
Abstract
RetinoStat® is an equine infectious anemia virus-based lentiviral gene therapy vector that expresses the angiostatic proteins endostatin and angiostatin that is delivered via a subretinal injection for the treatment of the wet form of age-related macular degeneration. We initiated 6-month safety and biodistribution studies in two species; rhesus macaques and Dutch belted rabbits. After subretinal administration of RetinoStat the level of human endostatin and angiostatin proteins in the vitreous of treated rabbit eyes peaked at ∼1 month after dosing and remained elevated for the duration of the study. Regular ocular examinations revealed a mild to moderate transient ocular inflammation that resolved within 1 month of dosing in both species. There were no significant long-term changes in the electroretinograms or intraocular pressure measurements in either rabbits or macaques postdosing compared with the baseline reading in RetinoStat-treated eyes. Histological evaluation did not reveal any structural changes in the eye although there was an infiltration of mononuclear cells in the vitreous, retina, and choroid. No antibodies to any of the RetinoStat vector components or the transgenes could be detected in the serum from either species, and biodistribution analysis demonstrated that the RetinoStat vector was maintained within the ocular compartment. In summary, these studies found RetinoStat to be well tolerated, localized, and capable of persistent expression after subretinal delivery.
Introduction
In wet AMD, it is the abnormal proliferation of blood vessels in the macula (the region providing central vision and sharp resolution) together with the increased permeability of these pathological vessels that causes the progressive deterioration in central vision. The onset of symptoms in patients diagnosed with wet AMD can be sudden, with a person often noticing a sudden change in vision or visual distortion. One study showed that 70% of people with untreated subfoveal choroidal neovascularization (CNV) became legally blind within 9 months of diagnosis (Sickenberg, 2001).
Therapeutic strategies to treat wet AMD involve treatment with anti-vascular endothelial growth factor (VEGF) drugs such as ranibizumab (Lucentis), pegaptanib (Macugen), and aflibercept (VEGF Trap-Eye) that target the abnormally high levels of this specific angiogenic growth factor, which is a predominant factor in supporting the pathological growth of blood vessels. Although these anti-VEGF therapies have proven efficacious for many patients, treatment requires repeated intravitreal injections every 4–6 weeks because of their short half-life in the vitreous, and cessation of treatment often results in reappearance of the disease within a few months. The requirement for these repeated intravitreal eye injections by ophthalmologists is costly and there is an accumulated risk of infection and trauma.
An effective antiangiogenic gene therapy with a long duration of action could offer distinct advantages for the treatment of wet AMD, not least because the key pathologic feature—excessive abnormal neovascularization and edema—appears to be chronic in nature. Previously, gene therapy approaches using an adenoviral vector expressing the angiogenesis inhibitor pigment epithelium-derived factor (PEDF) or small interfering RNA (siRNA) molecules that target the mRNA for VEGF receptor (VEGFR-1) have been evaluated in phase 1 human trials (Campochiaro et al., 2006; Kaiser et al., 2010). Although there were no reported significant adverse events or dose-limiting toxicities with either strategy the benefits of using a nonintegrating adenovirus or an siRNA molecule to treat a long-term chronic condition remain unclear. Alternatively, lentiviral vectors that integrate into the target cell genome give long-term expression, combined with an ability to accommodate large expression cassettes or multiple genes, making them ideal for the development of therapies to treat wet AMD or other chronic diseases in which long-term expression is paramount.
RetinoStat® is based on a nonreplicating nonhuman recombinant lentiviral vector called equine infectious anemia virus (EIAV). The wild-type EIAV causes a self-limiting anemia in horses and is nonpathogenic to humans (Sellon et al., 1992). RetinoStat is delivered subretinally to humans, where it predominantly transduces target retinal pigment epithelial (RPE) cells and ultimately leads to production of the potent angiostatic proteins human endostatin and angiostatin (Balaggan et al., 2006). The reason RetinoStat was designed to express both angiostatic proteins was because of the reported synergistic effects of angiostatin and endostatin on new blood vessel formation (Yokoyama et al., 2000; Scappaticci et al., 2001). Therefore, we hypothesized that expression of two potent angiostatic factors would provide greater efficacy than that measured with either alone. Gene expression for these two therapeutic proteins is regulated through a constitutive cytomegalovirus immediate-early (CMV/IE) promoter (Mitrophanous et al., 1999; Azzouz et al., 2002; Rohll et al., 2002; Bienemann et al., 2003). RetinoStat is pseudotyped with vesicular stomatitis virus G (VSV-G) protein, which facilitates entry into target RPE cells.
In these studies the toxicity, biodistribution, and shedding characteristics of RetinoStat were examined over 6 months after a single subretinal injection in two species: Dutch belted pigmented rabbits and rhesus macaques. The rabbit was selected as the historically preferred species for the assessment of ocular products, largely because of the size of the eye compared with rodents. The rhesus macaque was used because its ocular anatomy is similar to that of the human eye and, importantly, higher primates are the only other animals with a macula and a fovea centralis.
Materials and Methods
Animals
Studies were performed on adult male and female (2- to 7-year-old) rhesus macaques (Macaca mulatta) and adult male and female (5- to 7-month-old) Dutch belted rabbits. All animal studies were done in accordance with U.S. Food and Drug Administration (FDA) Good Laboratory Practice (GLP) regulations.
In both species the right eye was treated with either RetinoStat at a dose of 1.1×105 transducing units per eye (TU/eye) or control buffer (TSSM formulation: tromethamine, NaCl, sucrose, and mannitol (Lamikanra et al., 2005)) in a total volume of 100 μl, while the left eye remained untreated (Table 1).
D, day; M/F, male/female; TU, transducing units.
In the rabbit study equal numbers of male and female rabbits were killed at five interim time points on days 3, 8, 29, 92, and 183. At each time point tissues (ovary, testes, liver, heart, lung, spleen, kidney cortex and medulla, right harderian gland, right lacrimal gland, mandibular lymph node, right upper and lower eyelid, right nictitating membrane, right superior oblique, right ventral rectus, brain [occipital cortex], optic chiasm, right and left optic nerve, right and left retina/choroid, right and left sclera) and body fluids (urine, buffy coat, plasma, cerebrospinal fluid, saliva, and tear swabs) were collected for quantitative polymerase chain reaction (qPCR) analyses of vector presence and periodic blood samples were obtained for analysis of antibody responses to vector in serum. Vitreous fluid samples were also collected at each time point for analysis of the concentration of the therapeutic proteins, endostatin and angiostatin, and for qPCR analysis. Gross pathology and histological examination of a wide variety of tissues was performed on male and female rabbits on day 183 for signs of toxicity, together with urine and blood chemistry analysis.
In the rhesus macaque study, naive male and female animals received either RetinoStat or the control buffer in their right eyes only (Table 1). Samples of buffy coat, plasma, saliva, and tear swabs/eye exudates were collected from each animal on days 2, 3, and 5 for qPCR analysis of vector presence. At the end of the study a full macro- and microscopic examination was performed on a wide variety of tissues. Additional tissue and fluid samples were also collected for determination of vector presence outside the ocular target compartment, and periodic blood sampling was performed throughout the study to examine for possible antibody responses against either RetinoStat vector components or toward the transgenes. A full clinical chemistry and urine analysis was also performed on samples obtained 6 months after RetinoStat administration.
The assessment of toxicity against RetinoStat in both rabbits and macaques was based on mortality, clinical signs, body weight, and qualitative food consumption. In addition, regular in-life assessments of RetinoStat toleration were performed by slit-lamp biomicroscopy, indirect ophthalmoscopy, intraocular pressure (IOP) measurements by Tono-Pen VET (Reichert Inc, of Buffalo, NY) applanation tonometry, full-field electroretinography (ERG), and color fundus photography. Anterior chamber and vitreous cell scores were assigned by using an estimate of cells per field of the focused slit-lamp beam; 0, no cells; 1+, 5–25 cells; 2+, 25–50 cells; 3+, 50–100 cells; 4+, greater than 100 cells. The criteria for the conjunctival hyperemia scores were as follows: 1+, mild; 2–3+, moderate; 4+, severe. RetinoStat biodistribution and shedding were assessed in tear swabs, saliva, plasma, white buffy coat, cerebrospinal fluid (CSF), urine, and vitreous fluid; and in multiple tissue samples collected for qPCR analyses.
