
Abstract
Sarcopenia is the progressive age-related loss of skeletal muscle mass associated with functional impairments that reduce mobility and quality of life. Overt muscle wasting with sarcopenia is usually preceded by a slowing of the rate of relaxation and a reduction in maximum force production. Parvalbumin (PV) is a cytosolic Ca2+ buffer thought to facilitate relaxation in muscle. We tested the hypothesis that restoration of PV levels in muscles of old mice would increase the magnitude and hasten relaxation of submaximal and maximal force responses. The tibialis anterior (TA) muscles of young (6 month), adult (13 month), and old (26 month) C57BL/6 mice received electroporation-assisted gene transfer of plasmid encoding PV or empty plasmid (pcDNA3.1). Contractile properties of TA muscles were assessed in situ 14 days after transfer. In old mice, muscles with increased PV expression had a 40% slower rate of tetanic force development (p<0.01), and maximum twitch and tetanic force were 22% and 16% lower than control values, respectively (p<0.05). Muscles with increased PV expression from old mice had an 18% lower maximum specific (normalized) force than controls, and absolute force was ∼26% lower at higher stimulation frequencies (150–300 Hz, p<0.05). In contrast, there was no effect of increased PV expression on TA muscle contractile properties in young and adult mice. The impairments in skeletal muscle function in old mice argue against PV overexpression as a therapeutic strategy for ameliorating aspects of contractile dysfunction with sarcopenia and help clarify directions for therapeutic interventions for age-related changes in skeletal muscle structure and function.
Introduction
It is well accepted that impairments in Ca2+ regulation contribute to the muscle wasting and weakness that accompany aging. One of the contributing factors is that the loss of muscle mass in aging is preceded by a slowing in the maximal rate of relaxation of the twitch and tetanus (Ryall et al., 2004; Schertzer et al., 2005; Russ et al., 2011) and a reduction in both peak tetanic intracellular Ca2+ concentration ([Ca2+]i) and maximal tetanic force (Gonzalez et al., 2003; Ryall et al., 2004; Schertzer et al., 2005; Russ et al., 2011). The reduced peak tetanic [Ca2+]i is attributed to an elevated resting [Ca2+]i, which is 25% and 74% higher in soleus and extensor digitorum longus (EDL) muscles from old (27–30 months) compared with young rats (4–6 months), respectively (Fraysse et al., 2006). It may also be attributed to compromised stress-induced activation of Ca2+ sparks, which facilitate SR Ca2+ release under conditions of membrane deformation in muscles from young healthy mice, but which are absent in muscles from aged mice (Weisleder et al., 2006). The elevated resting [Ca2+]i is caused by effects on mitochondrial enzymes and increased Ca2+ extrusion (Gonzalez et al., 2003; Schertzer et al., 2005; Fraysse et al., 2006). These effects are consistent with age-associated reductions in SR Ca2+ uptake (Schertzer et al., 2005) and SR Ca2+ release (Wang et al., 2000). Unfortunately, very few studies have examined whether muscles in old humans also exhibit impaired muscle Ca2+ regulation, but experiments using single muscle fibers found reduced SR Ca2+ release due to uncoupling of the dihydropyridine receptor and SR ryanodine receptor in muscle fibers from older (65–75 years) compared with younger individuals (25–35 years) (Delbono et al., 1995). These results indicate that as for rodents, age-associated impairments in Ca2+ regulation may also contribute to the muscle wasting and weakness in humans. Thus, attenuating the rise in resting [Ca2+]i should ameliorate skeletal muscle weakness in old animals and humans (Fraysse et al., 2006).
Elevated resting [Ca2+]i is also a characteristic of Duchenne muscular dystrophy (DMD) and has been shown to be 275% higher in quadriceps muscle biopsies from DMD patients compared with healthy controls (Maunder-Sewry et al., 1980). The absence of parvalbumin (PV), an endogenous high-affinity Ca2+ buffer from EDL (fast-twitch) muscles of wild-type mice enhanced maximal force production, but in dystrophic mdx mice it reduced maximal force production (Raymackers et al., 2003). These findings suggest that PV is beneficial in conditions of pathologically elevated [Ca2+]i, but not in otherwise healthy controls. Based on the observation that aging is accompanied by a large (63%) decrease in PV protein abundance in rat EDL muscles (Cai et al., 2001), restoration of PV levels in skeletal muscles could potentially enhance force production in old mice. PV overexpression has therefore been proposed as a potential intervention for sarcopenia (Arif, 2009).
The aim of this study was to investigate whether increased PV expression enhanced peak tetanic force in muscles of old mice. We tested the hypothesis that restoration of PV levels via electroporation-assisted gene transfer would increase peak tetanic force and hasten tetanic relaxation in skeletal muscles of old mice.
Materials and Methods
Animals and experimental design
All experiments were approved by the Animal Ethics Committee of The University of Melbourne and conducted in accordance with the Australian code of practice for the care and use of animals for scientific purposes as stipulated by the National Health and Medical Research Council (Australia). Young (6 month, n=14), adult (13 month, n=14), and old (26 month, n=18) female C57BL/6 mice were obtained from The University of Queensland Biological Resources (Brisbane, Queensland) and housed in the Biological Research Facility at The University of Melbourne under a 12-hr light/dark cycle, with drinking water and standard chow provided ad libitum. Mice were allocated randomly into one of two groups: the first received an intramuscular injection of plasmid DNA encoding PV, followed by electroporation to facilitate plasmid DNA uptake into muscle fibers based on our published methods (Schertzer et al., 2006; Schertzer and Lynch, 2008); and the second group received an intramuscular injection of empty vector (pcDNA3.1) followed by electroporation. Two weeks after electroporation-assisted gene transfer, contractile properties of the right tibialis anterior (TA, predominantly fast-twitch) muscle were assessed in situ according to our well-established protocols (Gehrig et al., 2010; Murphy et al., 2010). Two weeks was selected for evaluation because we have previously shown maximum transfection efficiency and minimal muscle damage at this time following electroporation (Schertzer et al., 2006). At the conclusion of the functional measurements, the right and left TA muscles were carefully dissected, blotted on filter paper, and weighed on an analytical balance. Muscles were mounted in embedding medium, frozen in thawing isopentane, and stored at −80°C for subsequent analyses. Mice were killed by rapid cardiac excision while anesthetized deeply. The excised heart was trimmed of connective tissue, blotted on filter paper, and weighed.
Plasmid DNA preparation and electroporation
A mammalian expression vector encoding mouse PV (MMM1013-7513897, Open Biosystems, Huntsville, AL) was used in these experiments. Plasmid DNA was isolated by anion-exchange using Endotoxin free Giga kits from Qiagen (Doncaster, VIC, Australia), according to the manufacturer's instructions.
