
Abstract
Stem cell mobilization to injured tissue contributes to neovascularization, resulting in regeneration after myocardial infarction (MI). We previously showed that direct cardiac injection of a recombinant lentivirus (LV) that engineers expression of membrane-bound stem cell factor (mSCF) improves outcomes immediately after MI. In this study, we evaluated the effect of neonatal LV/mSCF transduction on MI outcomes in aged mice. We constructed a recombinant LV harboring an α-myosin heavy chain promoter that drives mSCF expression and injected it into the temporal vein of neonatal mice. One year later, sustained expression of mSCF in the adult mouse hearts was detected by genomic and quantitative RT-PCR and immunohistochemistry. To evaluate the contribution of neonatal LV/mSCF delivery to recovery from MI, we induced an MI in adult LV/mSCF-transduced, LV only-transduced, and nontransduced control mice. Strikingly, LV/mSCF transduction reduced infarct scar size, enhanced angiogenesis, improved ventricular function, and significantly increased survival of the mice. Regional overexpression of CD11b, a marker of monocytes and proangiogenic cells, was observed on monocytes isolated from the infarcted hearts of LV/mSCF-transduced mice. Our data suggest a model of neonatal gene delivery that leads to sustained mSCF expression during adulthood to aid recovery from MI and prevent heart failure.
Introduction
Stem cell factor (SCF; also known as Kit ligand) is a cytokine that binds to the c-Kit receptor (CD117). Ligand binding leads to receptor dimerization and activation of multiple signaling pathways involved in cell survival, proliferation, and differentiation (Ashman, 1999; Jiang et al., 2000; Ronnstrand, 2004). SCF has two isoforms as a result of alternative splicing and proteolytic cleavage. The membrane form of SCF (mSCF) is missing a proteolytic cleavage site encoded by exon 6, which causes anchoring of the majority of protein to the membrane (Ashman, 1999). However, mSCF may also yield secreted SCF (sSCF) by proteolytic cleavage (Longley et al., 1997). Association of sSCF with CD117 results in transient activation of the receptor, whereas mSCF prevents internalization of the receptor and promotes sustained downstream activation (Miyazawa et al., 1995; Kurosawa et al., 1996). Expression of both SCF ligand and its receptor are altered during cardiac injury, which suggests that this axis is important in cardiac remodeling. Expression of SCF mRNA is present in the normal heart (Huang et al., 1992) but is downregulated post-MI (Woldbaek et al., 2002). Furthermore, we have previously reported involvement of the CD117 axis in the mechanism of cardiac rescue and remodeling post-MI (Ayach et al., 2006). These reports suggest specific roles for SCF in the context of ischemic heart disease.
Lentiviral (LV) vectors are efficient gene delivery vehicles that have broad tropisms and infect both dividing and nondividing cells. We have reported that LV-mediated delivery of mSCF by a novel direct injection strategy improved survival post-MI (Higuchi et al., 2009). We have also shown that neonatal injection of LVs harboring cardiac-specific promoters successfully engineers transgene expression in the heart (Lee et al., 2011).
In the present study, we sought to determine the effects of neonatal LV-mediated delivery of mSCF on mouse cardiac function and recovery from MI later in life. We found that cardiac function at 12 to 14 months was normal. After MI, cardiac outcomes were significantly improved in aged LV/mSCF-transduced animals compared with LV only-transduced and nontransduced controls. This opens the door to potential novel treatments employing an LV-mediated neonatal delivery strategy that may subsequently protect susceptible individuals from heart failure after MI.
Materials and Methods
LV construction
The HIV-1-based recombinant LV backbone pCCL was kindly provided by L. Naldini (Milan, Italy). Briefly, the LV/mSCF used in these studies was constructed (see Fig. 1a) using the myocardial-specific promoter α-myosin heavy chain (α-MHC) to drive Kit ligand-2 (mSCF) expression (Higuchi et al., 2009; Lee et al., 2011). The vector control LV carried the α-MHC promoter to drive enhanced green fluorescent protein (GFP) expression.