Full-field electrophysiology
Animals were anesthetized with ketamine (10 mg/kg) and dexmedetomidine (0.5 mg/kg). Before scotopic testing the animals were dark adapted for at least 2 hr, and afterward the animals were light adapted (80 candela-seconds [cd]/m2) for 10 min before photopic testing. Full-field ERGs were recorded with a UTAS-EMWin visual ERG testing system (LKC Technologies, Gaithersburg, MD). ERG responses were collected with binocular stimulation. Scotopic ERG conditions consisted of a rod-isolating flash (–10 dB blue, 470 nm), a cone stimulus flash (0 dB red, 627 nm), and a standard flash (0 dB white or 2.7 cd/m2, as calibrated by LKC Technologies). Photopic ERG conditions were as follows: standard flash (0 dB white) presented on a rod-saturating background (80 cd/m2), single-flash stimuli, and 30-Hz cone-isolating flicker stimuli. ERG response measures were the average of five presentations separated by 5 sec for the single-flash conditions. For visual evoked cortical potential (VEP) responses, monocular stimulation was applied with the unstimulated eye covered with an opaque occlude. All ERG and VEP conditions were presented with a ganzfeld stimulator and responses recorded from all animals at predose (baseline) and postdose (26 weeks) intervals.
Production of RetinoStat “in the spirit of GMP” by transient transfection
The EIAV lentiviral vector system used to produce RetinoStat consists of a transient transfection of human embryonic kidney 293T cells with three plasmid constructs comprising the vector genome encoding the therapeutic endostatin and angiostatin genes, the codon-optimized gag-pol packaging component, and the VSV-G envelope packaging component (Fig. 1). The RetinoStat vector used for the GLP toxicology studies was produced in a manner analogous to the GMP-grade clinical vector, in which the virus-containing supernatant was harvested and the virus was purified and concentrated by anion-exchange chromatography and hollow fiber technology (patent WO/2009/153563; Truran et al., 2009). Briefly, the vector-containing supernatant is filter-clarified (SartocleanCA pore size of 1.2 μm/0.45 μm filter capsule, Sartorius, Goettingen, Germany). To digest DNA, Benzonase (cat. no. 1.01695.0002; Merck, Darmstadt, Germany) is added to a final concentration of 5 U/ml. After incubation overnight at 2–8°C, the supernatant is applied to an anion-exchange membrane chromatography unit (Sartobind Q Multistep, cat. no. 91-Q-02K-15-06; Sartorius). A stepwise gradient of sodium chloride is passed through the membrane and the vector-containing fraction of the eluate is collected and then concentrated and purified by hollow fiber tangential flow ultrafiltration/diafiltration. During this process the vector medium is exchanged into formulation buffer. The vector material is then filter-sterilized (mini Kleenpak filters, 0.2 μm, cat. no. KA02EKVP2S; Pall, Port Washington, NY) and further concentrated by hollow fiber ultrafiltration. The RetinoStat batch used in these studies was labeled IH39 and the titer was ascertained by integration (DNA) titer assay as 1.1×107 TU/ml, using the method previously described (Stewart et al., 2009).
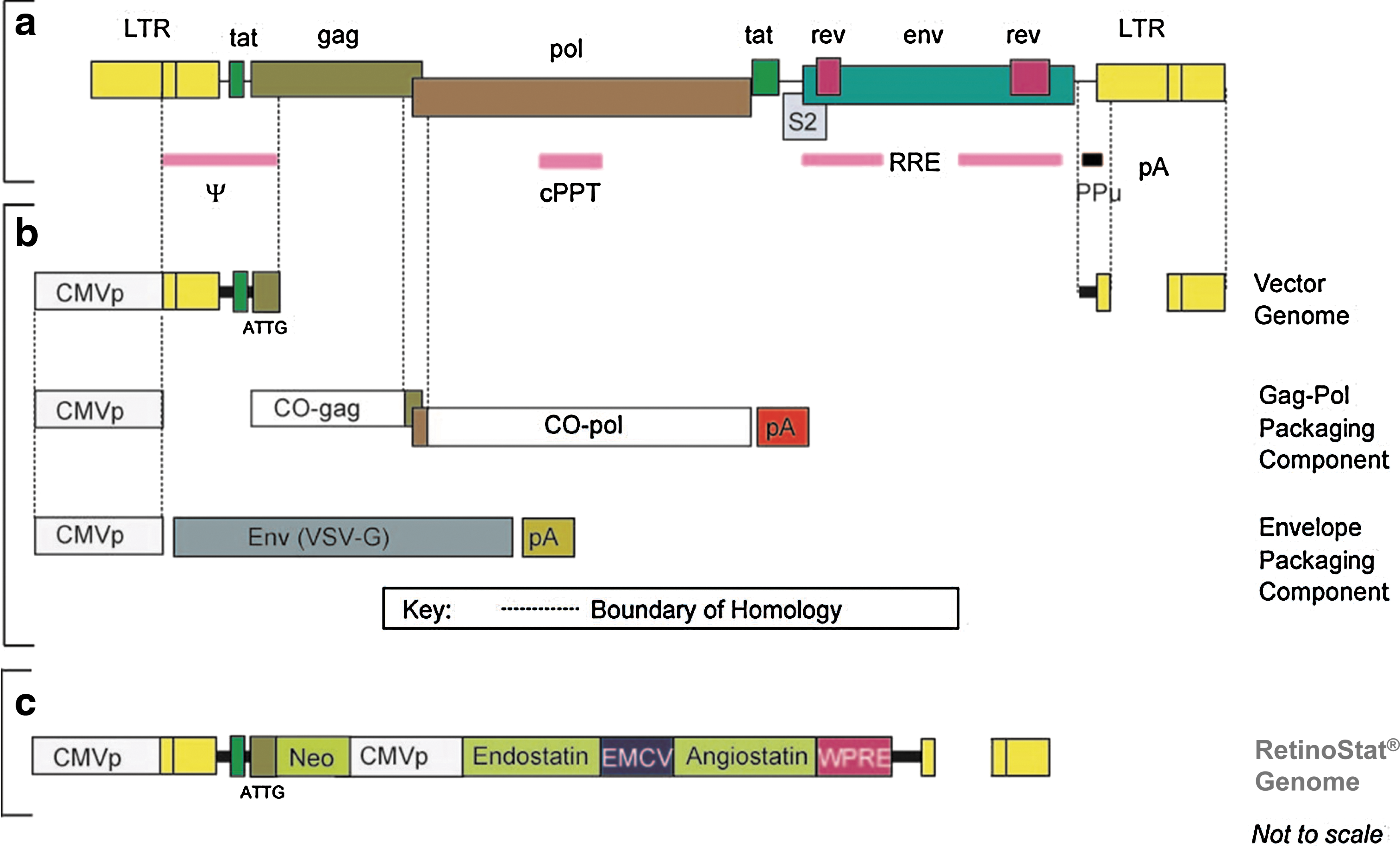
Schematic diagram showing the genetic structure of
Product enhanced reverse transcriptase (PERT), predicted vector titer, and RNA copy number (concentration) assays were also carried out on RetinoStat IH39. The PERT assay is an indirect method for quantifying vector particles by measuring the amount of RT activity within vector preparations (Rohll et al., 2002). The RNA copy number RT-PCR assay quantifies the number of genomic RNA molecules within a given vector preparation by using qRT-PCR technology and oligonucleotides specific to the EIAV packaging (ψ) signal sequence (Rohll et al., 2002). PERT and RNA copy number for IH39 were 3.7×107 TU/ml PERT predicted titer and 1.5×1012 RNA copies/ml, respectively.
Additional safety- and purity-specific assessments were carried out on RetinoStat IH39 to ensure that the vector was suitable for use in the GLP rabbit and macaque toxicology studies. These assessments included confirming sterility, mycoplasma-free status, endotoxin (<15 IU/ml), pH (7.0–7.5), residual bovine serum albumin (BSA), and total protein and residual DNA content (PicoGreen dsDNA; Life Technologies, Paisley, UK). In addition, the identity of the RetinoStat vector in the IH39 batch used in the GLP studies was confirmed in a PCR-based assay employing primers specific for regions of the RetinoStat RNA sequence, and potency was determined by measuring the level of endostatin and angiostatin after in vitro transduction.
The assay to detect replication-competent lentivirus (RCL) is not routinely carried out on nonclinical vector preparations, such as RetinoStat IH39, that are produced in the “spirit of GMP.” This is because the manufacturing methodology and vector engineering minimize the risk of RCL formation, and EIAV replication is restricted by the absence of essential cis-acting sequences and regulatory proteins in the host species used in these studies. However, in accordance with regulatory requirements, the RCL assay has been used to confirm that no RCL are detected in any GMP clinical batches of RetinoStat (Miskin et al., 2006). Additional tests specifically required for clinical use, or for product characterization, are carried out on GMP batches of RetinoStat for clinical use but were not carried out on batch IH39.