Mice were anesthetized intraperitoneally with a mixture of ketamine (100 mg/kg) and xylazine (10 mg/kg) such that they were unresponsive to tactile stimuli. We have previously optimized the method of electroporation-assisted plasmid DNA transfer in mouse skeletal muscle to maximize transfer and to minimize muscle damage (Schertzer et al., 2006; Schertzer and Lynch, 2008). Briefly, the TA muscle of the right hindlimb was surgically exposed and injected with 30 μl of 0.5 U/μl hyaluronidase (Sigma-Aldrich, Castle Hill, NSW, Australia) and 2 hr later, after re-anesthetizing the mice with ketamine and xylazine, and injecting with (40 μl, 2 μg/μl) plasmid encoding PV or vector alone (pcDNA3.1) using a 29-gauge needle. Approximately 1 min after the plasmid DNA injection, three 20-msec square wave pulses (150 V/cm) of 1-Hz frequency generated by a Grass stimulator (Grass S88, Quincy, MA) were delivered transcutaneously across the TA muscle using custom-built platinum plate electrodes attached to digital calipers (Schertzer et al., 2006; Schertzer and Lynch, 2008). The polarity was then reversed and a further three pulses were delivered to the muscle (Schertzer et al., 2006; Schertzer and Lynch, 2008).
In situ analysis of TA contractile properties
Mice were anesthetized with sodium pentobarbitone (Nembutal, 60 mg/kg, Sigma-Aldrich) via intraperitoneal injection. The methods for assessment of the contractile properties of mouse TA muscles in situ have been described in detail by us previously (Gehrig et al., 2010; Murphy et al., 2010). Briefly, optimal muscle length (Lo) was determined from micromanipulation of muscle length to produce maximum isometric twitch force (Pt). Maximum isometric tetanic force (Po) was recorded from the plateau of the frequency-force relationship (10 Hz to 300 Hz for 350 msec with a 2-min rest between stimuli). Optimum fiber length (Lf) was determined by multiplying Lo by the TA Lf/Lo ratio of 0.6 (Burkholder et al., 1994). Muscle mass, Lf, and Po were used to calculate specific force (sPo), or the force normalized per total muscle fiber CSA (kN/m2) taking into account muscle density (1.06 mg/mm3).
Real-time RT-PCR analyses
Total RNA was extracted from 10–20 mg of TA muscle using a commercially available kit, according to the manufacturer's instructions (PureLink RNA Mini Kit, Invitrogen, Carlsbad, CA). RNA concentration was determined spectrophotometrically at 260 nm, and the samples were stored at −80°C. RNA was transcribed into cDNA using the Invitrogen SuperScript™ VILO cDNA Synthesis Kit, and the resulting cDNA stored at −20°C for subsequent analysis. Real-Time RT-PCR was performed as described previously (Murphy et al., 2010). The forward and reverse primer sequences used were: PV, 5′-TCTCCTGCCTGCTTGGTACT-3′ and 5′-CTTCCCTTCCCTTCTGCTCT-3′; troponin C fast, 5′-GGAGATCGAATCCCTGATGA-3′ and 5′-TGGATGGACACGAACAAAGA-3′; and troponin C slow, 5′-CTGCAGGAGATGATTGACGA-3′ and 5′-CCTCAGACTTCCCTTTGCTG-3′, respectively. Primer sequences for the myosin heavy chains (MHCs) MHCI, MHCIIa, MHCIIx, and MHCIIb were as detailed previously (Murphy et al., 2010). The content of single-stranded DNA (ssDNA) in each sample was determined using the Quanti-iT™ OliGreen® ssDNA Assay Kit (Molecular Probes, Eugene, OR), as described previously (Lundby et al., 2005; Murphy et al., 2010). Gene expression was quantified by normalizing the logarithmic cycle threshold (CT) value (2−CT) to the cDNA content of each sample to obtain the expression 2−CT/cDNA content (ng/ml).
Western blotting analyses
Muscle samples (20–30 mg) were homogenized (Polytron 2100, Kinematica, Lucerne, Switzerland) three times for 15 sec on ice in a 1:10 dilution with homogenizing buffer: 10 mM Tris-HCl, pH 7.5, 100 mM sodium chloride, 1 mM ethylenediamine-tetraacetic acid (EDTA), 1 mM ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA), 10% glycerol, 1% Triton X-100, 0.1% sodium dodecyl sulfate (SDS), 1 mM sodium fluoride, 20 mM sodium pyrophosphate, 2 mM sodium orthovanadate, 0.5% sodium deoxycholate, 1 mM phenylmethanesulphonylfluoride (PMSF), 0.1% protease inhibitor cocktail (P8340, Sigma-Aldrich), and 0.1% phosphatase inhibitor cocktail (P2850, P5726, Sigma-Aldrich). Samples were centrifuged at 10,000×g for 10 min at 4°C and the resulting supernatant analyzed for total protein content (Bio-Rad DC Protein Assay, Bio-Rad, Gladesville, NSW, Australia), with bovine serum albumin (BSA) as the standard. Protein loading buffer (40% glycerol, 6% SDS, 250 mM Tris-HCl, pH 6.8, 0.04% bromophenol blue, and 415 nM dithiothreitol) was added to 50 μg of sample in a 1:4 dilution and samples were heated at 95°C for 5 min before being stored at −20°C for immunoblotting.
SDS-PAGE (4%–12% NuPAGE® Novex® Bis-Tris Gel, Invitrogen) was performed and loaded with 15 μg of protein. After electrophoresis (30 min at 100 V, 90 min at 150 V), the protein was wet transferred overnight (100 mA) at 4°C (Trans-Blot Cell, Bio-Rad) to polyvinylidene difluoride membrane (Immobilon-P; Millipore, North Ryde, NSW, Australia). The following day the membrane was incubated in Ponceau stain for 5 min to visually verify equal protein transfer. The membrane was destained with distilled water and washed with Tris-buffered saline-Tween 20 (TBST) before being blocked for 2 hr at room temperature in TBST containing 5% skim milk powder. Membranes were incubated overnight at 4°C in anti-PV antibody (1:5,000, 12 kDa, PA1-933, Thermo Fisher Scientific, Rockford, IL). Membranes underwent three 10-min washes in TBST and were incubated for 1 hr at room temperature in a donkey anti-rabbit IgG horseradish peroxidase (HRP)-conjugated secondary antibody (GE Healthcare, Little Chalfont, Buckinghamshire, UK) diluted in TBST containing 5% skim milk powder. After three 10-min washes in TBST, membranes were treated with enhanced chemiluminescence (SuperSignal West Pico Chemiluminescent Substrate, Thermo Fisher Scientific). The signal was imaged using ChemiDoc XRS machine (Bio-Rad) and blots were quantified using Quantity One® software (Bio-Rad). Membranes were then stripped and blocked for 1 hr at room temperature in TBST containing 5% skim milk powder before being incubated for 2 hr at room temperature in anti-GAPDH (1:10,000, 37 kDa, G8795, Sigma-Aldrich). Membranes were then washed three times for 10 min in TBST and incubated for 1 hr at room temperature in a sheep anti-mouse IgG HRP-conjugated secondary antibody (GE Healthcare) diluted in TBST containing 5% skim milk powder. After three 10-min washes in TBST, membranes were treated with chemiluminescence and the signal was imaged and quantified. PV protein abundance was normalized against GAPDH protein abundance.