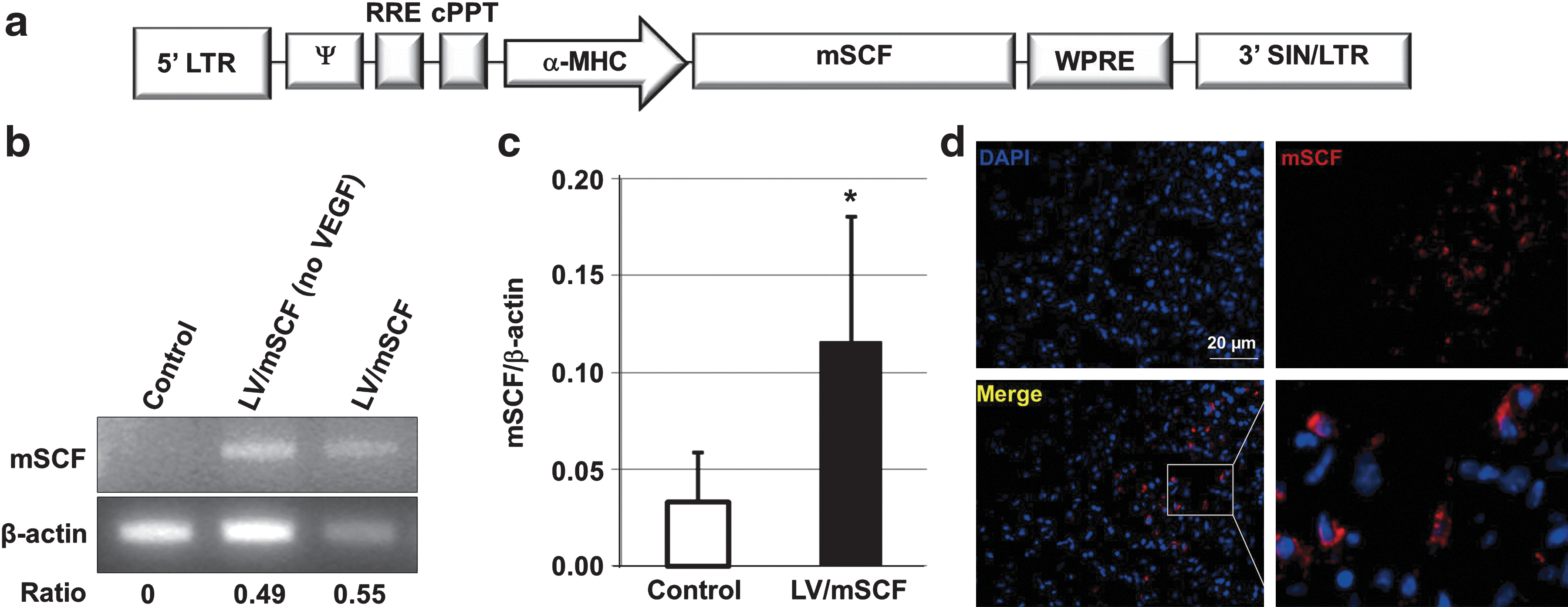
Expression of mSCF in aged mouse hearts 1 year after neonatal LV injection.
LV production and titration
Vesicular stomatitis virus glycoprotein (VSV-G)-pseudotyped LVs were generated by transient transfection of 293T cells with three plasmids (LV plasmid construct, packaging plasmid pCMVΔ8.91, and VSV-G envelope-coding plasmid pMD.G), using the polyethyleneimine transfection procedure as described previously (Lee et al., 2011). The concentrated virus was suspended in serum-free RPMI 1640 medium and stored at −80°C until use. Viral titers were determined with an HIV-1 p24 ELISA kit (PerkinElmer Canada, Vaudreuil-Dorion, QC, Canada).
Animals
All animal procedures were approved by the University Health Network Animal Care Committee and performed according to the Guide for the Care and Use of Laboratory Animals (NIH, revised 2010). C57BL/6 (wild-type) mice were purchased from Jackson Laboratory (Bar Harbor, ME). Neonatal C57BL/6 mice were transduced with LV/mSCF and housed until adults (n=25). Nontransduced (n=38) and LV only-transduced (n=18) C57BL/6 mice were age-matched to the LV/mSCF-transduced animals for all studies. The “aged” mice ranged in age from 12 to 14 months.
Injection of recombinant LV
The viral administration procedure has been described previously (Sato et al., 2009). We found that the administration of vascular endothelial growth factor (VEGF) before therapy enhances the efficiency of viral gene delivery to the heart (Sato et al., 2009); therefore, 1- to 3-day-old mice were injected with 1.7 ng of recombinant mouse VEGF (R&D Systems, Minneapolis, MN) via the superficial temporal vein 2 hr before LV injection. Then a maximum volume of 100 μl of virus (normalized to a titer of 300 ng of p24 per 100 μl for each construct) was injected into the superficial temporal vein of each recipient, using a 30-gauge needle.