Subretinal injection
Subretinal injections of RetinoStat or control formulation buffer were performed in the right eye of each animal, using the RetinaJect apparatus (SurModics, Eden Prairie, MN). A wide-angle contact lens viewing system was employed for the rabbits in order to better visualize the medullary ray. Rabbit subretinal injections were made 1 or 2 disc diameters nasal to the optic nerve at the level of the medullary ray. The blebs typically extended from the medullary ray inferiorly to include part of the visual streak.
A plano-convex lens was used to visualize the macula in the macaque eyes. A 25-gauge fiberoptic light was passed into the vitreous cavity via a 25-gauge cannula to better illuminate the retina of the heavily pigmented macaque eyes. The RetinaJect was passed through the conjunctiva and sclera 2.5 mm posterior to the superotemporal limbus. The 25-gauge cannula for the fiberoptic light was passed 2.5 mm posterior to the superonasal limbus. The 100-μl retinal injections were made along the superotemporal macula near a retinal vessel. Most of the blebs extended inferiorly and often included the fovea. The 25-gauge needle sclerotomies were self-sealing, and suturing was not required in either rabbits or macaques.
Although the RetinaJect is a nonclinical device that can be used only for nonhuman studies, it has been shown that subretinal delivery is comparable whether the RetinoStat vector is administered by RetinaJect or the device currently in clinical use, which consists of a 41-gauge needle/cannula tubing apparatus. It has also been demonstrated that the RetinoStat vector is compatible with both the RetinaJect and clinical devices (data not shown).
Endostatin and angiostatin ELISAs
Biochemical measurements of human endostatin and angiostatin protein levels in rabbit vitreal fluid samples were performed with a commercially available endostatin enzyme-linked immunosorbent assay (ELISA) (cat. no. DY1098; R&D Systems, Minneapolis, MN) and an angiostatin ELISA employing a recombinant human angiostatin standard (cat. no. ABC2510; Autogen Bioclear UK, Calne, UK), an anti-human angiostatin capture antibody (cat. no. AF226; R&D Systems), and a horseradish peroxidase (HRP)-conjugated goat anti-human plasminogen antibody (cat. no. CL20085APHP; Cedarlane Laboratories, Burlington, ON, Canada). The limit of detection for each assay was 3.12 and 1.95 ng/ml for endostatin and angiostatin, respectively.
Immunogenicity assay
Antibody responses against the RetinoStat vector, including the envelope protein (VSV-G) and the transgene products angiostatin and endostatin and other EIAV vector components, were assessed by semiquantitative ELISA in rabbit serum samples and by Western blotting in rhesus macaque serum samples.
Rabbit anti-VSV-G peptide antibody was used to construct an ELISA standard curve (cat. no. V4888; Sigma-Aldrich, St. Louis, MO). For Western blotting, cell lysates prepared from the negative and RetinoStat-positive control cell pellets were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) on 12% Tris–glycine gels, blotted onto polyvinylidene difluoride (PVDF) and incubated with either rhesus macaque test or control serum samples. After washing, the membrane was incubated with an appropriate HRP-conjugated secondary antibody.
Vector PCR
Real-time PCR analysis of biological samples from the macaque and rabbit toxicology studies was used to measure vector persistence, biodistribution, and shedding after a single subretinal administration of RetinoStat.
DNA was isolated from buffy coat samples, using a Qiagen DNA mini kit (Qiagen, West Sussex, UK). DNA was isolated from tissue samples with a MagNA pure LC DNA isolation kit II (03186229001; Roche). The amount of DNA analyzed per replicate qPCR was up to 1000 ng. A plasmid DNA assay standard containing the EIAV ψ signal was used to generate a standard curve (Rohll et al., 2002). The qPCR mix consisted of EIAV ψ primers and probes as described previously (Stewart et al., 2009). The lower limit of quantification (LLOQ) was defined in each assay on the basis of the standards.
RNA was isolated from various fluid samples (including left and right eye vitreous fluid, plasma, urine, CSF, swabs of saliva, and left and right eye tears), using a QIAamp viral RNA mini kit (Qiagen). The extracted RNA samples were analyzed by reverse transcription-PCR (qRT-PCR) at a maximal volume of 8 μl per 25-μl reaction. An in vitro cRNA transcript containing the EIAV ψ signal was used to generate a standard curve (Rohll et al., 2002). The qRT-PCR mix consisted of EIAV ψ primers and probes as described previously (Stewart et al., 2009). The LLOQ was defined in each assay on the basis of the standards. Average copy numbers per cell were estimated by dividing the mean number of vector sequences detected by the predicted number of cells in 1 μg of genomic DNA (1.47×105 cells).
Nucleic acid isolations from test samples were performed alongside buffer-only controls; no-template control reactions were set up at the PCR stage to monitor for cross-contamination. For the DNA PCR analysis known quantities of an EIAV genome plasmid (called pONY8.0Z) was used at a range of copy numbers between 10 and 1×106 copies per reaction to construct a standard curve. To monitor for PCR inhibition, test DNA samples were spiked with 1000 copies of the pONY8.0Z plasmid and assayed in a PCR. For viral RNA PCR analysis, an in vitro transcript cRNA standard was used at a range of copy numbers between 100 and 1×106 copies per reaction to construct a standard curve. A cRNA spike (1×104 copies per reaction) was also used to monitor PCR inhibition in the test samples. In addition, all samples were amplified in parallel in the absence of reverse transcriptase to detect any contaminating EIAV ψ DNA in the viral RNA samples.
Results
Six-month toxicology, biodistribution, shedding, and immunogenicity studies
The studies performed in macaques and rabbits were designed to examine ocular tolerance of subretinally dosed RetinoStat vector (see Table 1).
Clinical assessments
There were no unscheduled deaths that were related to subretinal administration of RetinoStat or control buffer in either species, nor were there any statistically significant differences in body weights and relative organ weights at the scheduled necropsy over the 6 months in rabbits or macaques treated with RetinoStat as compared with control animals dosed subretinally with buffer. No treatment-related changes in blood chemistry were observed in the RetinoStat-treated animals at any of the sampling time points, nor was there any evidence of changes in red cell or white cell counts. RetinoStat did not change blood clotting times, and there were no significant treatment-related microscopic observations in nonocular tissues in either rabbits or macaques.
Clinical ophthalmic observations
Subretinal injections typically resulted in ophthalmoscopically visible small retinal holes at the injection site as well as a well-developed subretinal bleb in all treated right eyes in both species. The retinal holes sealed quickly in both rhesus macaque and rabbit eyes by day 3 and day 8, respectively. The retinal elevation had resolved in all animals by day 8 and day 22 in macaque and rabbit eyes, respectively, injected with RetinoStat. In two rabbits injected with control buffer, large retinal holes were observed at the point of maximal retinal detachment, and retinal thinning was also observed in some RetinoStat- and control buffer-treated rabbits. Neither of these findings was observed in the macaques. The exact cause and functional significance of the rabbit-specific findings were unclear, but it may be due to the nature of the retinal blood supply in rabbits, in which only part of the inner retina is supplied by blood vessels (merangiotic) which renders this tissue more susceptible to hypoxia than the fully vascularized (holangiotic) retina in macaques and humans (Schwab, 2011).
On slit-lamp biomicroscopy there was a transient rise in vitreal chamber cell score in both macaque and rabbit eyes, which peaked within 1 week of the dosing phase (Fig. 2). Importantly, similar inflammatory cellular responses were observed in control buffer-treated eyes, except that the frequency and duration of the cellular response were slightly greater in the RetinoStat-treated animals. The mean vitreal cell score was “mild” (1+) to “moderate” (2+) in the RetinoStat-treated rabbit and macaques, respectively, 1 week after dosing and rapidly decreased to “trace” levels by 3 weeks. Anterior chamber (aqueous cell) inflammatory scores were “mild” (1+) and transient, and disappeared within 1 week of vector administration in both species treated with either vector of control buffer. Mild to moderate (1+ to 2+) conjunctival hyperemia and focal swelling at the injection site were noted in rabbits and macaques of both the RetinoStat and control buffer groups on day 3 after injection (Fig. 2). This was attributed to the preparation of the ocular surface, the injection itself, and the mild intraocular inflammation. These observations were transient and resolved by day 8 of the dosing phase.