Cell culture, transfer, and measurement of [Ca2+]i
We invested considerable time and effort attempting to acquire accurate measurements of contraction-induced changes in [Ca2+]i in TA muscles transfected with plasmid encoding PV or an empty plasmid, but this was not possible because of movement of muscle fibers even during isometric contractions and the inaccuracies associated with tracking individual fibers during contraction and relaxation. The large size of the TA muscle also precluded reproducible delivery of the calcium ionophore, A23187, preventing assessments of PV overexpression attenuating the A23187-induced rise in [Ca2+]i in vivo. We used instead a cell culture preparation to assess the efficacy of PV gene transfer on [Ca2+]i because of its reproducibility and reliability. C2C12 cells were chosen because they are a mouse myoblast cell line. C2C12 cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% (v/v) fetal bovine serum and 2 mM L-glutamine (Invitrogen). Cells were plated into 12-well plates on glass coverslips coated with extracellular matrix proteins (ECM, Sigma-Aldrich), and upon reaching 80% confluence were transfected with 5 μg/well of plasmid using Lipofectamine™ 2000 (Invitrogen) according to the manufacturer's instructions. After 48 hr, cells were incubated with a loading solution consisting of HEPES-buffered physiological saline (120 mM NaCl, 4.75 mM KCl, 1.44 mM MgSO4, 11 mM glucose, 25 mM HEPES, 1.5 mM Ca2+, pH 7.3) supplemented with 2 μM Fluo-4/AM for 20 min, then incubated in the loading solution without Fluo-4 for a further 20 min to complete de-esterification of the probe.
Loading solution was then replaced with standard physiological saline and cells were imaged using an LSM 510 META multiphoton/confocal microscope equipped with 488 nm argon laser and a 40×IR-Achromat (NA 0.80) water-immersion objective (Carl Zeiss, Wrek Göttingen, Germany).
Skeletal muscle histology
Serial sections were cut transversely through the TA muscle using a refrigerated cryostat (−20°C, Microm cryostat, Microm International GmbH, Walldorf, Germany). To assess PV protein expression, sections were stained with an anti-PV antibody (PA1-933, Affinity Bioreagents, Rockford, IL), using techniques described previously (Murphy et al., 2010). Briefly, sections were fixed for 5 min in methanol, followed by 1 min in acetone. Slides were then incubated overnight at 4°C with an anti-PV antibody (1:200 in PBS). Slides were rinsed three times for 5 min with PBS and then incubated for 45 min with anti-rabbit secondary antibody in PBS. After several washes with PBS, reacted sections were embedded in Mowiol and covered with a coverslip. All muscle cross-sections were reacted with the same antibody preparation to minimize variability. Sections were also stained with hematoxylin and eosin (H&E) to determine general muscle fiber architecture. Digital images of stained sections were obtained using an upright microscope with camera (Axio Imager D1, Carl Zeiss, Wrek Göttingen, Germany), controlled by AxioVision AC software (AxioVision AC Rel. 4.7.1, Carl Zeiss Imaging Solutions, Wrek, Wrek Göttingen, Germany).
Statistical analyses
All values are expressed as mean±SE, unless otherwise stated. PV and pcDNA3.1 groups were compared using an unpaired t-test or a two-way ANOVA, where appropriate. Bonferroni's post hoc test was used to determine significant differences between individual groups. The level of significance was set at p<0.05 for all comparisons.
Results
Model of sarcopenia
We first established the suitability of the mice for studying age-related changes in skeletal muscle. We examined the mass of several hindlimb muscles including the TA, EDL, soleus, plantaris, gastrocnemius, and quadriceps, and all muscles exhibited an age-related reduction in mass (Fig. 1). Consistent with other animal models of sarcopenia, old mice had an increased heart mass in both absolute and relative terms (Table 1). Furthermore, in situ assessment of TA muscle function revealed an age-related deficit in maximum force-producing capacity (Fig. 2). The significant reduction in muscle mass and function in the old compared with adult and young mice established that this model was suitable for studying whether muscle-specific overexpression of PV ameliorated aspects of sarcopenia.
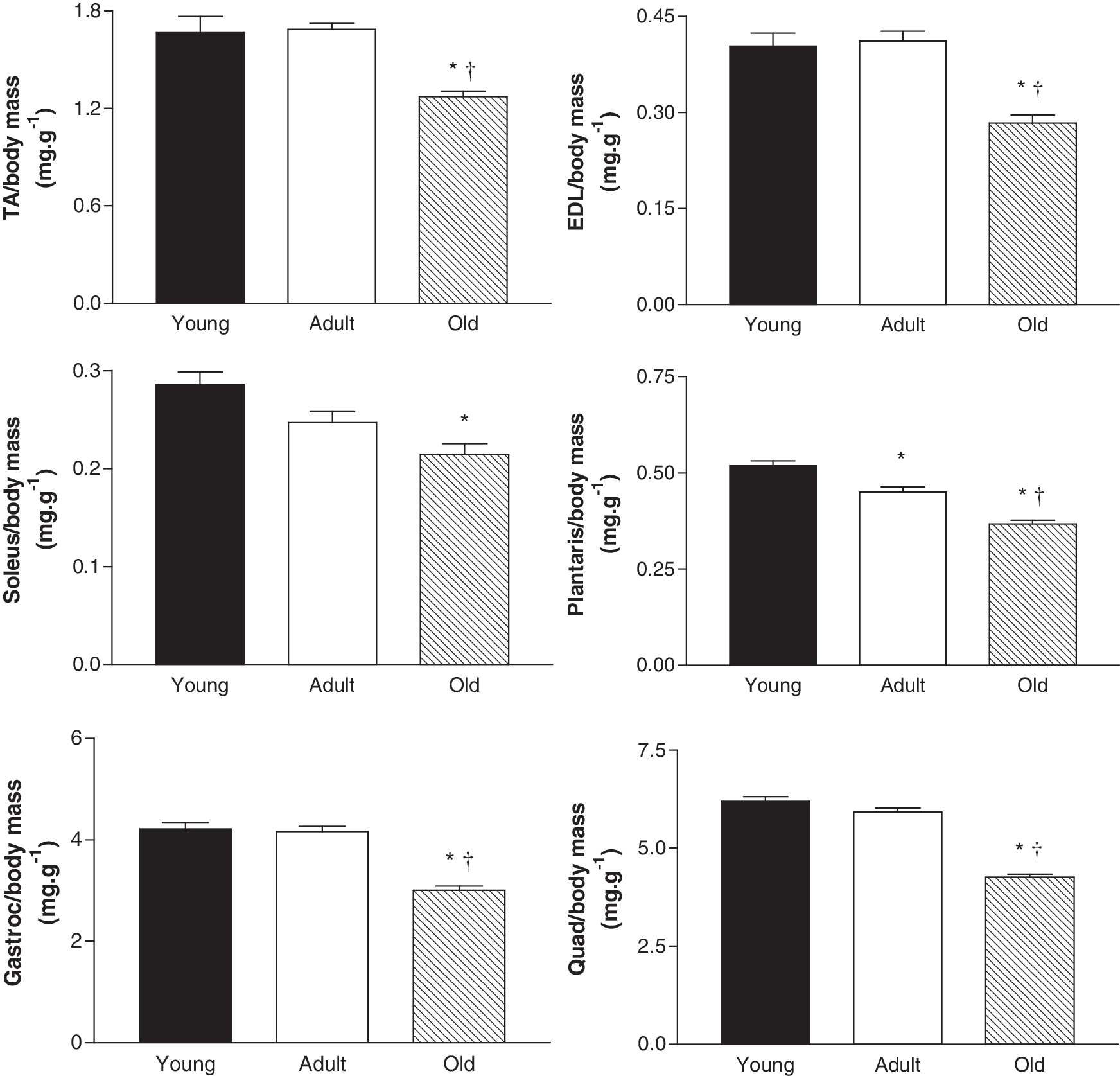
Relative mass of selected muscles from young (6 months), adult (13 months), and old (26 months) female C57BL/6 mice normalized to body mass. TA, tibialis anterior; EDL, extensor digitorum longus; Gastroc, gastrocnemius; Quad, quadriceps. Data are means±SE; n=14. *p<0.05 vs. young; †p<0.05 vs. adult.