Genomic DNA extraction and PCR analysis
Genomic DNA was extracted from 25 mg of heart tissue (n=8 per group), using a QIAamp DNA mini kit (Qiagen, Mississauga, ON, Canada); 100 ng of genomic DNA was subsequently amplified by PCR for mSCF sequences by use of specific primers 5′-ATGAAGAAGACACAAACTTGGATTATCACTTG-3′ and 5′-CAGGGGCCTTTGCGGCTTTC-3′. The cycling times used were 95°C for 2 min; followed by 30 cycles of 95°C for 15 sec, 55°C for 10 sec, and 70°C for 25 sec, using DNA polymerase from Thermococcus kodakaraensis (KOD polymerase, cat. no. 71086; Novagen, Mississauga, ON, Canada). β-Actin (primers 5′-TCCTGTGGCATCCACGAAACT-3′ and 5′-GAAGCATTTGCGGTGGACGA-3′) was amplified analogously as a control. PCR products were analyzed by electrophoresis on a 1.5% agarose gel and visualized by ethidium bromide staining.
Analysis of RNA expression
Total RNA was extracted from fresh tissue (n=8 per group) with an RNeasy kit (Qiagen), using a protocol recommended by the manufacturer. RNA was reverse transcribed with a ThermoScript RT-PCR kit (Invitrogen, Carlsbad, CA). mSCF and reporter gene β-actin mRNA expression levels were analyzed and calculated by quantitative reverse-transcription PCR (qRT-PCT) with SYBR green as a reporter, using the Corbett rotor gene RG-3000 system (Qiagen). Relative mSCF mRNA expression was determined by the ΔΔC T method (Livak and Schmittgen, 2001). Primer sets for mSCF and β-actin were designed with the help of Primer3Plus software (Untergasser et al., 2007), and specificity was confirmed by verification of the presence of a single peak of the expected melting temperature on a dissociation curve and by the presence of a single band of the PCR product after agarose gel electrophoresis. RNA primer sequences were as follows: mSCF forward, 5′-TCCGAAGAGGCCAGAAACTA-3′; reverse, 5′-TTTCTCGGGACCTAATGTTGA-3′; β-actin forward, 5′-GCTACAGCTTCACCACCACA-3′; reverse, 5′-TCTCCAGGGAGGAAGAGGAT-3′. Values are reported as mSCF mRNA expression relative to β-actin.
Immunohistochemical staining for mSCF expression
Cardiac mSCF expression in aged LV/mSCF-transduced mice was analyzed by immunohistochemical staining. Briefly, frozen heart tissue sections were fixed with methanol–acetone solution, stained with an anti-SCF antibody (SC-1494; Santa Cruz Biotechnology, Santa Cruz, CA), and then examined by fluorescence microscopy.
MI model
Mice were anesthetized with 2% isoflurane during mechanical ventilation (n=30 for nontransduced control, n=18 for LV only, and n=17 for LV/mSCF). Permanent ligation of the left anterior descending (LAD) coronary artery was used to generate an infarct in the hearts of adult mice as described previously (Sun et al., 2008). In brief, the left anterior thorax was opened after intubation, and the LAD coronary artery was exposed and permanently ligated with 7-0 SURGIPRO sutures (Covidien, Dublin, Ireland). Each surviving mouse received antibiotics (duplocillin LA, 150,000 IU/kg; Intervet Canada, Whitby, ON, Canada) and analgesic (buprenorphine, 0.01–0.05 mg/kg) subcutaneously after the surgery.
Evaluation of cardiac function
Mice were anesthetized with 2% isoflurane, and echocardiographic examinations (n=5 per group) were performed with an Acuson Sequoia C256 system (Siemens Medical Solutions USA, Malvern, PA) with a 13L8 transducer. Left ventricular dimensions (left ventricular end-diastolic diameter [LVEDd] and end-systolic diameter [LVEDs]) and left ventricular areas (left ventricular end-diastolic area [LVEAd] and end-systolic area [LVEAs]) were measured in M-mode at short-axis views of the left ventricle at mid-level. Fractional shortening (%) was calculated as follows: (LVEDd – LVEDs)/LVEDd×100. Fractional area contraction (%) was calculated as follows: (LVEAd – LVEAs)/LVEAd×100.
Cardiac function was also evaluated with a pressure–volume catheter 21 days after MI (n=5 per group). Mice were anesthetized with 2% isoflurane, and a calibrated Millar conductance pressure–volume catheter (Millar Instruments, Houston, TX) was inserted into the left ventricular cavity through the right carotid artery. After steady-state pressure and volume measurements, pressure–volume loops were recorded after inferior vena cava occlusion. Ejection fraction (%EF) was calculated as (LVEVd – LVEVs)/LVEVd×100. Parallel conductance was evaluated after the injection of a hypertonic saline solution (10% NaCl; BDH, Toronto, ON, Canada) into the jugular vein. The left ventricular pressure–volume relationship was analyzed as previously described (Sun et al., 2008).