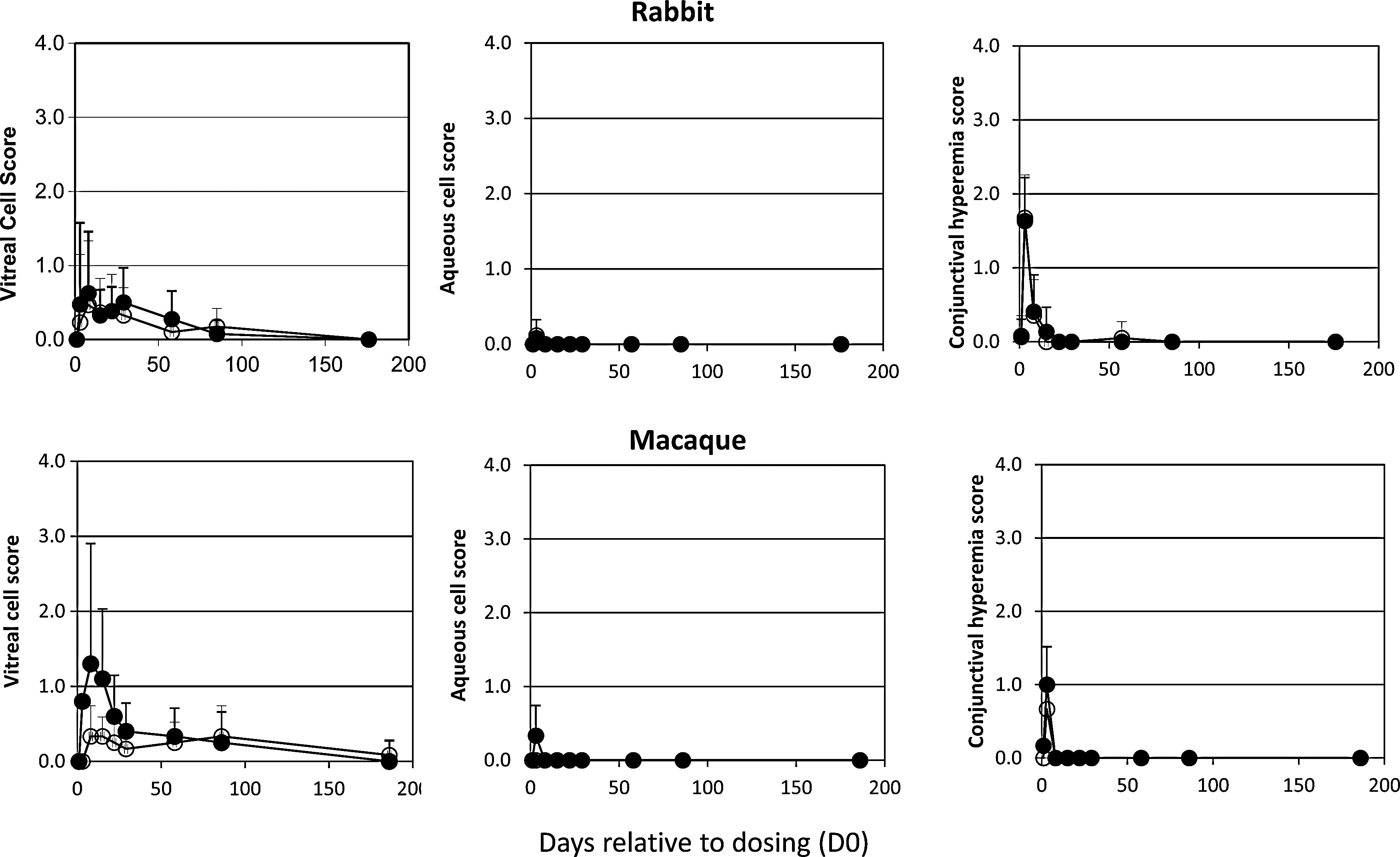
In vivo ocular inflammatory scores in macaques and rabbits. Inflammatory changes were assessed by slit-lamp biomicroscopy and indirect ophthalmoscopy. Solid circles, RetinoStat treated; open circles, buffer treated. Clinical changes were graded at the following levels of severity: 1+, mild; 2–3+, moderate; 4+, severe. The mean score at each time point for each group is shown±standard deviation.
Intraocular pressure (IOP) measurements remained within the normal range for both species (Fig. 3) throughout the studies. The slight transient reduction in IOP in vector-treated eyes, versus control buffer-treated eyes, in rabbits and macaques was attributed to mild intraocular inflammation, but was not significantly different between the contralateral eyes of either species. Electroretinography (ERG) was performed on all animals in both studies before dosing and then at 6 months to assess whether there were any statistically significant treatment-related changes between A wave and B wave amplitude and latency recordings in response to different stimuli (Supplementary Table S1; supplementary data are available online at
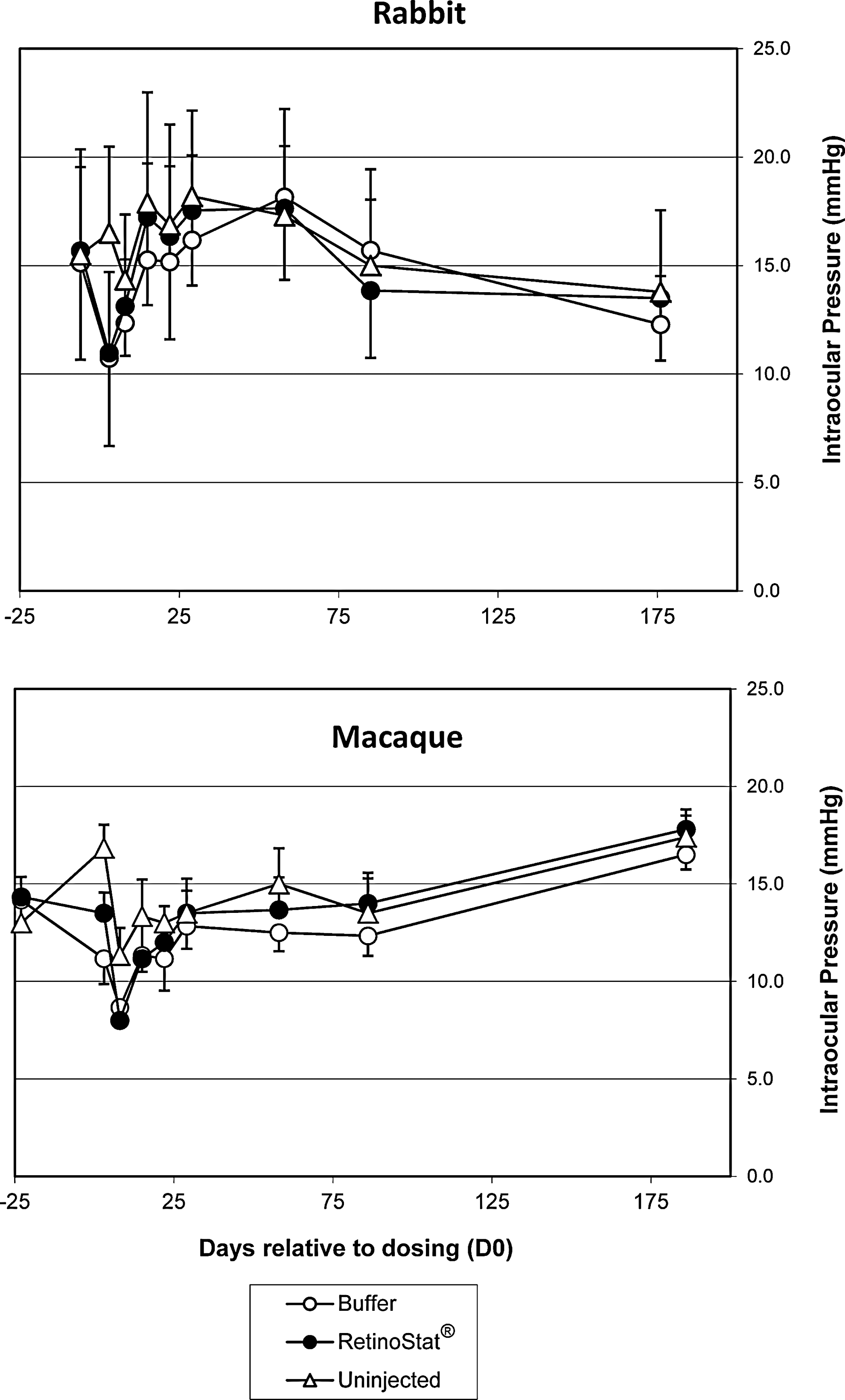
Intraocular pressure measurements. The mean intraocular pressure (IOP) measurements at each time point for each group are shown±standard deviation.