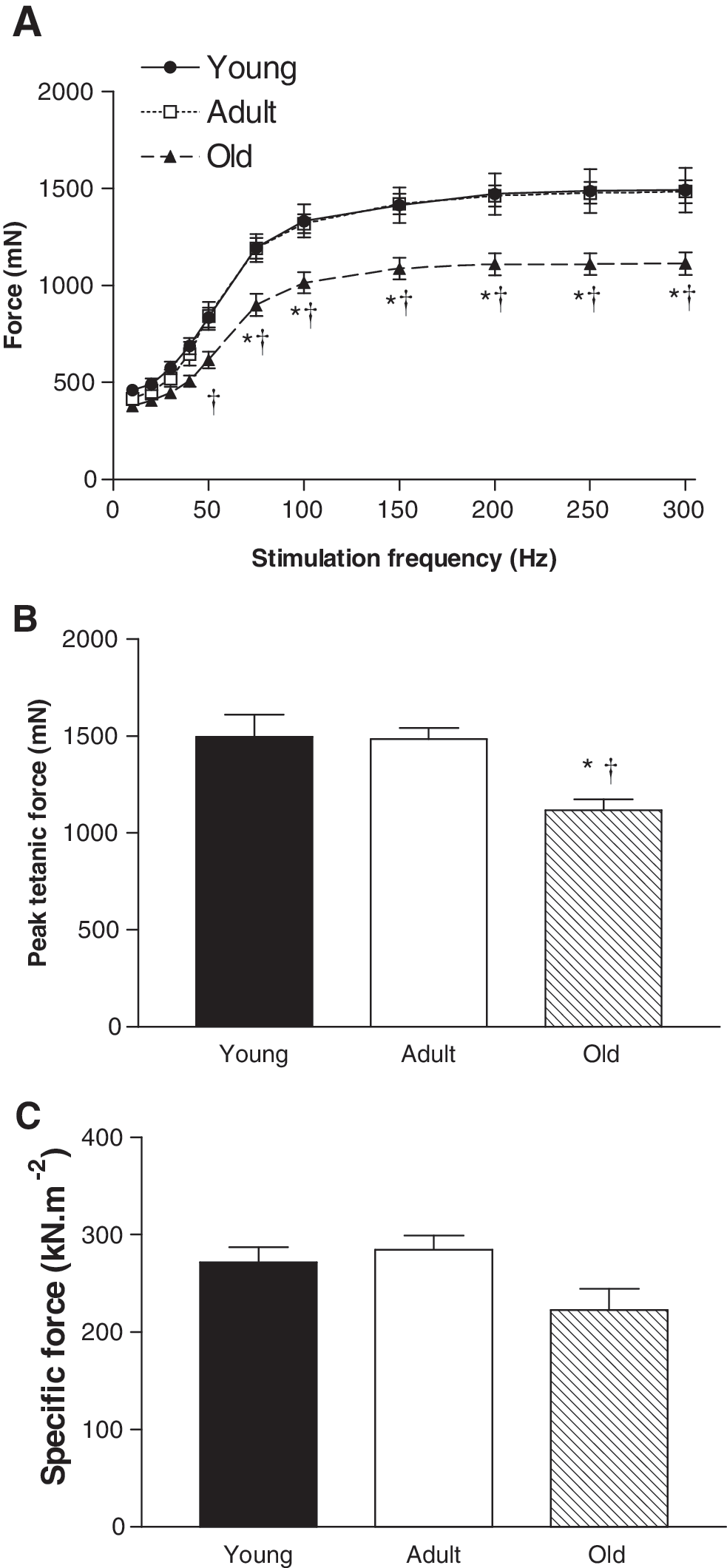
Functional properties of TA muscles from young (6 months), adult (13 months), and old (26 months) female C57BL/6 mice assessed in situ.
TA, tibialis anterior. Data are means±SE; n=13–16. No significant differences were found between treatment groups (pcDNA3.1 vs. PV) for any age.
P<0.05 vs. young pcDNA3.1.
P<0.05 vs. adult pcDNA3.1.
TA muscle PV mRNA expression was similar between young and adult mice, but was ∼90% lower in old compared with young and adult mice (p<0.05, Fig. 3A). Representative Western blots showing PV and GAPDH protein abundance in young, adult, and old mice are presented in Fig. 3B and quantification revealed that PV protein abundance was 35% lower in old compared with young mice (p<0.05, Fig. 3C).
