and continued during stable disease without a break. It was stopped after 13 vaccinations of the third cycle because liver metastases and CA-19-9 levels showed progression. Interestingly, the analysis of biopsy material derived from these liver metastases did not reveal the presence of the initially detected K-ras mutation. This suggests that a clone of tumor cells without ras mutation was the origin of the metastasis and that the development of this wild-type clone was responsible for the escape of this tumor from immunotherapy.
Two patients (#3 and #4) had a naturally occurring muRas response prior to vaccination. In patient #3, local relapse was detected by CT scan 13 months after pancreaticoduodenectomy, and a CA-19-9 level of 440 U/ml was measured (Fig. 3b). After four cycles of vaccination an only modest muRas-specific T-cell response was observed and therapy was continued without interruption. After eight cycles of weekly vaccinations there was a strong increase in muRas-specific T cells. However, CA 19-9 levels had risen again, and the size of the relapsed tumor had slightly increased. Vaccination was continued longer than initially planned (in two weekly intervals), but muRas-specific T-cell responses decreased in the course of treatment and the vaccination was stopped when further increase in CA-19-9 levels and clinical deterioration was noticed. This case shows that the number of vaccinations necessary to elicit a strong T-cell response varied between patients.
In order to compare the muRas-specific immune response against muRas-LCL with that of an antigen not relevant for the vaccine, we measured the immune response against cytomegalovirus (CMV; data not shown) and inosine 5-monophosphate (IMP) in all treated patients. It was shown that these responses were stable under vaccination with muRas-LCL in comparison to the muRas-specific response (Fig. 2 and Fig. 3a and b). Therefore, it seems to be unlikely that the boost of the muRas-specific