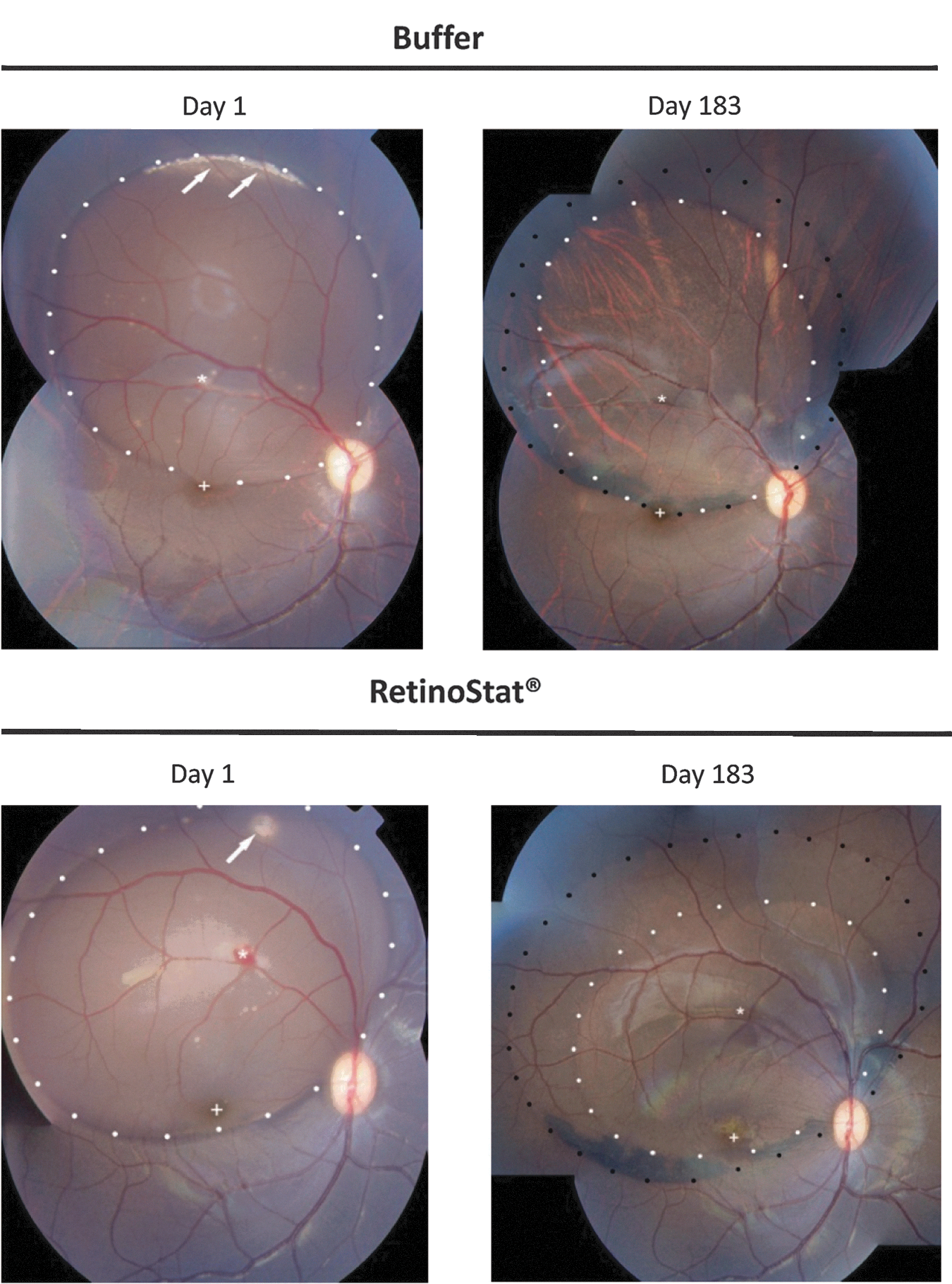
The location and size of the subretinal bleb were deduced on the basis of fundus color photographs in both rabbit and macaque eyes after administration of either RetinoStat or buffer on day 1 (Fig. 4). Administration of either RetinoStat or control buffer produced a mottling pigmentation change within the RPE that was limited to the subretinal bleb area from day 3 to 6 months in both rabbits and macaques. Over time the RPE mottling extended beyond the initial bleb site. This was thought to be due to spreading out of the bleb over the first few days after subretinal administration and was visible as a “double ring” on the retinal photograph at the end of the study. Some minor evidence of retinal injury, atrophic holes, loss of ganglion cells, and subretinal infiltrates was seen in both RetinoStat and control buffer-treated rabbit eyes, but this was attributed to the subretinal procedure (Fig. 5). A few small neovascular tufts developed on the inner surface of the rabbit retina at the inferior edge of the medullary ray in the region of the bleb in approximately one-third of the RetinoStat-treated rabbit eyes, 3 and 6 months after dosing, but this was not associated with any other pathological findings or changes in ERG recordings. This phenomenon did not increase in frequency or magnitude over time from 3 to 6 months. Unlike the holangiotic macaque retina, the rabbit retina is merangiotic and, thus, more prone to ischemic changes during the brief period of retinal detachment, such as atrophic holes and neovascularization.
Representative macaque retinal photography images. On day 1 the retinal image was taken after the subretinal procedure. The white dots delimit the bleb of subretinal fluid. Fovea (+) and injection site (*) are shown. Small air bubbles are also highlighted by arrows. Retinal pigment epithelial mottling is evident from day 8, extending beyond the region of the original bleb (white dots) to form a second ring (black dots) by day 183.
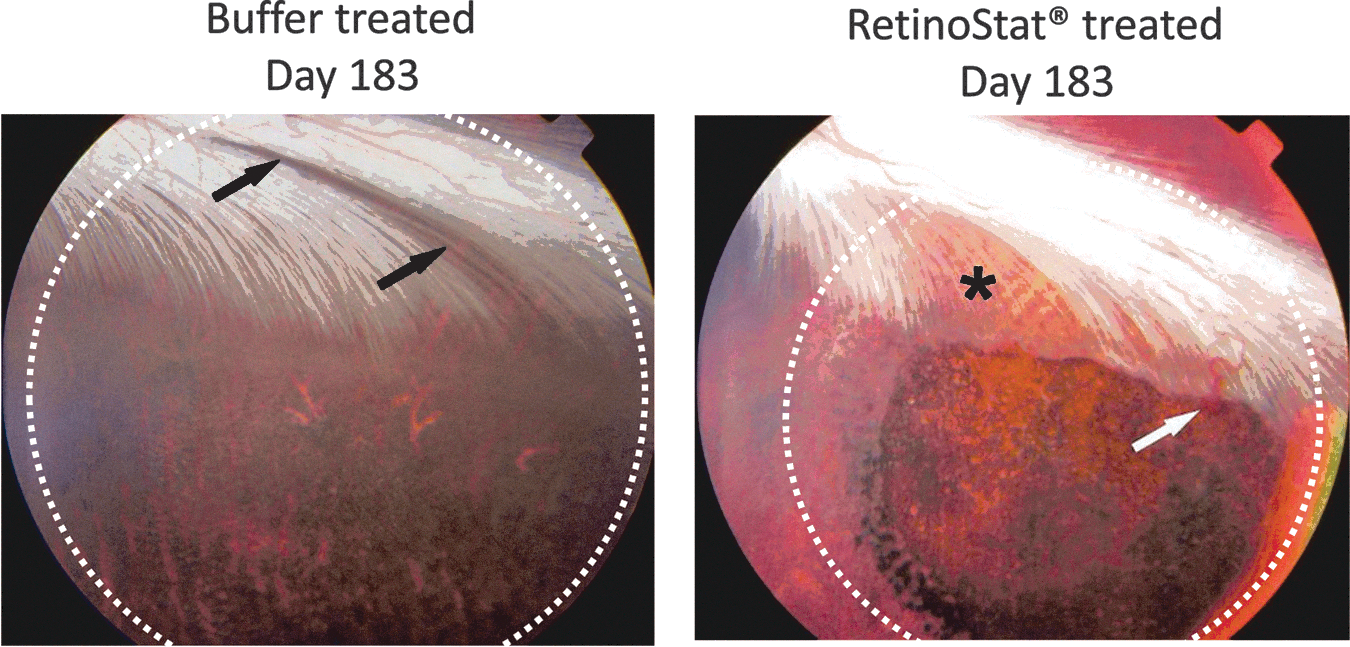
Rabbit retinal photography images from TSSM buffer control-treated or RetinoStat-treated eyes on day 183. The region of the subretinal bleb (white dots) is shown. The white arrow is pointing toward a single retinal “neovascular tuft” that were present in 2 of 14 RetinoStat-treated rabbit eyes on day 183. The black arrows show loss of myelin from the medullary ray within the area of the subretinal bleb in a TSSM buffer-treated rabbit eye. The black asterisk shows a more diffuse area of myelin loss in the RetinoStat-treated eye.
