
Abstract
For genetic diseases that manifest at a young age with irreversible consequences, early treatment is critical and essential. Neonatal gene therapy has the advantages of achieving therapeutic effects before disease manifestation, a low vector requirement and high vector-to-cell ratio, and a relatively immature immune system. Therapeutic effects or long-term rescue of neonatal lethality have been demonstrated in several animal models. However, vigorous cell proliferation in the newborn stage is a significant challenge for nonintegrating vectors, such as adeno-associated viral (AAV) vector. Slightly delaying the injection age, and readministration at a later time, are two of the alternative strategies to solve this problem. In this study, we demonstrated robust and efficient hepatic gene transfer by self-complementary AAV8 vector in neonatal mice. However, transduction quickly decreased over a few weeks because of vector dilution caused by fast proliferation. Delaying the injection age improved sustained expression, although it also increased neutralizing antibody (NAb) responses to AAV capsid. This approach can be used to treat genetic diseases with slow progression. For genetic diseases with early onset and severe consequences, early treatment is essential. A second injection of vector of a different serotype at a later time may overcome preexisting NAb and achieve sustained therapeutic effects.
Introduction
Among the viral vectors tested for neonatal gene therapy, vectors based on lentivirus or retrovirus have demonstrated substantial long-term efficacy in animal models of such conditions as lysosomal storage disorders, adenosine deaminase deficiency, and hemophilia (Yoshimitsu et al., 2004; Zhang et al., 2004; Kobayashi et al., 2005; Carbonaro et al., 2006; Traas et al., 2007). However, insertional mutagenesis remains a concern for retroviral or lentiviral vector-mediated gene transfer (Hacein-Bey-Abina et al., 2008; Ginn et al., 2010). Nonintegrating vectors based on adenovirus or adeno-associated virus (AAV) have shown high transduction efficiency in neonatal gene transfer; however, both vector systems suffer significant loss of transgene expression because of dilution of vector genome during rapid cell proliferation, especially in liver (Wang et al., 2005; Cunningham et al., 2008; Hu et al., 2011). Despite the reduction of transgene expression levels and vector genome copies, therapeutic effects or long-term rescue of neonatal lethality have been demonstrated in several animal models (Daly et al., 2001; Carrillo-Carrasco et al., 2010; Chandler and Venditti, 2010; Yiu et al., 2010; Chandler et al., 2011; Cotugno et al., 2011; Hu et al., 2011). In these cases, expression of the normal gene in a small percentage of stably transduced cells accounts for the clinical effects. Interestingly, when vector is delivered at serially higher ages up to adulthood, transgene expression was shown to be progressively more durable, correlating with the decline in liver growth (Wang et al., 2005; Cunningham et al., 2008; Yiu et al., 2010; Cotugno et al., 2011). This strategy can be used in the treatment of diseases in which pathological changes happen later in infant life, such as some forms of lysosomal storage disorders. Alternatively, readministration at a later time after the initial vector administration could restore and sustain the therapeutic effects, although neutralizing antibodies resulting from the initial vector are a concern. In the canine model of glycogen storage disease type Ia (GSDIa), a single injection of AAV2/8 vector into 1-day-old dogs achieved transient correction. Redosing with an AAV2/1 vector 20 weeks after the initial AAV2/8 injection achieved sustained correction of the GSDIa phenotype (Weinstein et al., 2010). In the current study, we present the early kinetics of transgene expression in mouse liver after neonatal gene transfer of self-complementary (sc) AAV8 vector, the effects of delayed vector administration on transgene expression and neutralizing antibody responses, and redosing of AAVrh.10 to restore transgene expression.
Materials and Methods
AAV vector production
The conventional single-stranded (ss) AAV2/8.TBG.EGFP and AAV2/rh10.TBG.EGFP and self-complementary (sc) AAV2/8sc.TBG.EGFP and scAAV2/8.TBG.mOTC1.1 (Wang et al., 2011c) vectors were produced by Penn Vector Core at the University of Pennsylvania (Philadelphia, PA) as described previously (Gao et al., 2006).