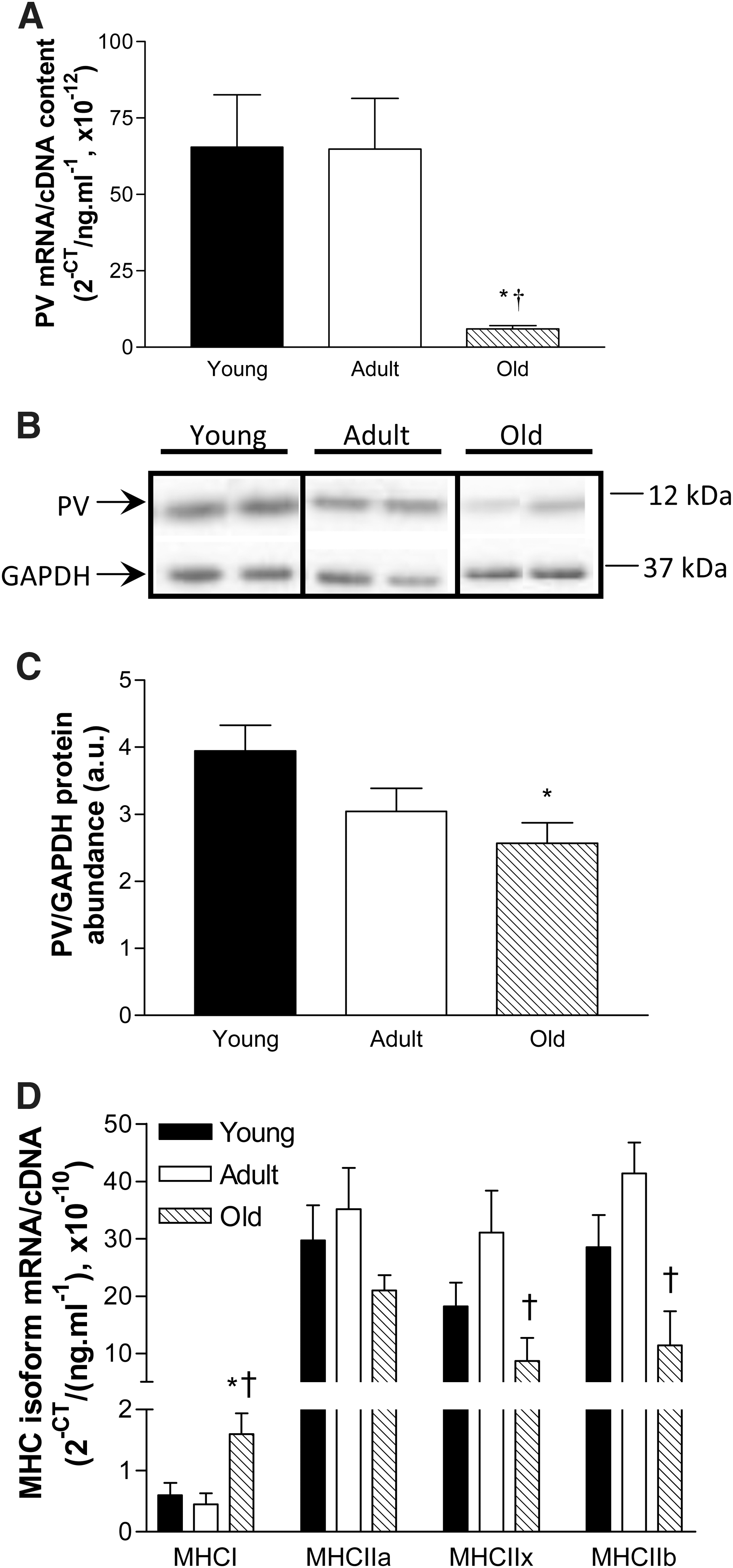
Parvalbumin (PV) mRNA and protein expression and myosin heavy chain (MHC) isoform mRNA expression in TA muscles from young (6 months), adult (13 months), and old (26 months) female C57BL/6 mice.
Gene expression of the MHC isoforms was investigated to determine whether there was an age-related shift in fiber type expression. MHCI mRNA expression was 159% and 254% higher in TA muscles from old mice compared with young and adult mice, respectively (P<0.05, Fig. 3D). MHCIIa mRNA expression was not significantly different between groups (p<0.31), but old mice had 72% and 74% lower expression of MHCIIx and MHCIIb mRNA than adult mice, respectively (p<0.05, Fig. 3D).
Efficacy of PV transfer in vitro and in vivo in old mice
When transfected C2C12 cells loaded with Fluo-4 were stimulated with the ionophore A23187 (1 μM), the peak increase in fluorescence in PV-transfected cells was 59% lower than in cells transfected with empty vector (pcDNA3.1, p<0.001, Fig. 4A).
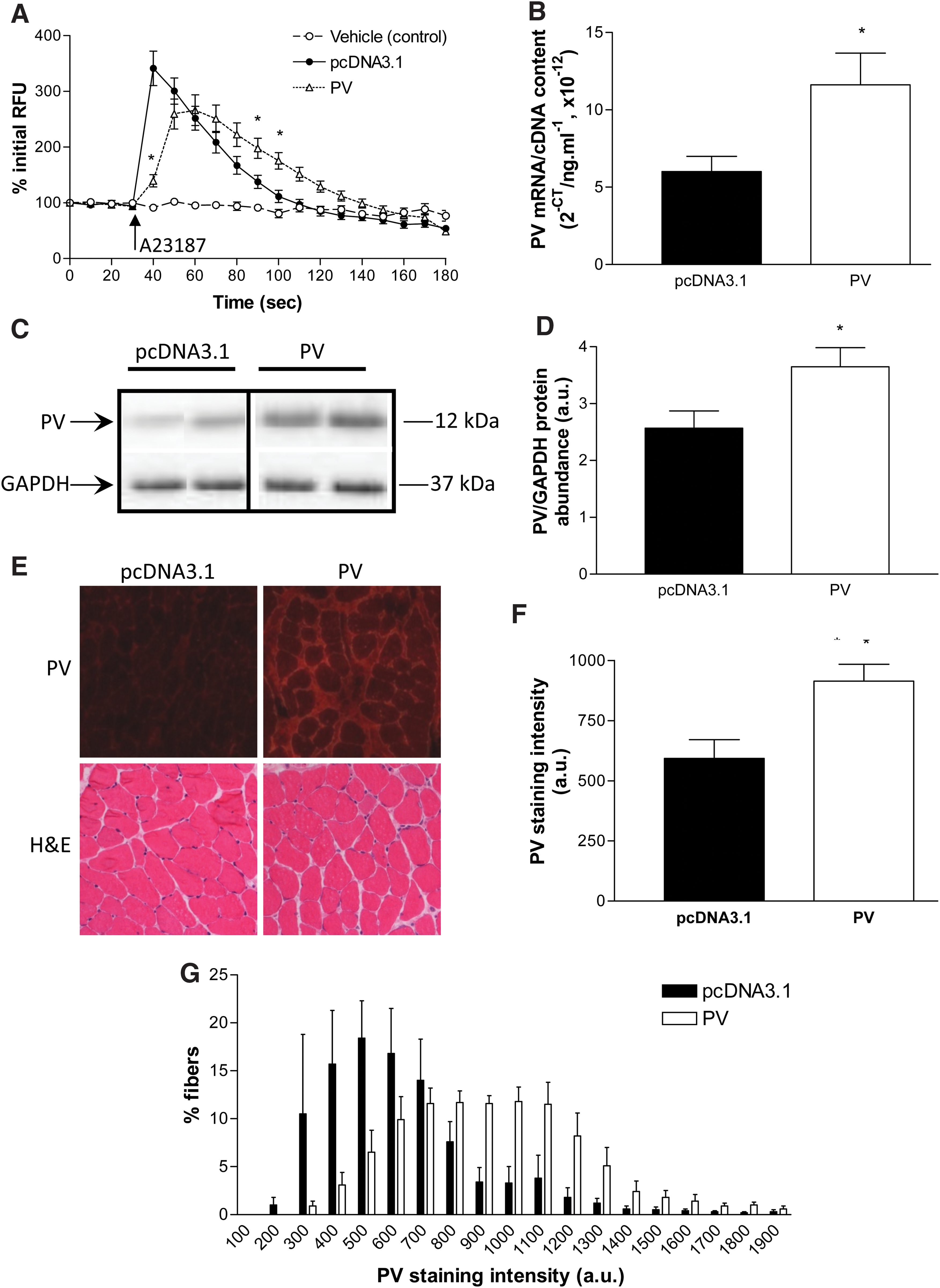
Efficacy of plasmid encoding PV for attenuating [Ca2+]i in vitro and quantification of PV overexpression induced via electroporation-assisted transfer in vivo in old (26 months) mice. C2C12 cells transfected with plasmid encoding PV (n=16) showed a smaller increase in fluorescence in response to stimulation with 1 μM A23187 compared with cells transfected with empty vector (pcDNA3.1, n=12)
TA muscles from old mice were examined at 2 weeks after in vivo electroporation-assisted transfer of plasmid encoding PV or empty vector and analyzed for PV mRNA (Fig. 4B) and protein expression (Fig. 4C, D). PV mRNA and protein expression were 95% and 42% higher, respectively, in old muscles transfected with plasmid encoding PV than with those transfected with empty vector (p<0.05, Fig. 4). TA muscle cross-sections were reacted for PV (Fig. 4E, top panel) and quantification revealed a 54% higher average PV staining intensity in old muscles transfected with plasmid encoding PV than those transfected with empty vector (Fig. 4F). The frequency histogram revealed a shift to the right for muscles transfected with plasmid encoding PV such that these muscles had a greater proportion of fibers with high PV staining (reaction) intensity and a smaller proportion of fibers with low PV staining (reaction) intensity (Fig. 4G). TA muscle cross-sections stained for H&E revealed similar muscle fiber architecture, including homogeneity of fiber size, peripherally located nuclei, and the amount of nonmuscle tissue, between groups (Fig. 4E, bottom panel).