T-cell response is caused by a general unspecific activation of the immune system. If this were the case, the T-cell responses against CMV or IMP should have risen similarly under vaccination. The only increase of IMP-specific T cells was found in a patient who suffered from a rhinobronchial infection one week before the IMP-specific T cells were measured, which suggests an influenza infection (Fig. 3a, Week 2).
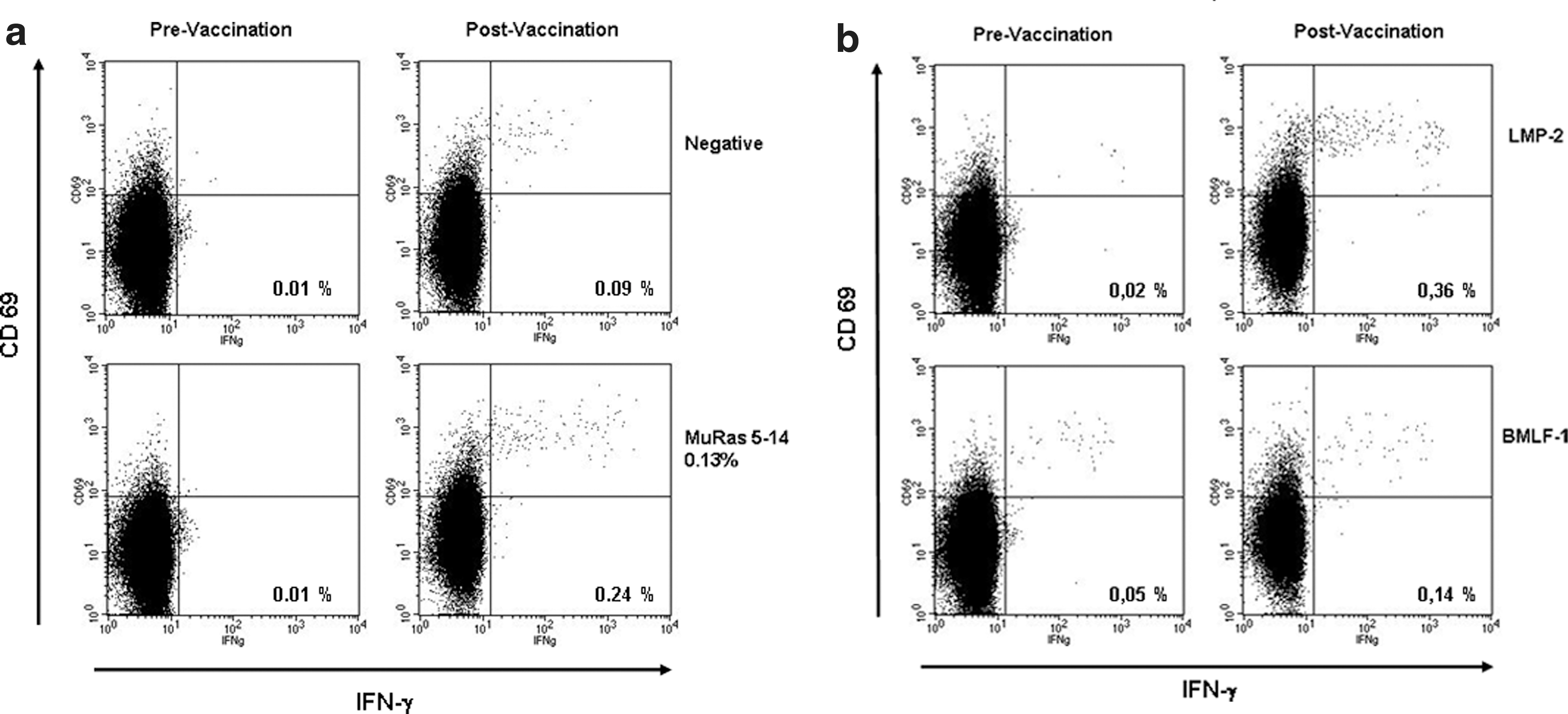
Finally we investigated the T-cell response against two EBV antigens in order to get an impression of the immunological response against the EBV-lymphoblastoid component of the vaccine. We observed an enhancement of the immune response against the latent membrane protein 2 (LMP2) nonamer after five vaccinations in four of seven patients (Table 2, Fig. 3b, and Fig. 5b), whereas the remaining three did not respond to LMP2 (Fig. 2 and Fig. 3a). An increase of the T-cell response against EBV antigen BamHIM leftward reading frame-1 (BMLF-1), which is expressed in B lymphocytes in the acute phase of EBV infection, was not or only weakly detected (Fig. 2, Fig. 3a and b, and Fig. 5b).
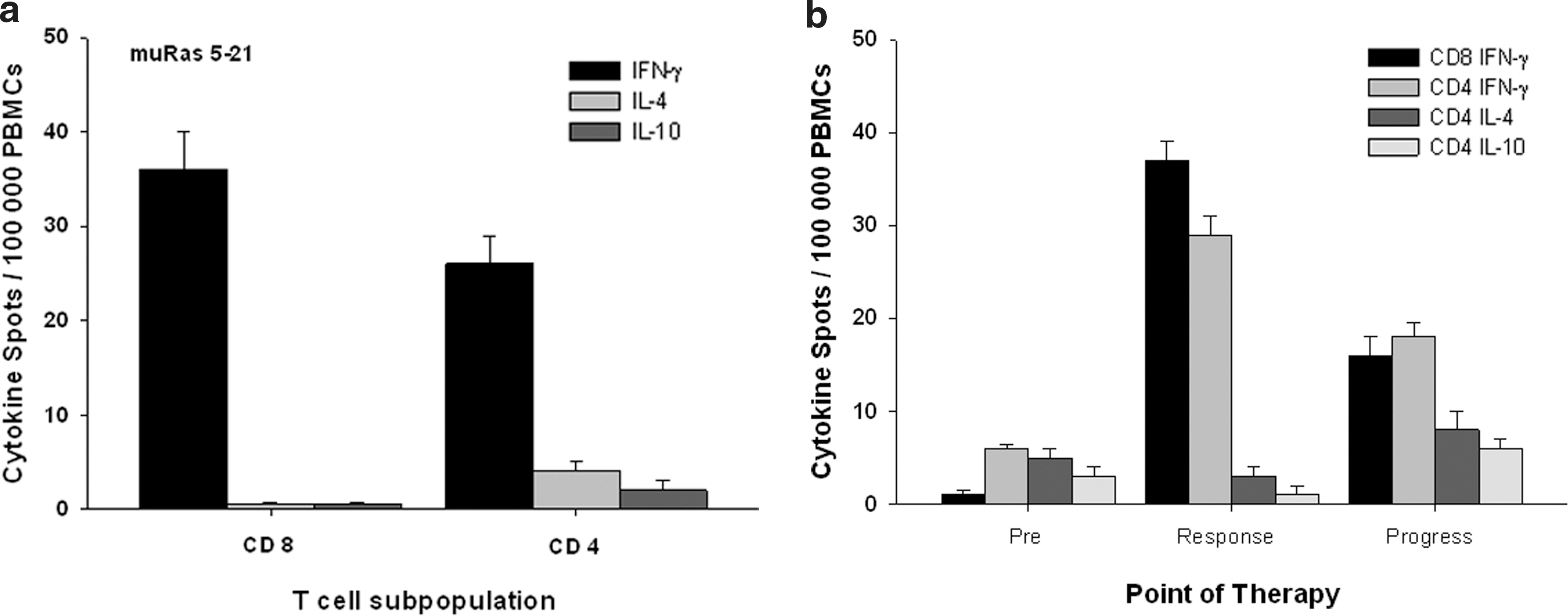
In a subgroup of patients (n=4), the T-cell response against muRas was analyzed according to the proportion of muRas-specific CD8+ and CD4+ T cells. This analysis revealed that the observed response was mediated by both CD8+ and CD4+ interferon-gamma–producing cells (Fig. 4a). In three of four cases, the CD8+ T-cell response was considerably stronger than the CD4+ T-cell response. The muRas-specific CD8+ T cells did not secrete the type-2 cytokines IL-4 or IL-10 (Fig. 4a), confirming the T-cytotoxic 1 (Tc1) type of the cytotoxic T lymphocytes and excluding a significant amount of Tc2 type or regulatory CD8+ T cells (Iezzi et al., 2006; Wang, 2008). In addition, very few muRas-specific CD4+ T cells produced IL-4 or IL-10 (Fig. 4a). This demonstrates that the CD4+ T-cell response is