Histological and immunofluorescence staining of hearts
For the histological study 3 days post-MI, mice were anesthetized with 2% isoflurane and then killed by continuous perfusion with 2% paraformaldehyde solution (Fisher Scientific, Ottawa, ON, Canada) through the ascending aorta. The hearts were fixed in the same solution for 24 hr and then transferred to 10% sucrose (BioShop Canada, Burlington, ON, Canada) for 1 hr, followed by 20% sucrose for 1 hr, and 30% sucrose for 24 hr. The hearts were embedded in O.C.T. compound and frozen for cryosectioning.
The histological study 21 days post-MI was performed after pressure–volume measurements. Hearts were arrested at diastole with 10% KCl (BioShop Canada) and perfusion-fixed at physiological pressure, using buffered 10% formalin (EMD Chemicals, Gibbstown, NJ). After fixation, 1-mm sections were embedded in paraffin wax, cut into 10-μm-thick sections, and stained with hematoxylin and eosin (H&E) and Masson's trichrome. Antibodies against CD11b (11-0112-85; eBioscience, San Diego, CA), CD31 (550274; BD Pharmingen, Mississauga, ON, Canada), and α-SMA (A-2574; Sigma, Oakville, ON, Canada) were used. Stained sections were viewed with a Nikon Ti-S Eclipse microscope. Images were captured with a Nikon Digital Sight DS-02 camera using NIS-Elements BR version 3.0 software. Blood vessels were counted in three sections per animal (n=4 per group). Four fields of 0.4 mm2 were analyzed per section by a blinded examiner.
Heart morphology studies
The hearts were fixed with 10% formaldehyde for 5 days and then sliced into 1-mm-thick slices. For scar size (n=5 per group), five consecutive slices were photographed, and the cross-sectional areas of the left ventricle free wall and the scar were measured by computed planimetry. For scar thickness (n=5 per group), the third slice was embedded in paraffin wax, cut into 10-μm-thick sections, and stained with Masson's trichrome. Scar thickness was calculated as the ratio (scar region vs. noninfarcted left wall) of the average of five randomly distributed measurements within the section, as described previously (Tang et al., 2010).
Flow cytometry
At the termination point of the study, mice were anesthetized, intubated, and ventilated with 2% isoflurane. Blood was collected from the right jugular vein, using a 0.5-ml insulin syringe. Blood was flushed from the heart by continuous saline perfusion through the aorta at physiological pressures until the liquid flowing from the right atrium was clear. Hearts were collected, minced, and digested with 0.1% collagenase (type II; Worthington, Lakewood, NJ) at 37°C for 30 min. Bone marrow was flushed from femurs and tibias. Red blood cells were depleted from blood samples, using lysis buffer (Tris and NH4Cl; BioShop Canada). Cells from tissues were passed through a 70-μm filter (Becton Dickinson, Franklin Lakes, NJ). Flow cytometric analyses (n=4 per group) used the following antibodies: Sca-1 (553336; BD Pharmingen), CD117 (12-1171-81; eBioscience), and CD11b (11-0112-85; eBioscience). The labeled cells were analyzed with an EPICS XLMCL flow cytometer with Expo32 ADC Xa software (Beckman Coulter Canada, Mississauga, ON, Canada).
Statistical analysis
All data are expressed as means±SD. Between-group comparisons were made by repeated measures t test for two groups. The echocardiography trend analysis data were evaluated with GraphPad Prism (GraphPad Software, San Diego, CA). Differences were specified by HSD (honestly significant difference) multiple range tests. Survival data were analyzed by Kaplan–Meier survival curves and log-rank statistics. Differences were deemed significant at p<0.05.
Results
Expression of mSCF in aged mice
To study the possible therapeutic role of mSCF in cardiac recovery after MI in aged mice, we constructed a novel recombinant LV harboring an α-MHC promoter that drives murine mSCF expression (LV/mSCF; Fig. 1a). One-day-old C57BL/6 mice were injected with LV/mSCF via the temporal vein, along with recombinant VEGF protein. We have previously found that the administration of VEGF before therapy enhances the efficiency of viral gene delivery to the heart (Sato et al., 2009). One year later, transgene expression in the hearts of the mice was detected by genomic PCR, qRT-PCR, and immunohistochemical staining. Importantly, we found that the heart tissue of the 1-year-old recipients still had substantial levels of mSCF expression as measured by DNA, RNA, and protein analyses (Fig. 1b–d). Quantitative RT-PCR demonstrated that LV/mSCF-transduced mice expressed significantly more mSCF than did the nontransduced controls (Fig. 1c). These results suggest that our LV delivery system and promoter of choice were successful in maintaining long-term therapeutic gene expression in the heart.