Murine experiments
C57BL/6 male and female mice (6–8 weeks old) were purchased from Charles River Laboratories (Wilmington, MA) for the setting up of mating pairs. All animal procedures were performed in accordance with protocols approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Pennsylvania. Mating cages were monitored daily for birth. Neonatal gene transfer was performed on 1-day-old mice. Vector (5×1010 or 2×1011 genome copies [GC] per mouse) in a volume of 50 μl was delivered via the temporal vein as described (Daly, 2004). At various time points after injection, mice were killed. Body weight was measured, and liver was harvested and weighed. For delayed age at injection, newborn mice (1 day old) and 1-, 2-, 3-, 4-, 5-, and 6-week-old mice received an intraperitoneal injection of vector. Blood was collected by retro-orbital bleeding 6 weeks after vector injection. For the readministration experiment, 1-day-old mice were injected via the temporal vein with scAAV2/8.TBG.mOTC1.1 vector or not injected (control), and then 4 weeks later blood was collected by retro-orbital bleeding, and all mice received a tail vein injection of AAV2/rh10.TBG.EGFP vector.
Quantification of enhanced green fluorescent protein expression in liver
Enhanced green fluorescent protein (EGFP) transduction in mouse liver was quantified as the percentage of area expressing EGFP and the intensity of EGFP as described (Wang et al., 2010).
Vector genome copy analysis
Genomic DNAs were extracted from liver with a QIAamp DNA mini kit (Qiagen, Valencia, CA). Detection and quantification of vector genomes in extracted DNA were performed by real-time PCR as described previously, using primers and probe corresponding to the EGFP cDNA (Bell et al., 2006).
AAV neutralizing antibody assay
Serum samples were heat inactivated at 56°C for 30 min. Neutralizing antibody (NAb) assays were performed on Huh7 cells as previously described (Calcedo et al., 2009).
Statistical analysis
The significance of differences between the two dose groups was determined by Student t test. p<0.05 was considered statistically significant.
Results and Discussion
Robust but unstable transduction in neonatal mouse liver by self-complementary AAV8 vector
To assess liver transduction efficiency and kinetics in neonatal mice, an scAAV8 vector expressing EGFP under the control of a liver-specific thyroxine-binding globulin (TBG) promoter (AAV2/8sc.TBG.EGFP) was injected into 1-day-old mice via the superficial temporal vein, at a dose of 5×1010 or 2×1011 GC per pup. Animals were killed 1, 2, 3, 4, 5, 7, 9, 14, 21, and 35 days after vector administration and liver tissue was processed for fluorescence microscopy (Fig. 1) and morphometric quantification (Fig. 2A and B). EGFP expression in liver was detectable as early as 1 day after vector administration in about 10% of the liver areas inspected, and quickly increased and peaked at 7 days in almost 100% of the liver areas. The fast kinetics of transgene expression are likely due to the self-complementary AAV vector structure (Wang et al., 2003). EGFP expression gradually decreased through day 14, followed by a steep decline in expression by day 21. Low expression remained in 1–2% of liver areas and was maintained through day 35 (the end of the experiment). An increase in vector dose from 5×1010 to 2×1011 GC significantly improved EGFP expression during days 3–9 (Fig. 2A), but it did not change the expression kinetics or expression levels at the later time points (day 14 and after) (Fig. 2A and B). These expression patterns were in general consistent with what has been observed in previous AAV neonatal gene transfer studies (Cunningham et al., 2008; Hu et al., 2010). The detailed description of the early kinetics in the current study may help in the design of gene therapy studies of diseases with early onset or newborn lethal phenotypes such as ornithine transcarbamylase deficiency.

Robust but unstable expression of EGFP in liver after temporal vein injection of scAAV8 vector into neonatal mice. Newborn mice (1 day old) were injected via the temporal vein with 5×1010 GC of AAV2/8sc.TBG.EGFP. Liver was harvested 1, 2, 3, 4, 5, 7, 9, 14, 21, and 35 days postinjection for evaluation of EGFP expression. Representative images from each group are shown. Scale bar: 200 μm.