dominated by T cells producing T-helper 1 cytokines (IFN-γ), but not by T cells secreting cytokines typical for the T-helper 2 phenotype (IL-4, IL10) or other T cells producing IL-10 (regulatory T cells). Interestingly, we could demonstrate that in case of progressing cancer under vaccination the T-cell immune response became weaker and the CD4+/CD8+ T-cell ratio was dominated by CD4+ T cells. The analysis of the cytokine profile of these CD4+ T cells revealed a higher amount of IL-4–producing CD4+ T cells in comparison to the first weeks of the T-cell response to the vaccine (Fig. 4b).
Function of the peripheral blood T-cell response against muRas vaccine
In order to evaluate the function of muRas5–21-specific CD8+ cells, we tested the cytotoxic activity of these T cells after weekly stimulation with muRas-loaded PBMC over a period of 4 weeks. CD8+ T cells stimulated in this way killed muRas6–14- and muRas5–14-loaded T2 cells but not empty T2 cells or T2 cells loaded with control peptide (HIV-pol) in six of seven patients (Table 2). These CD8+ T cells also showed cytotoxic effects against HLA-A2-positive but not HLA-A2-negative pancreatic carcinoma cell lines (Fig. 6). These results indicate that muRas-specific cytotoxic T lymphocytes can be generated in vivo from naturally occurring muRas6–14-and muRas5-14-specific pCTL cells after muRas vaccination.

Generation of muRas-specific T-cell responses against pancreatic carcinoma cell lines after muRas-LCL-vaccination. MuRas (Val12)-specific CD8-positive T-cell lines derived from vaccinated patients (n=6) killed HLA-A0201-positive, Val12-positive pancreatic carcinoma cells (Patu), or colon carcinoma cells (SW480), but not the HLA-A0201-negative, Val12-positive cell line S8988. For efficient lysis, pre-treatment of all tumor cell lines with IFN-γ (100U/ml) was required. Lysis of tumor cells was inhibited by an anti-HLA-A2 monoclonal antibody demonstrating the HLA-A0201-restriction of the cytotoxic effect.