Ocular histology
All microscopic findings including minimal to slight segmental hyperpigmentation/hypertrophy/hyperplasia of the RPE, retinal edema, mononuclear infiltration, and slight axon degeneration were observed in both RetinoStat- and control buffer-treated macaque (Fig. 6) and rabbit eyes (data not shown). The findings were localized to the subretinal injection site and were considered an effect of the injection procedure and delivery of a subretinal volume. The small neovascular tufts that developed in some of the RetinoStat-treated rabbit eyes could not be identified histologically because serial sections were not performed.
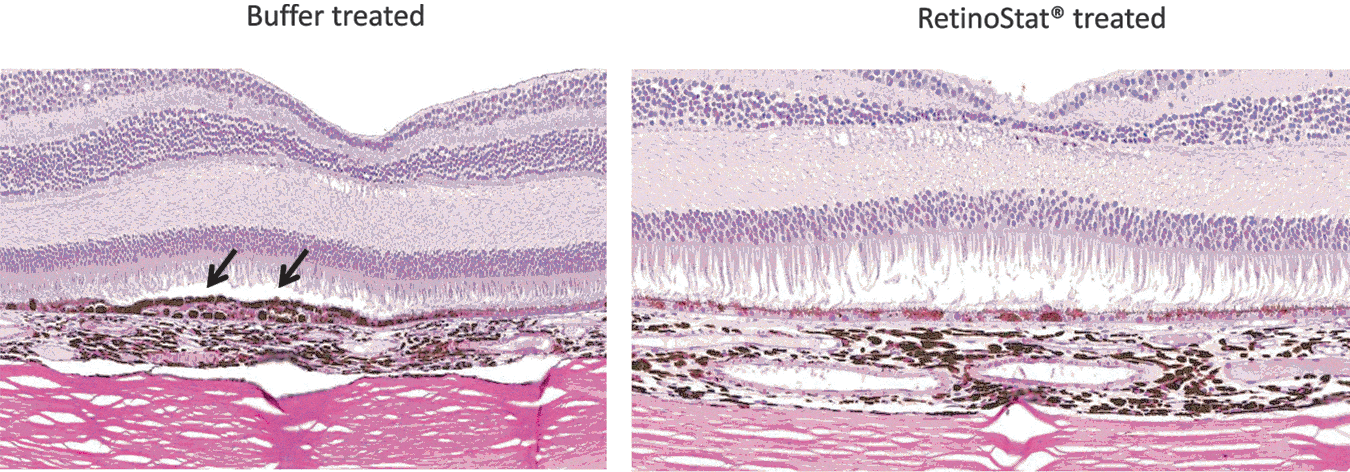
Macaque retinal cross-sections. The day 183 macaque retinal cross-sections show irregular hypertrophy and hyperpigmentation of the retinal pigment epithelium (RPE) in the region of the subretinal bleb in both RetinoStat- and control buffer-treated eyes. In addition to the hypertrophy and hyperpigmentation, RPE hyperplasia was also observed in some eyes from both groups and is highlighted (black arrows) in the control buffer-treated retina.
Biodistribution, persistence, and shedding of RetinoStat
Each RetinoStat vector particle contains two copies of the genome as positive-strand mRNA. Once the vector enters cells it undergoes reverse transcription and is copied into double-stranded DNA, which integrates into the target cell chromosomes. By measuring the amount of RetinoStat RNA (RetinoStat vector particles) or DNA (RetinoStat in cells) an estimate of vector distribution can be made. In the 6-month rabbit toxicology study samples from various fluids, tissues, and buffy coat were analyzed by qPCR at selected time points to determine the biodistribution, persistence, and shedding of the RetinoStat vector after subretinal delivery. Vector particle dissemination and/or persistence were assessed by analyzing RNA extracted from plasma and cerebrospinal fluid (CSF) samples for the presence of RetinoStat-derived EIAV Ψ sequences. The same qRT-PCR assay was used to assess vector shedding from RNA extracted from swabs of saliva and tears, urine, and RetinoStat-treated right eye vitreous fluid. Biodistribution was assessed by analyzing DNA extracted from various tissues and buffy coat samples for the presence of RetinoStat-derived EIAV packaging signal (Ψ) sequences.
There was no evidence of RetinoStat vector particles in rabbit urine or saliva on day 2. Similarly, RetinoStat vector particles were not detected in the majority of rabbit tear swabs on days 2, 3, and 5 of the dosing phase and when detected, levels were below the lower limit of quantification (<LLOQ) for the assay. RetinoStat vector particles were detected in the vitreous fluid from the vector-treated rabbit eyes in five of six animals on day 3 of the dosing phase, but by day 29 this had fallen to one of six vitreous samples (<LLOQ). Vector particles were detected in the vitreous fluid of two of six contralateral untreated rabbit eyes on day 3, but all were below the LLOQ and were not detected at any later time points. No dissemination of vector particles was observed in plasma or cerebrospinal fluid at any time point in the rabbit study.
In the rabbit tissue samples the highest concentrations of vector DNA was located in the target retina/choroid and the sclera and was completely absent in nonocular tissues by 6 months (Fig. 7A). The presence of RetinoStat in the sclera was concluded to be due to difficulties in separating the sclera from the overlying retinal/choroid layers, leaving some cross-tissue contamination because the sclera had <5% of the copies present in the retina/choroid. Sporadic positive signals for RetinoStat vector were observed at early time points (buffy coat, right optic nerve, superior oblique, nictitating membrane, lower right eyelid, right harderian gland, liver, left eye retina, and ovary) but only in a minority of rabbits and never above the LLOQ for the assay; signal was undetected in all of these sample types by the end of the study on day 183. No vector was detected in testes, heart, lung, spleen, kidney cortex, kidney medulla, lacrimal gland, mandibular lymph node, upper right eyelid, ventral rectus, optic chiasm, brain, left eye sclera, and left optic nerve of the rabbits. No vector sequences were detected above the LLOQ in the left uninjected eye at any time point, indicating that vector was unable to cross into the contralateral eye after the single subretinal delivery to the right eye.
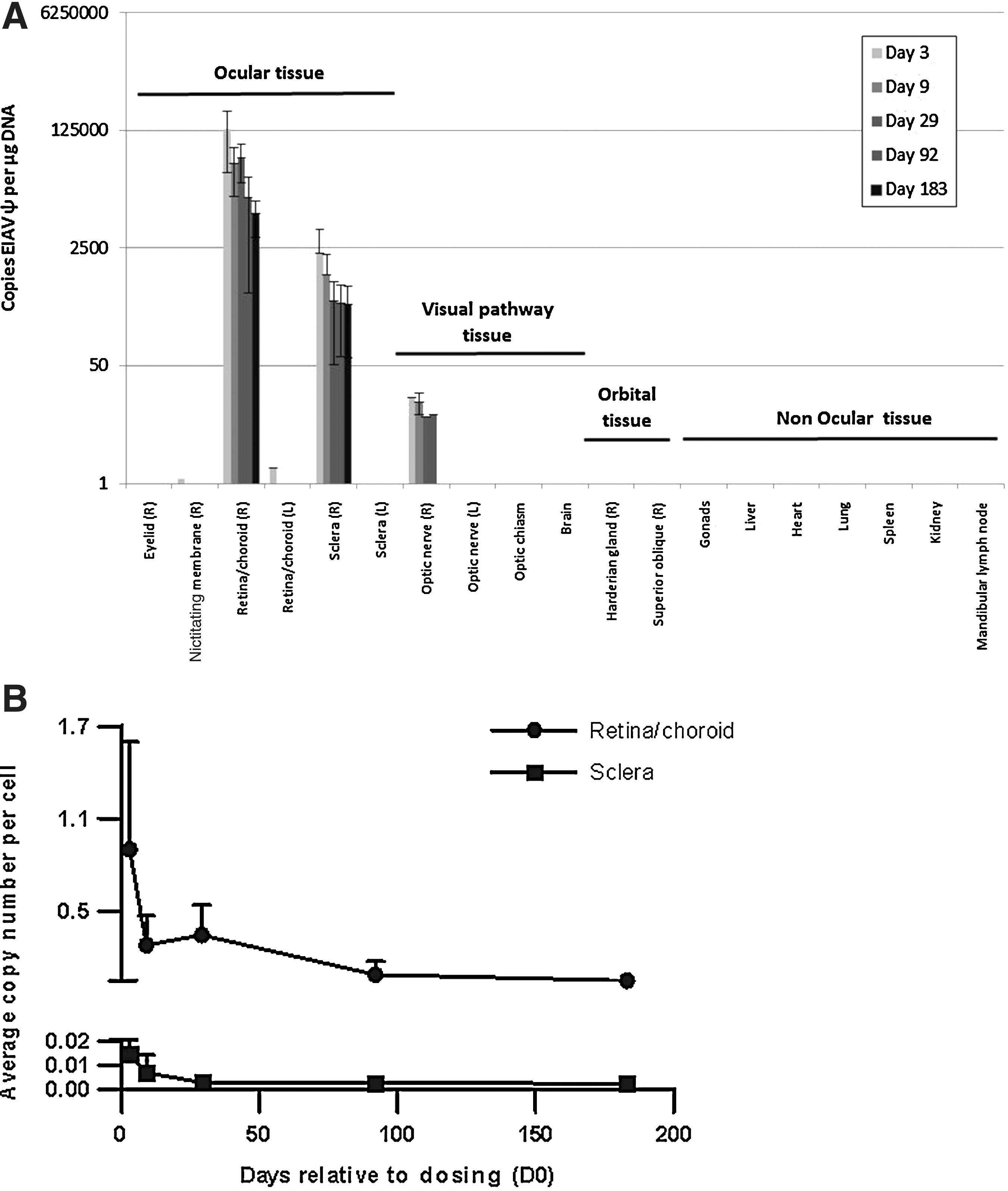
Biodistribution of RetinoStat in rabbits.