PV gene transfer does not alter muscle mass in old mice
As expected, there was no difference between treatment groups in the percentage change in body mass over the 2-week period after gene transfer in old mice (Table 1). There was also no difference between treatment groups in the mass of the right TA (transfected) or left TA muscle (untreated), when muscle mass was normalized for body mass or when mass of the right TA was normalized to that of the left TA muscle (Table 1). Absolute and normalized heart mass were not different between treatment groups (Table 1).
PV gene transfer slows relaxation and impairs tetanic force production in situ in old mice
There was no difference in the time-to-peak twitch tension in muscles from old mice injected with empty vector or plasmid encoding PV, but the one-half relaxation rate was 23% slower after PV gene transfer (p<0.04, Fig. 5A). During maximal tetanic contractions, muscles from old mice with PV gene transfer exhibited a 40% slower rate of force development (+dPt/dt, p<0.01) compared with controls, but there was no difference in the rate of tetanic relaxation (−dPt/dt, p<0.44) between groups (Fig. 5B).

Twitch and tetanic properties and force production in situ in TA muscles of old (26 months) mice 2 weeks after electroporation-assisted transfer of plasmid encoding parvalbumin (PV) or empty vector (pcDNA3.1).
In old mice, TA muscles with PV gene transfer produced 22% (p<0.01) and 16% (p<0.02) lower peak forces during a twitch and tetanic contraction, respectively (Fig. 5C). When peak tetanic force was normalized for the muscle cross-sectional area, specific force was 18% lower in muscles with PV gene transfer (p<0.05, Fig. 5D). Examination of the frequency–force relationship revealed that muscles with PV gene transfer produced 25%–27% lower forces at higher stimulation frequencies (150–300 Hz, p<0.05, Fig. 6A). When force was normalized to that produced at 300 Hz, there was a left shift of the frequency–force relationship at the lower stimulation frequencies (30–75 Hz) in muscles with PV gene transfer, with normalized force being 10%–18% higher than in control muscles (p<0.01, Fig. 6B). Muscles with PV gene transfer first achieved tetanus at a lower (19%) stimulation frequency than controls (p<0.01, Fig. 6C), and also achieved peak tetanic force at a lower (27%) stimulation frequency than controls (p<0.01, Fig. 6D).
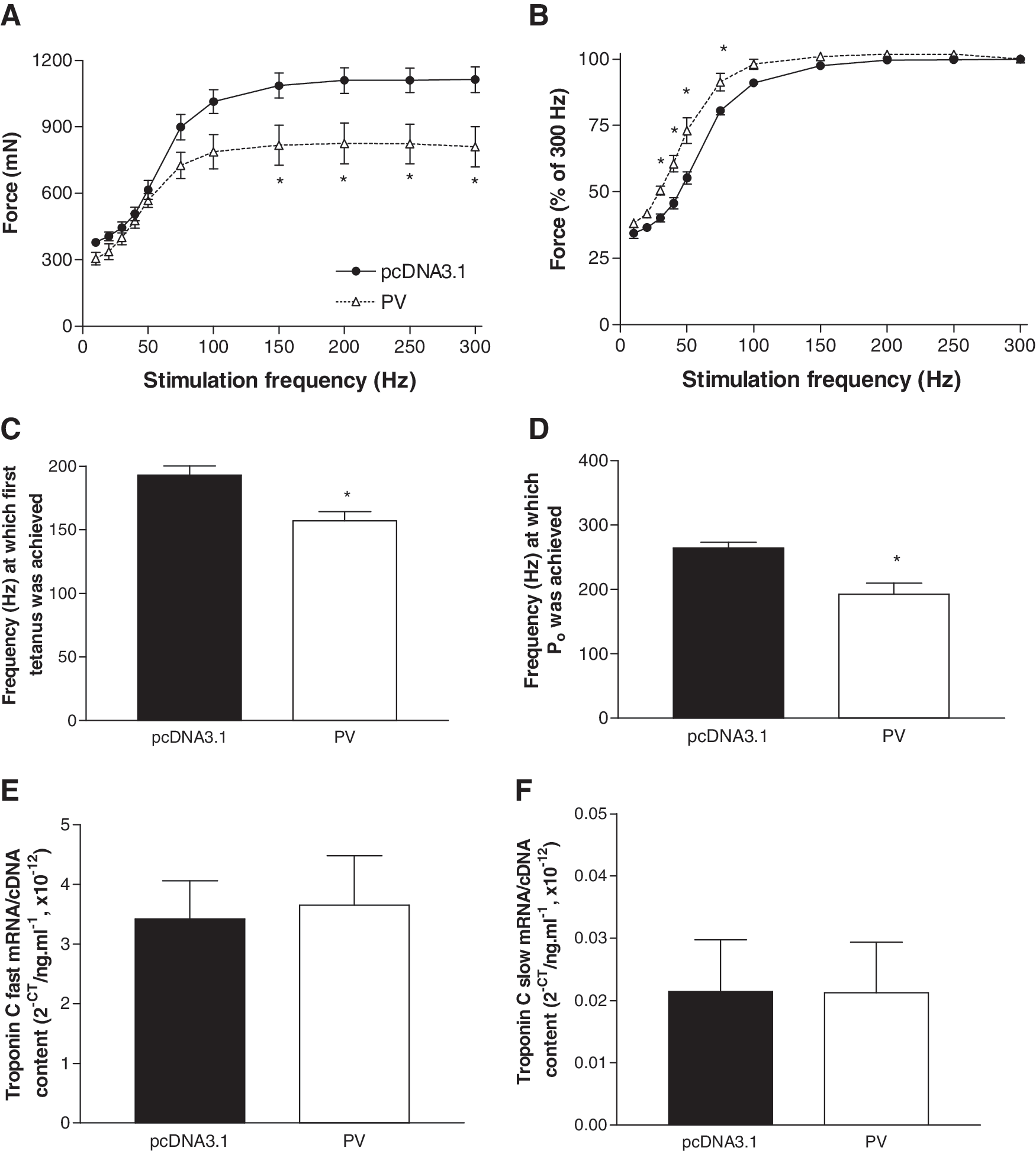
Frequency–force relationship and troponin C mRNA expression in TA muscles of old (26 months) mice 2 weeks after electroporation-assisted transfer of plasmid encoding parvalbumin (PV) or empty vector (pcDNA3.1).