Immunological response measured by DTH
The injection of muRas-LCL elicited a skin reaction with erythema and induration at the injection site in six of seven patients (Tables 2 and 3 and Fig. 7a). In most cases, the skin reaction developed after the first or second vaccination, and the strength of the reaction increased after the first three to four vaccinations, but decreased thereafter (Fig. 7e). The immunohistochemical investigation of the injection sites demonstrated an infiltration with activated (CD45-positive; Fig. 7c) CD4+ and CD8+ T lymphocytes with a preponderance of CD4+ T cells (Fig. 7b and d). Altogether the observations are in accordance with a DTH-reaction. In most cases, a positive DTH response was associated with a systemic muRas T-cell response in the peripheral blood as illustrated by data assessed in a representative patient (Fig. 7e). However, in comparison to the presence of muRas-specific T cells in the peripheral blood, the DTH reaction was a less reliable predictor of the clinical response to therapeutic vaccination, since two of four patients who had stable disease after 2 months lost their DTH response after the third vaccination, despite clinical response (Fig. 7f), whereas all patients with stable disease demonstrated a steady elevation of muRas-specific T cells.
Traverso operation+radiotherapy; bpalliative gastrojejunostomy; cin months; dpositive blood T-cell response is defined as minimum two-fold increase of the pCTL frequency (measured by ELISPOT) in comparison to the value before vaccination; epositive DTH reaction is defined as >5mm after 48 hr; ffor details see Table 4; gXiong et al., 2008; hJacobs et al., 2004; iNeoptolemos et al., 2010; j Neoptolemos et al., 2004; kMoore et al., 2007.
OP, Whipple-OP; Inop, inoperable; aCH, adjuvant chemotherapy (gemcitabine); noCH, no adjuvant chemotherapy; pCH, palliative chemotherapy (gemcitabine); No., number; PFS, progression free survival defined as the time from start of therapy until objective tumor progression or death; OS, overall survival; DTH, delayed-type hypersensitivity; SD, stable disease; PD, progressive disease; ↓↑=, decrease, increase, and no change of serum CA19-9 respectively.