Neonatal LV/mSCF injection improves survival in aged mice post-MI
To evaluate the effect of mSCF on survival after MI, we induced MI in aged LV/mSCF-transduced, LV only-transduced, and nontransduced control mice. We observed 67% survival in nontransduced control mice, 72% survival in LV only-transduced mice, and 94% survival in LV/mSCF-transduced mice 3 weeks after MI (Fig. 2a). This implies that heart-targeted mSCF transduction can significantly decrease mortality post-MI, even 1 year after LV/mSCF administration. In addition, we measured the body and heart weights of the aged animals at the end of the study (21 days post-MI). Body weight did not differ among the groups (Fig. 2b); however, the heart-to-body weight ratio was significantly higher in nontransduced control mice and LV only-transduced mice compared with LV/mSCF-transduced mice (Fig. 2c). This compensatory hypertrophy in the infarcted hearts was significantly less in the LV/mSCF mice, indicating better cardiac outcomes in these mice post-MI versus the vector and nontransduced control animals.
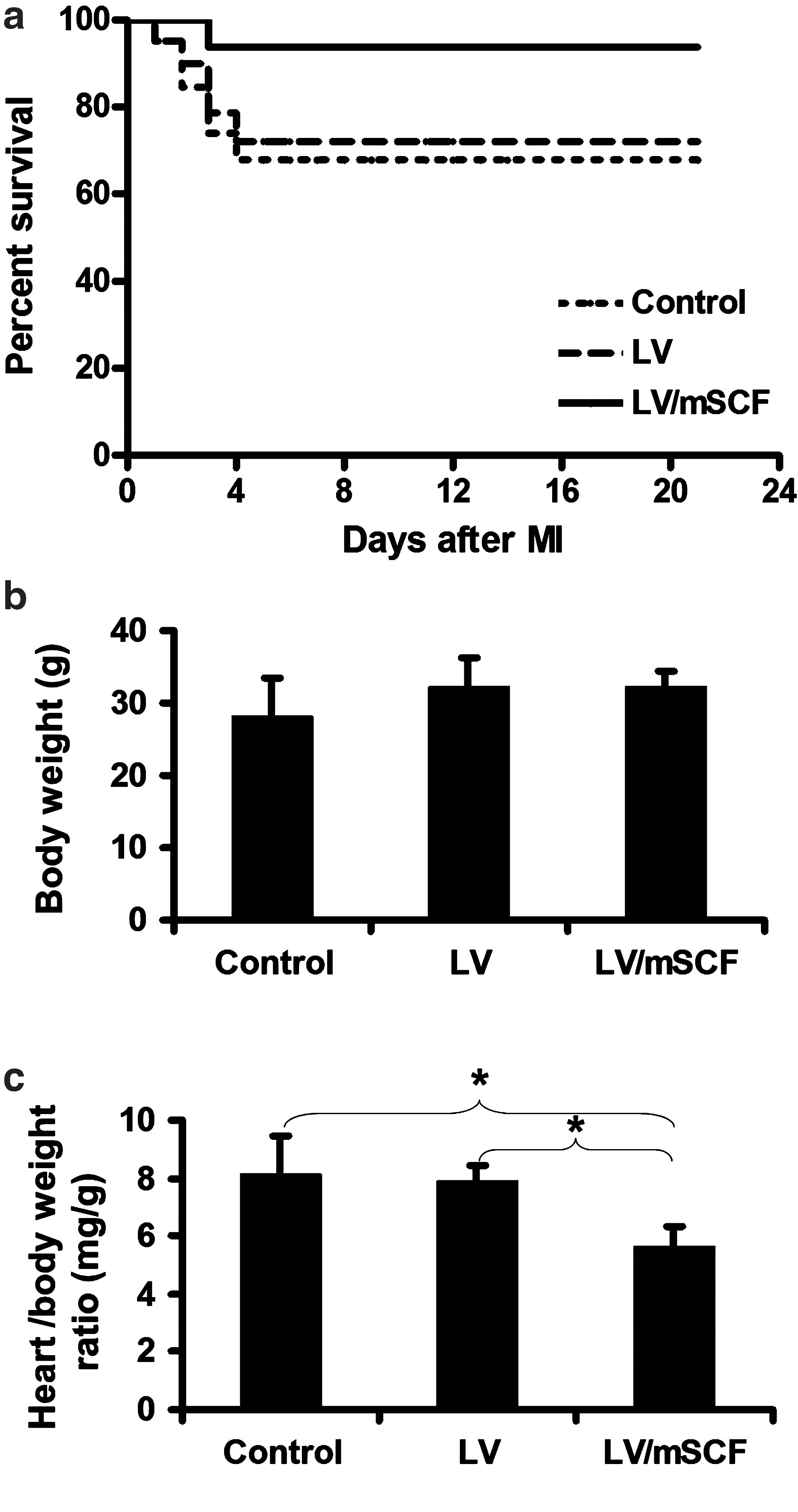
Neonatal LV/mSCF injection improved survival in aged mice post-myocardial infarction (MI).