Correlation of gene expression, vector genomes in liver, and body and liver growth in neonatal mice after temporal vein injection of scAAV8 vector. Newborn mice (1 day old) were injected via the temporal vein with 5×1010 or 2×1011 GC of AAV2/8sc.TBG.EGFP. Livers were harvested 1, 2, 3, 4, 5, 7, 9, 14, 21, and 35 days postinjection. Transduction efficiency was evaluated by quantitative morphometric analysis of the intensity of the green fluorescence
Dilution of vector genomes and fast growth rate
Real-time quantitative PCR (Q-PCR) analysis of liver DNA showed a rapid decrease in vector genome copies. Vector GCs per diploid genome were reduced from 108 and 14 for the high- and low-dose groups, respectively, on day 7 to 1.2 and 0.5 GC, respectively, on day 21 after vector administration (Fig. 2C). The rate of decrease was equivalent to an approximately 10-fold reduction every 7 days. After day 21, the rate of decrease slowed and the vector GCs decreased only modestly from day 21 through day 35 (0.6 and 0.14 GC per diploid genome for the high- and low-dose groups, respectively). Growth rates of body and liver weight were monitored throughout the experiment. At the time of weaning, the average body weight increased from 1.5 g at 1 day after vector administration to 9.5 g on day 21, a 6.3-fold increase (Fig. 2D). Liver growth occurred at a similar rate. The fast proliferation of liver during this period is likely the main cause for the reduction of vector genomes in liver, because the majority of AAV DNA does not integrate. Although fast growth continued from day 21 to day 28, the reduction of vector genomes was modest (Fig. 2C). It is possible that some of the remaining vector DNA was integrated. EGFP-positive hepatocytes at the later time points appeared in clusters (Fig. 1). Further vector DNA structure analysis is required to confirm the integration.
Delaying the age at vector administration increases the stability of transduction
To investigate whether delaying the age at which vector was injected would improve gene transfer stability, AAV2/8.TBG.EGFP vector was injected intraperitoneally into newborn mice (1 day old) and 1-, 2-, 3-, 4-, 5-, and 6-week-old mice at a dose of 5×1010 GC per animal. Livers were evaluated for EGFP expression either at 6 weeks after vector administration or when mice were 12 weeks old (Fig. 3A and B). Transduction efficiency increased as the age at vector injection was increased (Fig. 3C), despite the fact that older mice received less vector on the basis of weight. Significant improvement of liver transduction stability (more than 30-fold) was observed when vector administration was delayed from newborn to 2 weeks of age. Further delaying the injection age to 4 weeks increased the stability only modestly. Vector genome copies in liver correlated well with transduction efficiency, with a significant increase from newborn to 2-week-old mice and a modest gain afterward (Fig. 3D). Consistent with previous findings (Cunningham et al., 2008; Cotugno et al., 2011), these data suggest that slightly delaying the age at which vector injection is done can significantly increase gene transfer stability. However, the length of the delay could vary between animal species. Cotugno and colleagues showed that delaying injection age from postnatal day 5 to day 50 in mucopolysaccharidosis VI cats greatly improved the long-term efficacy and vector dose required to achieve normal levels of serum levels of arylsulfatase B (Cotugno et al., 2011). Our gene transfer study in infant rhesus monkeys has shown that delaying the injection age from 1 week postnatal to 1 month was not sufficient to improve gene transfer stability (Wang et al., 2011a).