There was no difference between groups in mRNA expression of either the fast isoform (Fig. 6E) or slow isoform of troponin C (Fig. 6F).
Efficacy of PV transfer in vivo in muscles of young and adult mice
Cross-sections of TA muscles from young and adult mice that had been transfected with plasmid encoding PV or empty vector were reacted for PV (Fig. 7A), and quantification revealed a 45% and 37% higher average PV staining (reaction) intensity in muscles transfected with plasmid encoding PV than those transfected with empty vector, respectively (Fig. 7B). TA muscle cross-sections stained for H&E revealed similar muscle fiber architecture, including homogeneity in fiber size, peripherally located nuclei, and the amount of nonmuscle tissue, between treatment groups for both young and adult mice (Fig. 7C).
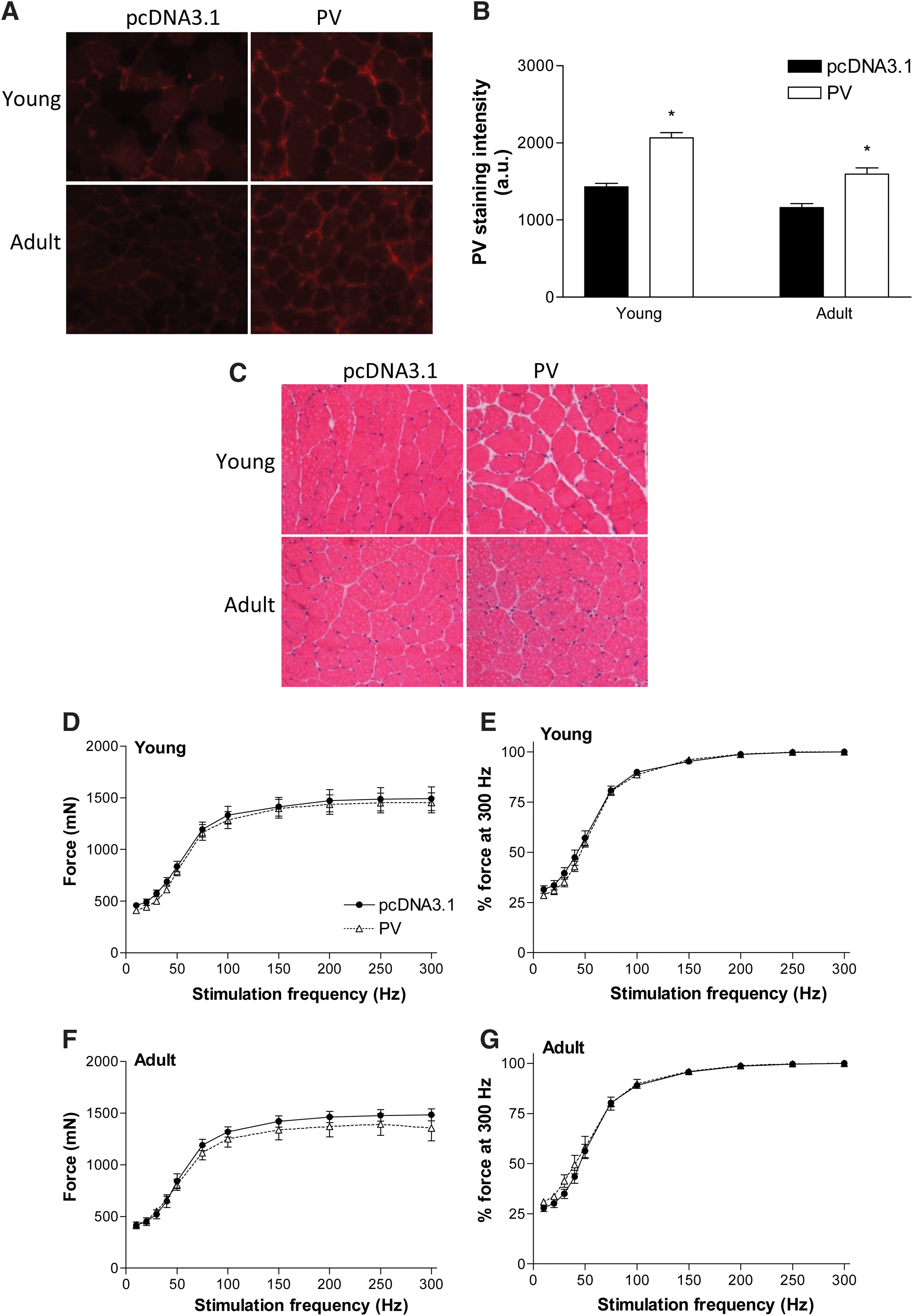
Quantification of PV overexpression and effect on frequency–force relationship in TA muscles of young (6 months) and adult mice (13 months) 2 weeks after electroporation-assisted transfer of plasmid encoding PV or empty vector (pcDNA3.1).
PV gene transfer does not alter muscle mass in young and adult mice
There were no differences between treatment groups in the percentage change in body mass or in the absolute and normalized mass of the right and left TA muscles and the heart in either young or adult mice (Table 1).
PV gene transfer does not alter twitch properties or tetanic force production in situ in young and adult mice
There were no differences in twitch characteristics, peak tetanic force, or specific force of TA muscles from young and adult mice injected with empty vector or plasmid encoding PV (Table 2).
TPT, time-to-peak twitch tension; ½ RT, one-half relaxation time;+dPt/dt, maximal rate of force development; −dPt/dt, maximal rate of relaxation; Pt, peak twitch force; Po, peak tetanic force; sPo, specific (normalized) force. Data are means±SE; n=6–8. No significant differences were found between groups.
Examination of the frequency–force relationship revealed no differences in absolute force or force normalized to that produced at 300 Hz between muscles injected with empty vector or plasmid encoding PV in young (Fig. 7D, E) and adult mice (Fig. 7F, G). There was also no effect of PV gene transfer on the stimulation frequency at which tetanus was first achieved and at which peak tetanic force was achieved in TA muscles of young and adult mice (Table 2).