Neonatal LV/mSCF injection improves heart function in aged mice post-MI
Cardiac function was evaluated by echocardiography before and for 3 weeks after MI. All mice exhibited normal heart function on day 0, as measured by fractional shortening and fractional area contraction, with a similar sharp decrease in function 3 days post-MI (Fig. 3a). However, LV/mSCF transduction prevented the progressive deterioration in ventricular function that was exhibited in the LV and nontransduced control mice from day 7 onward.
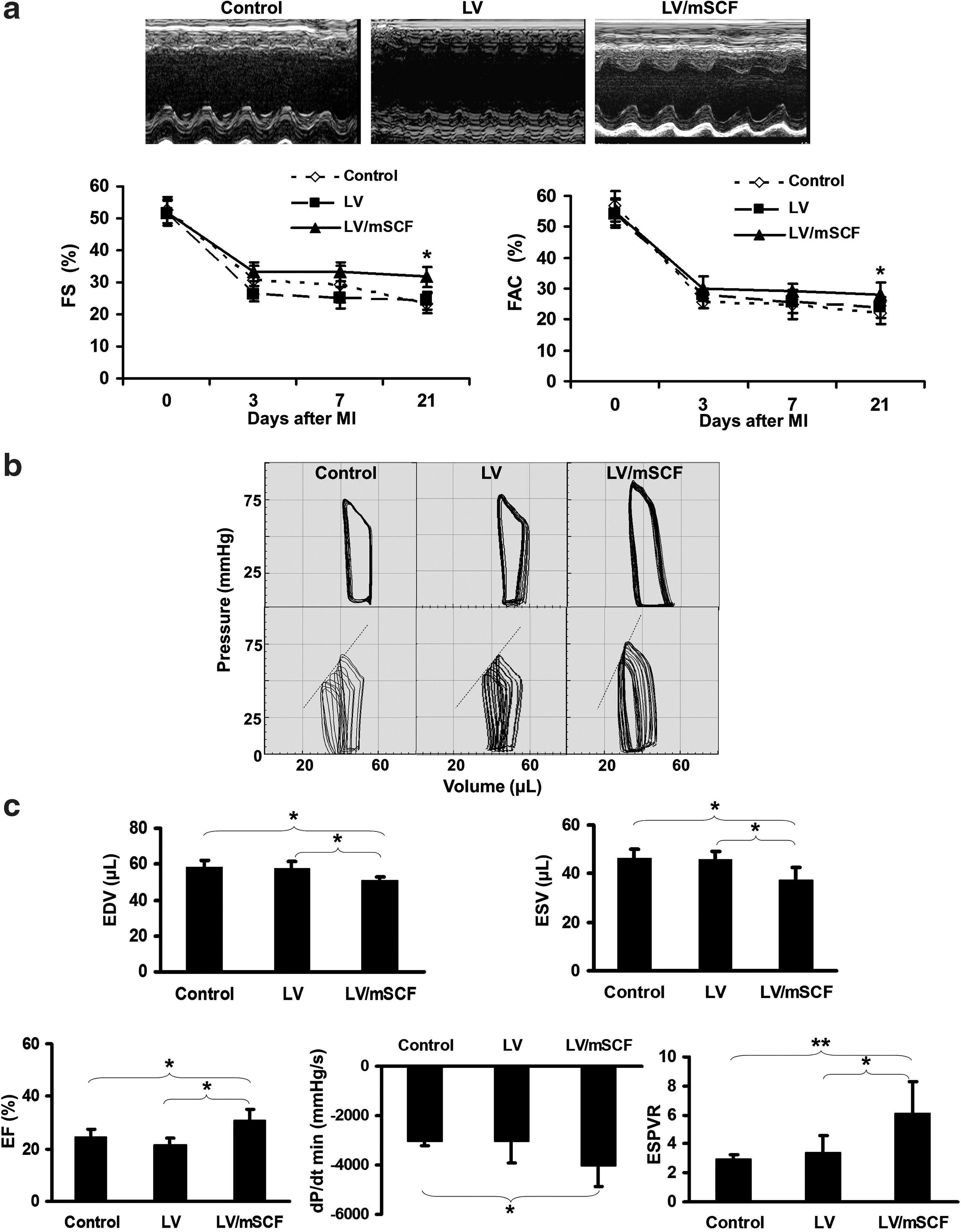
Neonatal LV/mSCF injection improved heart function in aged mice post-MI.