In the macaque study regular body fluid sampling showed that RetinoStat vector particles were detected only in a minority of animals on day 2 (saliva swabs, right eye tear swabs; all <LLOQ), but were undetected in plasma, and were absent by days 3 to 5. In the biodistribution component of the macaque study RetinoStat vector DNA was detected only in one of the buffy coat samples on day 2 at a level that was less than the LLOQ; this signal was absent by day 5. At the end of the study RetinoStat vector particles were not detected in urine or CSF and RetinoStat vector DNA was not detected above the LLOQ in ovary, testis, liver, heart, lung, spleen, kidney cortex, kidney medulla, brain, right lacrimal gland, right lower/upper eyelids, ventral rectus, optic chiasm, right optic nerve except for superior oblique sample, and mandibular lymph node.
In the target retina/choroid the average copy number of RetinoStat-specific sequences fell from a mean of 0.9 copies per cell to 0.05 copies per cell by 6 months (Fig. 7B), a reduction of 18-fold. A similar pattern was seen in the sclera samples albeit at a lower overall level of 0.01 copies per cell on day 3, falling to 0.002 copies per cell by 6 months (Fig. 7B). These numbers represent an average, as when the retina was dissected it may not exactly correlate with the region of the subretinal bleb.
Expression of human endostatin and angiostatin
Concentrations of human endostatin and angiostatin proteins in the RetinoStat-treated rabbit eyes rose to a peak on day 29 with means of 539±232 ng/ml for human endostatin and 391±127 ng/ml for angiostatin (Fig. 8). The levels progressively decreased from day 92 to day 183, at which time the mean levels of endostatin and angiostatin were 306±126 and 184±89 ng/ml, respectively. Although there was a statistically significant 2-fold reduction in vitreous angiostatin concentrations (p>0.05) there was no statistically significant reduction in vitreous endostatin concentration over time. The slight drop in angiostatin concentration may be due to the loss of unintegrated EIAV species over the course of the 6-month study. Unintegrated species do not express as well as integrated species, and therefore a 10-fold drop in these may lead to a smaller drop in overall protein expression levels (Wanisch and Yáñez-Muñoz, 2009).
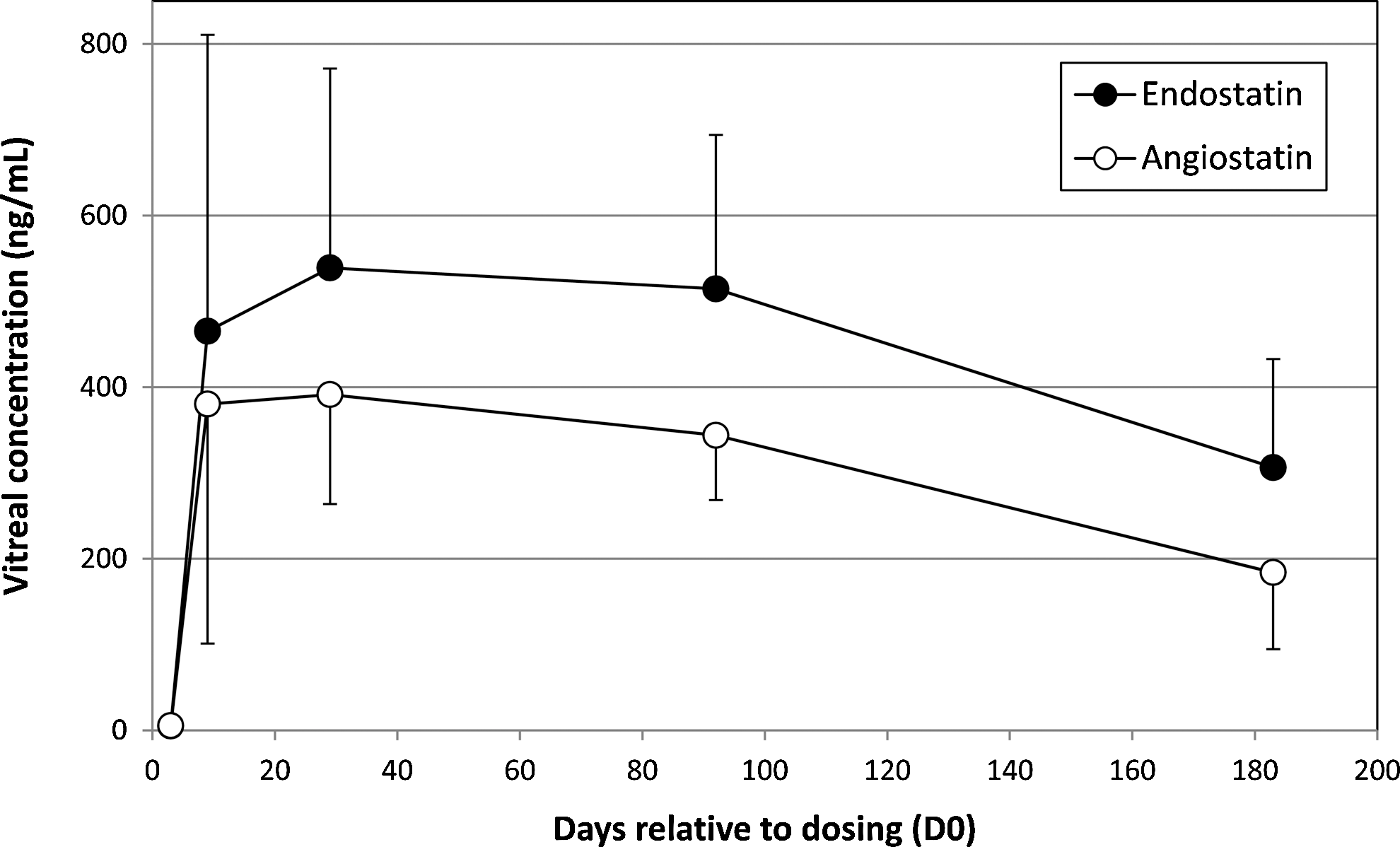
Expression of human endostatin and angiostatin in the rabbit vitreous humor. The vitreous humor harvested at necropsy from the RetinoStat-injected (right) eye of the rabbits on days 3, 9, 29, 92, and 183 after dosing of the vector. The vitreous was assayed for the level of endostatin and angiostatin via ELISAs.
Immunogenicity
All the rabbit serum samples tested negative for antibodies against envelope protein (VSV-G), the human transgenes angiostatin and endostatin, EIAV components, neomycin phosphotransferase (Neo PT), and other RetinoStat components, as measured by semiquantitative ELISA (data not shown). Samples of macaque serum obtained before administration of either RetinoStat or control buffer revealed an antibody response against HEK293T cell-associated antigens by Western blotting. This was attributed to prior exposure of the animals to similar antigens before the study was initiated and was not considered a treatment-related phenomenon. Antibody responses against the RetinoStat envelope protein, VSV-G were not detected in any serum samples collected at any time point and no other humoral responses were generated against the human transgenes angiostatin and endostatin or the EIAV vector components.