Discussion
Ameliorating age-related effects on skeletal muscle function has widespread clinical implications for the elderly, including reducing the risk of injury from sudden falls and improving functional independence and overall quality of life (Lynch, 2008). Dysregulation of skeletal muscle [Ca2+]i has been implicated in the deleterious changes in muscle function with aging (Delbono et al., 1995; Wang et al., 2000; Gonzalez et al., 2003; Ryall et al., 2004; Schertzer et al., 2005; Weisleder et al., 2006; Russ et al., 2011). PV is a high-affinity, slow buffer of Ca2+, whose expression decreases in rat fast-twitch skeletal muscle with aging (by 63% in EDL muscles from 8-month-old rats compared with 24-month-old rats) (Cai et al., 2001). Since PV contributes significantly to relaxation rates in fast muscles (Carroll et al., 1997), restoration of PV levels has been proposed as a potential therapeutic strategy to improve Ca2+ handling mechanisms and thus help maintain muscle function during aging (Arif, 2009). We therefore investigated whether increased expression of PV would improve muscle function in aged mice.
Sarcopenia has been suggested to be a condition in which muscle fibers exhibit pathologically elevated [Ca2+]i, similar to that of DMD, and muscles from PV-deficient dystrophic mdx mice (PVKO-mdx) were shown to have lower twitch and tetanic forces than mdx mice (Raymackers et al., 2003). Based on this premise, we hypothesized that increased PV expression would enhance in situ muscle function in old mice. In conditions of normal resting [Ca2+]i, PV binds Ca2+ in competition with Mg2+ and at rest, ∼90% of PV is bound with Mg2+ (Gillis et al., 1982). When Ca2+ is released from the SR, it displaces Mg2+ on PV after the initiation of muscle contraction due to the slow dissociation of Mg2+(Haiech et al., 1979). Due to the relatively slow on/off rates for Ca2+ binding to PV, it was expected that increased PV expression would affect the characteristics of tetanic contractions but only minimally affect twitch contractions.
We found that PV mRNA (−90%) and protein expression (−35%) was lower in TA muscles from old compared with young mice, but using electroporation-assisted gene transfer, we were able to restore PV mRNA and protein expression in old mice to levels comparable with those in young mice. Consistent with published findings (Andersen, 2003), muscles from old mice exhibited a shift in fiber composition to a slower phenotype, with higher MHCI mRNA expression and lower MHCIIx and MHCIIb mRNA expression compared with muscles from adult mice. Since PV is preferentially expressed in fast-twitch fibers (Cai et al., 2001), the reduction in PV expression in old mice is entirely consistent with the shift to a slower muscle phenotype. To our knowledge, age-related changes in PV expression in humans has not been investigated, but since there is a well-documented fiber type shift towards more slow fibers (Andersen, 2003), it would be reasonably expected that muscles from aged humans would also demonstrated reduced PV expression. The discrepancy between PV mRNA and protein expression may be explained by the apparent tight post-translational regulation of PV (Huber and Pette, 1996). Previous studies have found much more rapid changes in PV mRNA expression compared to protein abundance (Huber and Pette, 1996), so it is possible that PV protein abundance may have increased to a similar extent as the gene transcript had we examined muscles another 1–2 weeks after electroporation. Using immunohistochemical techniques, we were also able to show that the overall increase in muscle PV expression with PV gene transfer was attributed to a greater proportion of fibers with high levels of PV staining (reaction) intensity. We have previously optimized the method of electroporation-assisted gene transfer in mouse skeletal muscle to maximize transfer and to minimize muscle fiber damage (Schertzer et al., 2006; Schertzer and Lynch, 2008), and the similarity in mass between the transfected right TA muscle and the contralateral left TA muscle confirm minimal muscle damage with our method. H&E staining revealed similar fiber architecture between muscles transfected with plasmid encoding PV or an empty plasmid, demonstrating that the PV plasmid did not cause muscle damage. The lack of change in cardiac mass and body mass confirmed that PV overexpression was muscle specific and was not deleterious for animal health. Experiments in vitro confirmed that the transfected PV was functional because PV expression in C2C12 cells attenuated the increase in [Ca2+]i induced by an ionophore. Increased PV expression did slow the rate of tetanic force development in muscles of old mice, but contrary to our hypothesis, increased PV expression depressed maximum force in muscles of old mice. These findings are consistent with the binding affinity of Ca2+ being greater for PV than for troponin C (Potter and Gergely, 1975) or the Ca2+-ATPase (Inesi et al., 1980) and suggest that PV overexpression might only provide functional benefit in conditions in which [Ca2+]i is elevated pathologically, as in DMD. For example, resting [Ca2+]i is 275% higher in muscle from boys with DMD compared with muscles from healthy controls (Maunder-Sewry et al., 1980), whereas it is only 23%–75% higher in muscles from old rats compared with young rats (Fraysse et al., 2006). The results are also consistent with studies showing that aging is associated with impairments in excitation–contraction coupling that cause an increased dependence on external Ca2+ for maintaining intracellular Ca2+ and therefore force production (Payne et al., 2004, 2009). As a consequence, therapies aiming to correct the age-related alterations in excitation–contraction coupling may offer better functional benefits that those aiming to increase Ca2+ buffering capacity.
Despite expecting only minimal effects of increased PV expression on twitch characteristics, we found increasing PV expression prolonged the one-half relaxation rate, an observation in contrast to that in previous studies in young rats and mice in which PV overexpression increased the rate of the slow-twitch soleus muscle relaxation (Müntener et al., 1995; Chin et al., 2003). Since PV gene transfer did not affect twitch characteristics in TA muscles of young and adult mice, differences in muscle fiber composition may explain the difference between the studies and requires further investigation.
The fact that increased PV expression caused a leftward shift of the frequency–force relationship and increased relative tetanic force at submaximal stimulation frequencies in muscles of old mice suggests that increased PV expression may enhance the affinity of the contractile apparatus for cytosolic Ca2+. This would be consistent with PV overexpression enhancing Ca2+ buffering, and so an increased affinity of Ca2+ for troponin C may be a protective mechanism to maintain muscle contraction. We found no effect of increased PV levels on the mRNA expression of either the fast or slow isoforms of troponin C.
Although increased PV gene transfer altered the twitch characteristics and reduced tetanic force in muscles from old mice, there was no effect on muscles from young and adult mice. Immunohistochemistry revealed similar increases in PV staining intensity between groups so the differences could not be accounted for by differences in transfection efficiency. These results are consistent with the low [Ca2+]i in muscles from young and adult mice and confirm that increased expression of PV is only deleterious for muscle contractility in conditions of elevated resting [Ca2+]i, such as in sarcopenia.
Our findings that electroporation-assisted gene transfer of PV impaired the force-producing capacity of skeletal muscles from old mice argue against increased PV expression as a therapeutic strategy for sarcopenia and help clarify directions for future therapeutic strategies to attenuate age-related deficits in skeletal muscle structure and function.
Footnotes
Acknowledgments
This study was supported by research grants from the Australian Research Council Discovery-Project funding scheme (DP0665071, DP0772781) and The University of Melbourne Early Career Researcher scheme. The support of the Rebecca L. Cooper Medical Research Foundation is also gratefully acknowledged. K.T. Murphy holds a Career Development Fellowship from the National Health and Medical Research Council (NHMRC).
Author Disclosure Statement
No conflicts of interest, financial or otherwise, are declared by the authors.