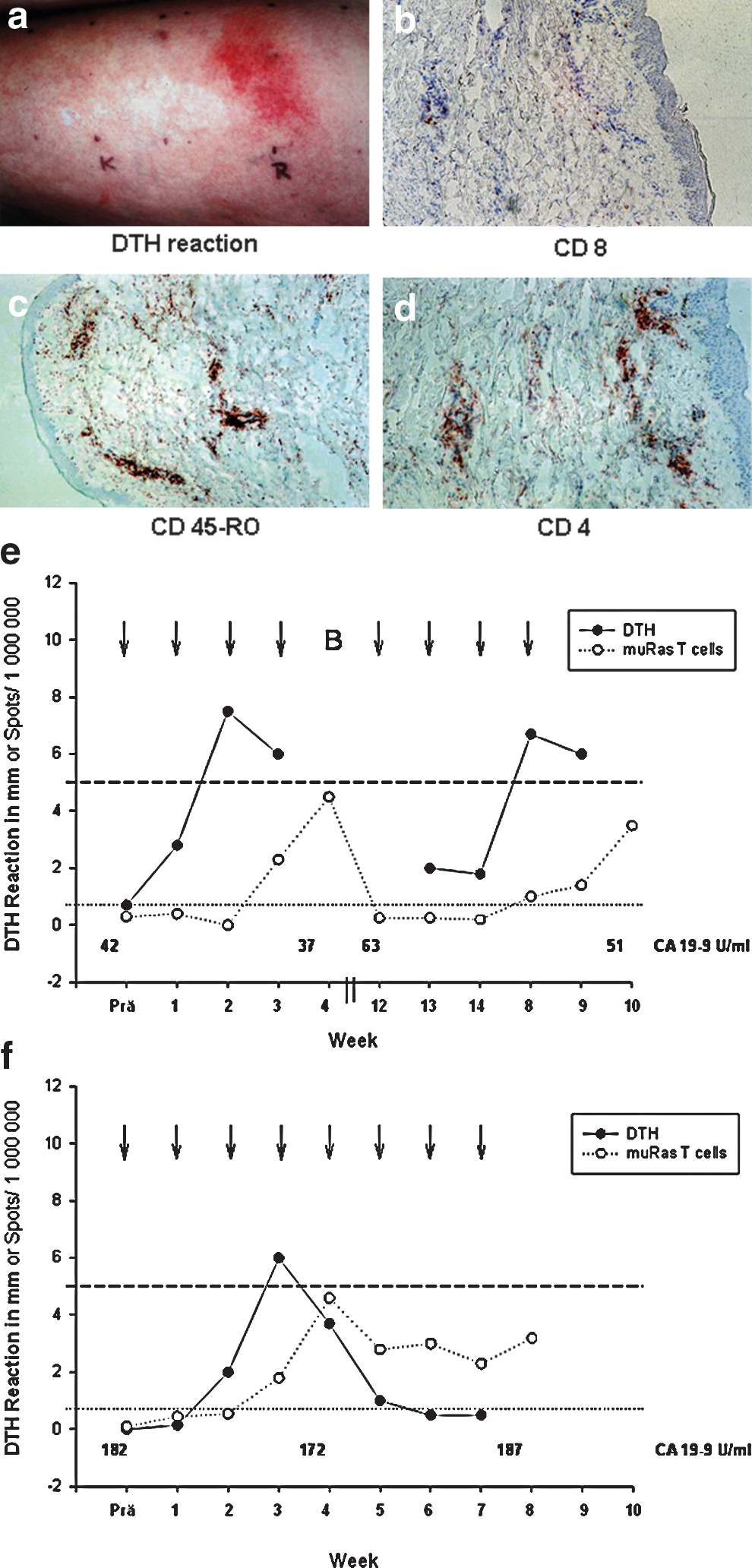
Delayed-type hypersensitivity (DTH) reaction after subcutaneous application of muRas-LCL.
Clinical response
Follow-up investigations included abdominal MRI and assessment of CA-19-9 value every 2 months. After 2 months, stable disease was found in four of seven patients (57%), according to Response Evaluation Criteria in Solid Tumors (RECIST) (Therasse et al., 2000). After 4 months of follow-up, all patients had a progressive disease (Table 2), resulting in a median progression-free survival (PFS) of 3.1 months. According to tumor stage, all patients who achieved a stable response had no lymph node metastases (Table 3). This indicates that vaccination is more effective in earlier tumor stages; although, this conclusion must be drawn with caution, because it could also be an effect of the natural course of the disease.
PFS after vaccination and overall survival (OS) after vaccination for the individual patient are shown in Table 3. For comparison, the median PFS and OS for a group of patients without vaccination but physicians' best choice of treatment (including second or third line chemotherapy) matched for TNM stage, first line therapy, and time to progression after first line therapy are listed in Table 3 for each patient (Jacobs et al., 2004; Neoptolemos et al., 2004, 2010; Moore et al., 2007; Xiong et al., 2008). Patients with vaccination may have had a slightly longer PFS and OS than the average patients, as well as the lymph node–positive cases, but vaccinated patients with an inoperable tumor had a similar PFS and an even worse survival than the average.
Because PFS and OS after vaccination were not the primary endpoints of the study, extensive conclusions cannot be drawn. However, it is remarkable that the vaccine-responding patient #1 had a favorable outcome after vaccination with an overall survival of 26 months. The latter observations and conclusions are also supported by the evaluation of the overall survival after diagnosis, which indicates longer times for OS after vaccination with the caveats of the above-mentioned restrictions.