Cardiac function was further assessed by pressure–volume analysis 3 weeks post-MI (Fig. 3b). The end-systolic pressure–volume relationship (ESPVR), a load-independent hemodynamic parameter measured during transient inferior vena cava occlusion, is used to evaluate changes in systolic function. As an index of cardiac contractility that can be defined by the slope of the line obtained, a decline in the ESPVR indicates a decrease in contractility (Fig. 3b, bottom). Ventricular volumes, both end-diastolic and end-systolic, were significantly smaller in the LV/mSCF-transduced group compared with the LV only-transduced and nontransduced control groups (Fig. 3c). Consistent with the echocardiography results, the ejection fraction, an index of systolic function, was significantly greater in LV/mSCF-transduced mice than in the control mice (Fig. 3c). dP/dt min, an index of diastolic function, was also significantly improved in the LV/mSCF group compared with the nontransduced control group (Fig. 3c). The ESPVR in the LV/mSCF-transduced group was significantly greater than that in both control groups (Fig. 3c). These results further demonstrate that mSCF expression preserved heart function in aged mice post-MI.
Neonatal LV/mSCF injection reduces infarct scar size and prevents scar thinning in aged mice post-MI
Morphometric analysis was performed 21 days post-MI. In the LV/mSCF-transduced group, left ventricular infarct scar size was significantly smaller compared with controls (38.5±2.6 vs. 48.3±4.7% in nontransduced mice and 51.3±6.3% in LV only-transduced mice; Fig. 4a). Scar thickness was significantly greater in LV/mSCF-transduced mice (64.1±3.6%) compared with nontransduced (43.9±5.1%) and LV control (39.6±7.5%) groups (Fig. 4b). These results are consistent with the heart-to-body weight ratios (Fig. 2c), and together these data indicate that neonatal LV/mSCF transduction prevented adverse cardiac remodeling.
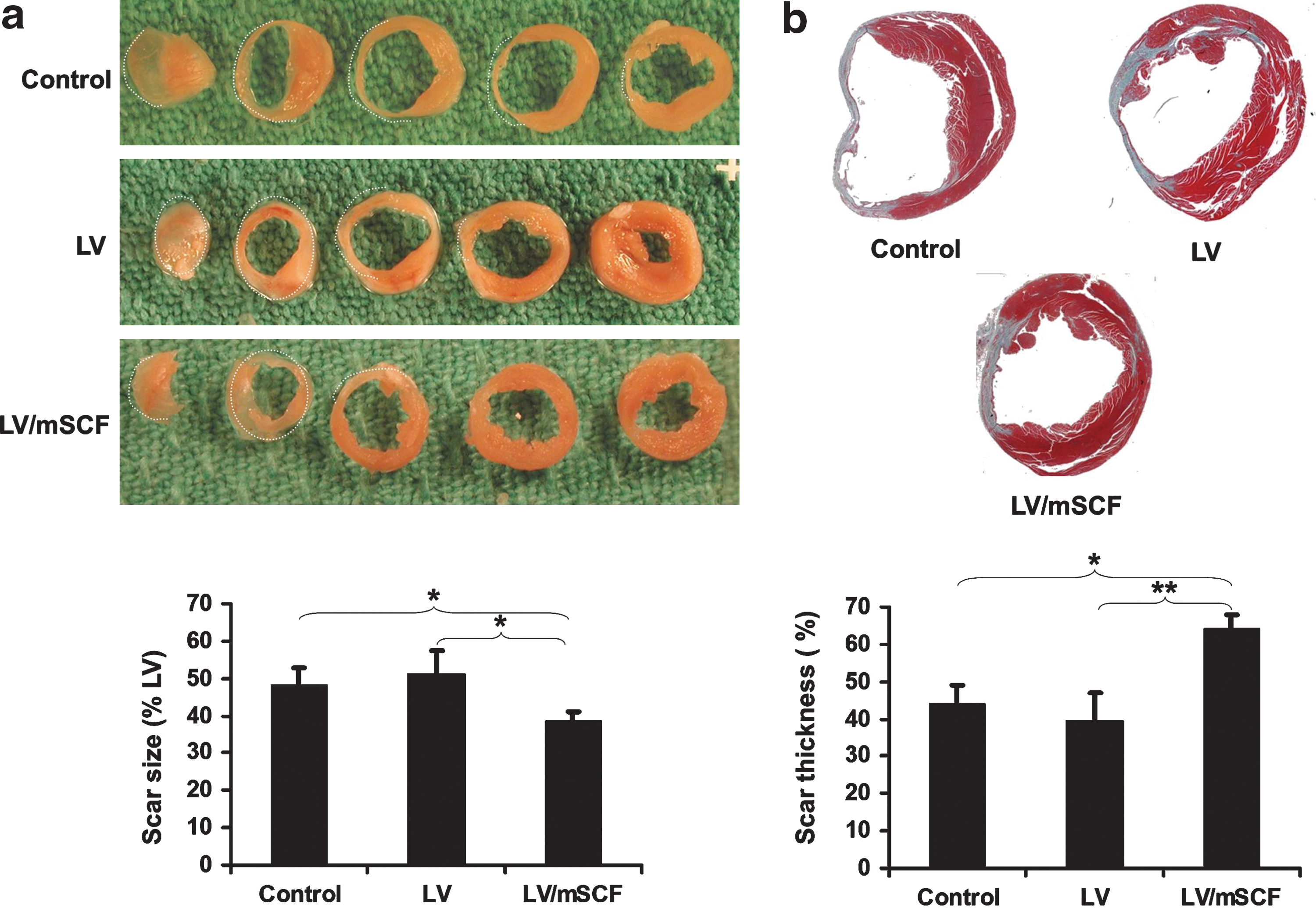
Neonatal LV/mSCF injection reduced infarct scar size and prevented scar thinning in aged mice post-MI.