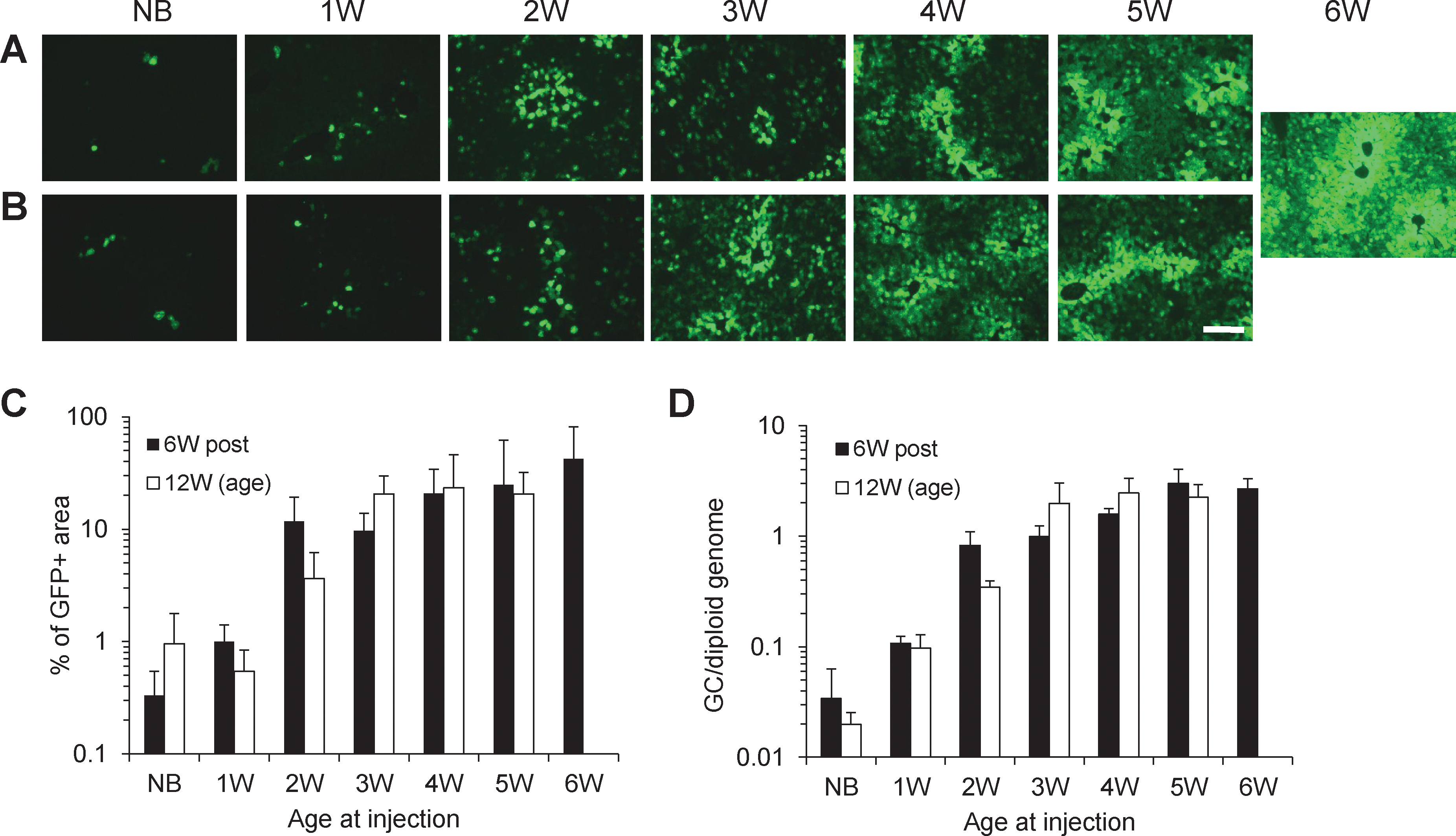
Delaying the age at which vector was injected increases the persistence of gene expression in liver. Mice at the age of 1 day (NB) or 1–6 weeks (1W–6W) were injected intraperitoneally with 5×1010 GC of AAV2/8.TBG.EGFP. EGFP expression was evaluated 6 weeks after vector administration
Delaying the age at vector administration increases the neutralizing antibody responses to AAV capsid
The influence of age of injection on NAb responses to AAV8 was assessed in animals 6 weeks after vector injection. As shown in Fig. 4, animals that received vector injection at incrementally older ages developed higher NAb titers to AAV8. Newborn mice had the lowest NAb titers to AAV8 at 6 weeks after vector injection, 1–2 logs lower than for older animals. On the basis of studies in nonhuman primates, NAb titers at 1:10 may not affect gene transfer efficiency (Wang et al., 2011b). Therefore, a second vector administration in these animals should be able to restore efficient gene transfer. However, the influence of age of injection on NAb response could also vary between animal species. Infant monkeys that received vector injection at postnatal age 1 month developed lower titers of AAV NAb than those injected at postnatal age 1 week, and the NAb titers quickly declined to 1:5 or 1:20 within 3 weeks of vector injection (Wang et al., 2011a).