Toxicity profile and feasibility of vaccine generation
Vaccination with muRas-LCL was well tolerated in all patients (injection site reaction 5/7, fever 1/7). No clinical signs or laboratory parameters characteristic of an acute EBV infection or the onset of a lymphatic malignancy were observed. A limited number of adverse events was noted, but considered as treatment-unrelated, tumor-related symptoms (Table 4). The most common of these events were fatigue, nausea, and abdominal pain and did not exceed grade 3 of the common toxicity criteria (Trotti et al., 2003). Two grade 4 adverse events classified as tumor-related were observed: One patient suffered a pulmonary embolism and one patient committed suicide (Tables 3 and 4).
Adverse events according to common toxicity criteria.
Discussion
We report here the results of a phase I clinical study, in which seven patients were vaccinated with muRas-LCL. In six of seven patients (85%) we observed an increase in the number of muRas-specific T-lymphocytes in the peripheral blood by vaccination. Thus, it could be demonstrated that the muRas vaccine is immunogenic in vivo, confirming the observation that muRas-LCL elicit a muRas-specific T-cell response in vitro (Kubuschok et al., 2002). The strong DTH response in all of these six patients suggests that EBV viral components in the muRas vaccine effectively work as adjuvants (Schirrmacher, 2005). A clinical response was observed transiently in some of the patients (57%), especially in patients with earlier tumor stages. After 4 months follow-up, no patient had a persistent clinical response.
With respect to the interpretation of these results, the most important finding is that muRas-LCL can generate a strong immune response in cancer patients in vivo. Thus, tumor antigen-transfected LCL can serve as an efficient and easily available alternative to dendritic cells as antigen presenters in cancer vaccine trials (Palucka et al., 2011). We further demonstrated that in nearly all patients, the T-cell response after vaccination was dominated by interferon-gamma–secreting CD8+ and CD4+ T lymphocytes. In contrast, we found very few muRas-specific CD4+ T cells and no CD8+ T cells, which produced IL-4 or IL-10, confirming a predominantly Th1 T cell response to the vaccine. Moreover, we tested the cytotoxic activity of peripheral blood CD8+ T cells induced by the muRas vaccine and demonstrated that they are able to kill HLA-matched pancreatic cancer cells in vitro. However, since tumor progression was halted transiently in the majority of patients, we also assume a cytotoxic effect of the generated T-cell response against pancreatic cancer cells in vivo.
On the other hand, we eventually observed tumor progression despite the presence of muRas-specific T cells in the peripheral blood in all patients, after longer periods of vaccination. Possible reasons could be mechanisms of immune evasion adopted by the tumor cells as well as the local tumor micromilieu, with released cytokines (von Bernstorff et al., 2001; Tassi et al., 2008). For example, we could demonstrate that in case of progressing cancer under vaccination, the T-cell immune response became weaker and revealed a higher amount of IL-4–producing CD4+ T cells in comparison to the first weeks of the T-cell response to the vaccine. This may have indicated a beginning shift toward a Th2-type response, for example, mediated by cancer-released immunosuppressive cytokines like TGF-beta (von Bernstorff et al., 2001). Another reason for the progression despite presence of muRas-specific T cells may be that tumor antigen-specific T cells generated by vaccination can be characterized by low avidity and/or impaired function (Stuge et al., 2004). Moreover, the number of tumor antigen-specific T cells may also play a role in the efficacy of the tumor response. Some studies show that a frequency of one tumor antigen-specific T cell per 200 peripheral blood monocytes is not enough for an efficient anti-tumor response (Rosenberg et al., 1998; Dudley et al., 2002). In our study, the maximal frequency of tumor antigen-specific T cells measured during vaccination was 1 per 2500 peripheral blood monocytes.