Discussion
Tolerance and safety of subretinally injected RetinoStat were investigated in both rabbits and macaques. The studies were designed to examine the consequences of long-term gene integration and transgene expression from RetinoStat in the eye and to determine the biodistribution, shedding, persistence of the vector, and immunogenicity after subretinal administration.
After subretinal delivery of the RetinoStat vector at a dose of 1.1×105 TU/eye there was an initial mild to moderate cellular inflammatory response in both species that persisted for approximately 2–3 weeks. This cellular inflammation occurred to a slightly lesser degree in the control animals treated with buffer. The finding of ocular inflammation was not unexpected, as other preclinical studies using subretinal delivery to the eye have observed similar mild to moderate ocular inflammation (Jacobson et al., 2006; Lai et al., 2011). In terms of the ocular inflammatory response subretinally delivered vector appears to be better tolerated than those delivered intravitreally (Husain et al., 2005; Macclachlan et al., 2011). For example, intravitreal administration of an adeno-associated virus type 2 (AAV2)-based vector in monkeys induced a mild to moderate inflammation that in some cases persisted for up to 15 months. Similarly, clinical studies investigating ranibizumab, an anti-VEGF antibody treatment delivered intravitreally for treating AMD, observed a moderate short-term inflammatory response in both the anterior and posterior portions of treated eyes (Rasmussen et al., 2003; Brown et al., 2006; Rosenfeld et al., 2006).
Although no RetinoStat-related toxicities were observed in either species, small neovascular tufts on the inner surface of the retina were noted in a minority of RetinoStat-treated rabbit eyes, but not in control rabbit eyes or in the macaques. The reason for the development of these neovascular tufts remains unclear but it may be related to the proteinaceous nature of the RetinoStat vector compared with the control buffer and the anatomy of the rabbit retina, which is quite different from that of primates. Unlike the primate retina, which is fully vascularized, the rabbit retina is only partially vascularized and the oxygen demands of the metabolically active visual streak where the RetinoStat injections were placed are only just met in the normal eye (Yu and Cringle, 2004). Therefore, if the reabsorption of the retinal detachment is protracted by the proteinaceous RetinoStat vector this could lead to differential changes in local oxygen supply and demand and therefore drive new vessel production in an attempt to improve the oxygen supply to the retina. In support of this hypothesis, the neovascular tufts were located at the inferonasal edge of the medullary ray, which was located within the region of the bleb where the risk of a short-term anoxic insult would be most likely to occur.
In contrast, primate retinas have a remarkable ability to tolerate even total retinal detachments for days, weeks, or even a few months because even though there is some ischemia in chronic retinal detachment in humans, retinal neovascularization is seldom observed. For example, it has been shown that subretinal blebs of balanced salt solution are well tolerated in the macaque eye (Nork et al., 2012). The choroid is responsible for the retinal oxygen supply in primates; however, during a detachment the photoreceptors can go into a state of reduced metabolism and, thus, require less oxygen. Thus, because of the differences in retinal structure between the rabbit and the primate eye a retinal detachment is not predicted to have a significant impact on “neovascular tuft” formation in the vector-administered primate eyes.
Another consequence of the subretinal procedure is mottling of the RPE that occurred in both species treated with either RetinoStat or control buffer. This phenomenon readily occurs when the neurosensory retina is separated from the RPE due to hyperpigmentation, hypertrophy, and hypopigmentation of the melanin granules within the RPE, and can readily be used microscopically to determine whether a retinal detachment is real or an artifact of the processing. In these studies RPE mottling was evident in the fundus photographs and in the histology sections taken through the retina at the level of the subretinal bleb area after dosing in both species. Normally these changes occur within hours of retinal elevation, and if the process of “reattachment” takes longer, more changes are expected. Hyperpigmentation and hypertrophy of the RPE cells was more prominent in RetinoStat-treated eyes, where it took longer for the retina to reattach after subretinal delivery. The hyperpigmentation of the RPE did not interfere with the electroretinogram (ERG) and the effect slowly diminished over the course of the study in both species, being more prominent shortly after dosing, and decreasing slowly over time, where presumably excessive RPE cells were gradually removed from the retina by macrophagic activity.
The eye is an ideal target for gene therapy because it is anatomically separated from the rest of the body via the blood–retina barrier, which restricts any vector from leaving the target ocular structures. Most of the RetinoStat vector was detected within the rabbit ocular compartment, confirming the ability of the ocular anatomy to maintain the vector after subretinal dosing. Although the breach in Bruch's membrane in patients with wet AMD may potentially give RetinoStat access to the blood stream, it is expected that the VSV-G-pseudotyped vector would be inactivated by the human complement system and it is therefore unlikely that the RetinoStat vector would gain access to and affect tissues other than those in the eye (DePolo et al., 2000).
The average vector copy number per cell in the rabbit target retina/choroid was 0.9 on day 3 but dropped to 0.05 by day 183, a reduction of 18-fold. Because the method used to measure the presence of EIAV sequences does not distinguish between integrated and unintegrated vector sequences (such as 1-LTR and 2-LTR circles), this may represent the loss of unintegrated sequences. However, because this has not been shown empirically by LTR circle-specific PCR techniques, alternative explanations such as the turnover or removal of vector-transduced cells cannot be ruled out. However, there was only a 2-fold reduction in the level of endostatin and angiostatin in the vitreous over the course of the rabbit study. If the unintegrated species are lost over time and do not express as well as the integrated species, an 18-fold drop in copy number may translate to a 2-fold drop in protein levels, consistent with what we observed (Wanisch and Yáñez-Muñoz, 2009).
In the rabbit toxicology study the level of endostatin and angiostatin protein in the vitreous over the 6-month study period was in the range of 300–500 ng/ml for human endostatin and 300–400 ng/ml for human angiostatin. However, there is limited data reported in the literature to indicate potency levels of endostatin+angiostatin in the eye. To ensure that the histopathology was not compromised in the macaque GLP study vitreous samples were not obtained; however, EIAV-mediated gene expression in the macaque retina has previously been shown using reporter genes following a subretinal delivery route (Hamirally et al., 2009).
Because the retina is part of the ocular immune-privileged compartment, it was unknown whether subretinal administration of RetinoStat would result in the induction of an antibody response against components of RetinoStat. Although physical barriers and tolerance mechanisms are present within the eye to prevent and suppress inflammatory immune responses, they are not fully understood and may be affected by disease, age, injury (e.g., surgery), and presence of foreign material. In addition, the procedure used to administer RetinoStat may breach capillaries and therefore result in exposure of RetinoStat components to the systemic immune system. In these studies the immunological analysis in both rabbits and macaques indicated no antibody response to any vector components, including VSV-G or the transgenes, at early or late time points. Future studies investigating RetinoStat vector delivery to both eyes on separate occasions would be informative because ocular diseases are generally bilateral in nature and a second dose of RetinoStat in the contralateral eye may be required at some stage.
In conclusion, long-term safety studies performed in both rabbits and macaques demonstrated that a single subretinal injection of RetinoStat was safe and well tolerated with only a mild and transient cellular inflammatory response in the target eye. Ocular gene therapy after subretinal delivery is already being evaluated in individuals with Leber congenital amaurosis (LCA), which is a group of hereditary retinal dystrophies characterized by severe loss of retinal and visual function (Bainbridge et al., 2008; Hauswirth et al., 2008; Maguire et al., 2008). In these studies the RPE65 gene is delivered by an adeno-associated virus vector (AAV), and the safety of the subretinal intervention and results showing stability of the improvement in visual and retinal function support the use of gene therapy for chronic ocular diseases (Simonelli et al., 2010). These preclinical data with RetinoStat provided support for the initiation of the first-in-human clinical trial of RetinoStat in patients with the “wet” form of age-related macular degeneration that is currently underway.
Footnotes
Acknowledgments
The authors are grateful to the manufacturing and PAR teams at Oxford BioMedica for the production of RetinoStat IH39 vector. The authors thank Sanofi for supporting this work.
Author Disclosure Statement
Competing financial interests exist for Katie Binley, Michelle Kelleher, Jackie de Belin, Julie Loader, Georgina Ferrige, Marie Carlucci, Margaret Esapa, Daniel Chipchase, Diana Angell-Manning, Scott Ellis, Kyriacos Mitrophanous, James Miskin, and Stuart Naylor. No competing financial interests exist for Peter Widdowson, Vlad Bantseev, T. Michael Nork, and Paul Miller.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.