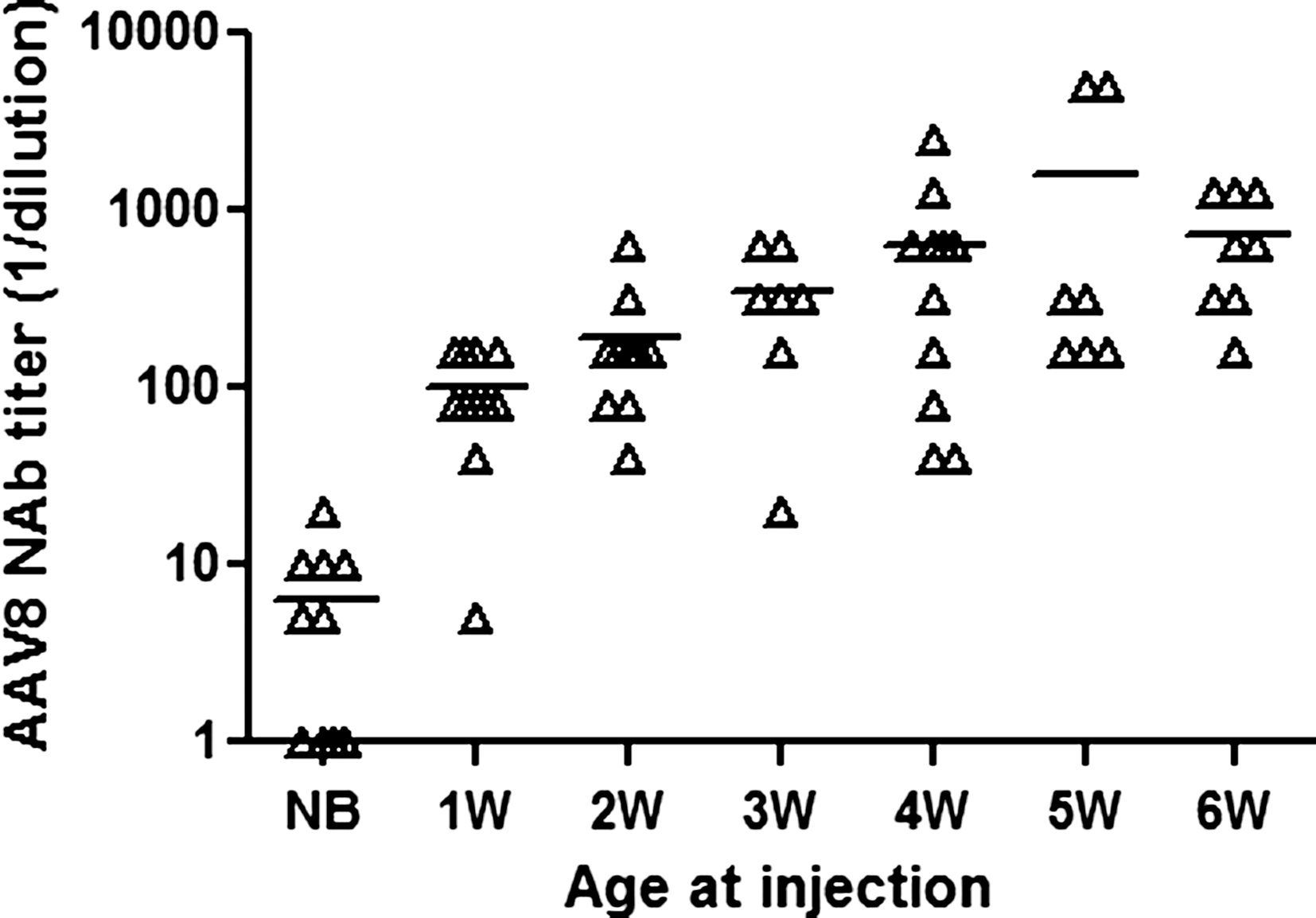
Lower AAV8 NAb titer in mice injected at a younger age. Mice at the age of 1 day (NB) or 1–6 weeks (1W–6W) were injected intraperitoneally with 5×1010 GC of AAV2/8.TBG.EGFP. Serum was obtained 6 weeks after vector injection for measurement of AAV8 NAb titer.