Remarkably, despite the fact that no muRas-specific T-cell responses could be measured in two cancer patients before vaccination with muRas-LCL, a potent T-cell response was seen in these two patients after vaccination. This seems to be contradictory to the results of our in vitro vaccine experiments in which we could not demonstrate tumor antigen-specific T-cell responses, if the peripheral blood T cells were not primed by prior stimulation with tumor antigen peptide-pulsed dendritic cells (Kubuschok et al., 2002). Therefore, we had not expected the generation of a muRas-specific T-cell response in these two patients. Nonetheless, there are two explanations for this observation. It is possible that the T-cell response before vaccination against muRas was present but below the detection limit of our ELISPOT assay. Alternatively, one could imagine that the application of muRas-LCL in vivo can lead to the activation of dendritic cells, which themselves prime muRas-specific T cells. We hypothesize that subcutaneously administered muRas-LCL or antigenic material derived thereof arrive at the draining lymph nodes by migration (Walter et al., 1992) or transport, and can stimulate dendritic cells there upon phagocytosis and crosspresentation of muRas-LCL (Bickham et al., 2003; Kurts et al., 2010). The activation of dendritic cells is supported by EBV viral structures in the vaccine (Zinkernagel, 2003; Fontana et al., 2009), which may function as danger signals. In addition to the activation of dendritic cells, the subcutaneous way of administration assures an efficient stimulation of the memory T-cell response (Mullins et al., 2003).
The side effects of the muRas-LCL vaccination can be judged as low from our preliminary data. Besides the known side effects of vaccinations (local reaction and flu-like symptoms), there were no signs of toxicity or severe complications. It is important to emphasize that, to the best of our knowledge, we have applied lymphoblastoid cell lines for the first time in human subjects (Moore et al., 1957), and the effects of such an application are unknown. At least in animal experiments, SCID mice subcutaneously inoculated with lymphoblastoid cell lines developed human EBV-positive B-cell lymphoproliferative disorders (EBV-LPD) similar to those arising in immunodeficient humans. These proliferations could be eradicated by transfusion of in vitro–generated, autologous EBV-specific cytotoxic T lymphocytes (Lacerda et al., 1996). Therefore, we paid particular attention to the development of EBV-lymphoproliferative disease and observed neither local or systemic lymphomas nor clinical signs of lymphoproliferative disease in our LCL-treated patients. However, conclusions from these observations should be drawn with caution because of the limited number of patients and the limited follow-up time, which may exclude late manifestations of lymphomas. With regard to the T-cell response against EBV after vaccination with LCL, we noticed a clear increase of CD8+ T cells specific for LMP-2, which represents a latent protein relevant as target antigen for the immune control of EBV-positive malignancies (Murray et al., 1992). This indicates that the T-cell response against at least some EBV antigens is intact in pancreatic cancer patients and is able to control the proliferation of lymphomas in these patients.
In conclusion, genetically antigen-modified LCL represents a valuable and easily available tool for in vivo autologous tumor vaccination. LCL can be transfected with any known tumor antigen and therefore should be clinically investigated further for the treatment of cancers expressing molecularly defined tumor antigens (Disis et al., 2009). The most encouraging results concerning the cellular immune response and the progression-free survival were observed in patients in an early tumor stage (Kantoff et al., 2010; Schuster et al., 2011). Therefore, if this vaccine is applied in an advanced tumor stage it is reasonable to consider combination with blockade of inhibitory signals, for example, anti-CTLA-4 antibodies (Hodi et al., 2010), or depletion of down-regulatory T cells (Buckwalter and Srivastava, 2008; Schuster et al., 2011).
Footnotes
Acknowledgments
The authors gratefully acknowledge the excellent technical assistance of Ms. C. Schormann. We thank Prof. G. Bornkamm (Gesellschaft für Strahlen- und Umweltforschung, Munich, Germany), Prof. S. Stevanovic, Dr. C. Gouttefangeas (Institute of Cell Biology, University of Tuebingen), Dr. M. Ried (Gene Center, University of Munich), Prof. B. Gärtner, Prof. F. Grässer (Institute of Virology, University of Saarland) for helpful discussions, and Dr. D. Stevens (Department of Physiology, University of Saarland) for editing the manuscript. Supported by a grant of Saarland University (HOMFOR), Freunde der Universitätskliniken des Saarlandes e.V. and Alois-Lauer-Stiftung to B.K. and M.P.
Author Disclosure Statement
The authors declare no competing financial interests.