Successful readministration of AAV2/rh.10 vector
AAVrh.10, an AAV isolate from rhesus monkey (Gao et al., 2002, 2004), has been shown to be an efficient gene transfer vector in adult and neonatal mice (De et al., 2006; Nathwani et al., 2009; Hu et al., 2010). It also has been shown that despite more than 90% of amino acid sequence homology between the capsid proteins of AAV8 and AAVrh.10, there was little humoral immune cross-reactivity between these two AAV serotypes in mice (Nathwani et al., 2009). Therefore, we chose AAVrh.10 as a serotype for readministration in mice that received AAV8 vector at the neonatal stage. Newborn mice (1 day old) were first injected via the temporal vein with 5×1010 GC of scAAV2/8.TBG.mOTC1.1 vector or were not injected (control), and then, 4 weeks later, all mice received a tail vein injection of AAV2/rh10.TBG.EGFP at a dose of 1×1011 GC. EGFP expression in liver was evaluated 4 weeks after the AAVrh.10 injection. Equivalent and high levels of EGFP expression were observed in mice pretreated with AAV2/8 as newborns and in control mice injected only with AAVrh.10.TBG.EGFP (Fig. 5A–C). As expected, NAb titers to AAV8 and AAVrh.10 measured 4 weeks after the initial AAV2/8 vector treatment (before the injection of AAVrh.10) were low (Fig. 5D). Therefore, a readministration approach could be used to achieve sustained expression of therapeutic proteins in diseases with neonatal onset.

Efficient liver transduction by AAVrh.10 vector in mice preinjected with AAV8 vector at the newborn stage. Newborn mice (1 day old) were injected via the temporal vein with 5×1010 GC of AAV2/8sc.TBG.mOTC vector, and 4 weeks later received a second injection, via the tail vein, of 1×1011 GC of AAVrh.10sc.TBG.EGFP vector. Control animals received no vector injection as newborns, only AAVrh.10 at week 4. Livers were harvested 4 weeks after the second injection for evaluation of EGFP transduction.
In summary, we demonstrated robust and efficient hepatic gene transfer with an AAV8 vector in neonatal mice. However, transduction quickly decreased over a few weeks due to vector dilution caused by fast proliferation. Delaying the injection age improved sustained expression, although it also increased NAb responses to AAV capsid. This approach can be used to treat genetic diseases with slow progression or with effective methods to control disease progression. For genetic diseases with early onset and severe consequences, early treatment is essential. A second injection of vector of a different serotype at a later time may overcome preexisting NAb and achieve sustained therapeutic effects.
Footnotes
Acknowledgments
The authors thank Julie Johnston, Arbans Sandhu, Martin Lock, and Shu-Jen Chen (Penn Vector) for supplying vectors; Roberto Calcedo and the Immunology Core of the Gene Therapy Program for neutralizing antibody analysis; and Hongwei Yu and Theodore Drivas for technical assistance. This work was supported in part by the Kettering Family Foundation and the following grants to J.M.W.: P01-HD057247, P01-HL059407, and P30-DK047757. H.W. was supported by a scholarship from the China Scholarship Council.
Author Disclosure Statement
J.M.W. is a consultant to ReGenX Holdings, and is a founder of, holds equity in, and receives a grant from affiliates of ReGenX Holdings; in addition, he is an inventor on patents licensed to various biopharmaceutical companies, including affiliates of ReGenX Holdings.