Sustained mSCF expression recruits increased numbers of CD11b+ cells to the heart post-MI
To gain insight into this mSCF-based protection, heart, bone marrow (BM), and peripheral blood were obtained from aged mice before ligation and 3 days postligation. Stem cell and mononuclear cell markers, including CD117, Sca-1, and CD11b, were analyzed by flow cytometry. On day 3 post-MI, no differences were observed in the percentage of heart cells expressing CD117 and Sca-1 (stem cell markers) between LV/mSCF-transduced and nontransduced control mice (Fig. 5a). However, a dramatic change in CD11b (a monocyte marker and marker of proangiogenic cells; Sasajima et al., 2010) expression was observed in heart cells from before ligation to 3 days after ligation. The percentage of cells derived from LV/mSCF-transduced hearts that expressed CD11b increased from 2.4% before MI to 58.0% 3 days post-MI, which is significantly higher than the 24.7% of analogous cells from the nontransduced control mice (Fig. 5b). We also evaluated CD11b expression on cells from the BM and peripheral blood; no differences were observed between the groups 3 days post-MI.

Localized overexpression of mSCF recruited more CD11b+ cells to the heart post-MI.
CD11b+ cells promote neovascularization post-MI
To confirm the flow cytometric results described above, we performed immunofluorescence staining on heart tissue sections. Because CD11b is a marker for proangiogenic cells, we also stained for CD31 on day 3 post-MI to identify the presence of endothelial cells and investigate whether the accumulated CD11b+ cells possessed the capacity to regenerate vessels. We observed significantly more CD31+ vasculature in the ischemic region of LV/mSCF-transduced hearts than in nontransduced control hearts (Fig. 6a). Increased numbers of CD11b+ cells were again observed in the hearts of LV/mSCF-transduced mice compared with nontransduced control mice (Fig. 6b). As demonstrated by CD11b/CD31 costaining (Fig. 6b), the CD11b+ cells in the LV/mSCF-transduced hearts were localized primarily to the perivascular area, closely associated with the vasculature rather than directly differentiating into vascular endothelial cells.

CD11b+ cells promoted neovascularization and functional vasculature post-MI.
Because the CD31+ vasculature formed on day 3 post-MI may not represent functional vessels, we evaluated whether the vessels formed in LV/mSCF-transduced hearts were structurally mature. α-SMA expression delineates the smooth muscle layer in arterioles and arteries, which provide blood perfusion to tissues; therefore, staining for α-SMA was performed on heart sections obtained from animals 21 days post-MI. Vascular density was significantly greater in LV/mSCF-transduced mice than in nontransduced control mice (Fig. 6c).
Discussion
In this study, we evaluated the feasibility of neonatal LV/mSCF administration in protecting aged mice from cardiac dysfunction after MI. We found evidence of LV/mSCF persistence/expression in the mice 1 year after neonatal transduction. In a mouse MI model, LV/mSCF transduction reduced infarct scar size, enhanced angiogenesis, prevented ventricular dysfunction, and dramatically improved survival of the animals. In addition, we also provided evidence that sustained mSCF expression in the heart may contribute to CD11b+ monocyte recruitment for neovascularization of the infarcted myocardium.
SCF can bind to c-Kit (CD117), a receptor expressed on the surface of stem and progenitor cells, and exert a chemoattractant effect (Kim and Broxmeyer, 1998). Indeed, Vandervelde and colleagues (2005) demonstrated that upregulation of SCF after MI potentially results in the incorporation, proliferation, differentiation, and survival of BM stem cells in the infarcted heart, which is essential for cell-mediated repair post-MI. Others have hypothesized that the role of SCF may not be limited to c-Kit+ cells (Frangogiannis et al., 1998), suggesting that SCF may promote recruitment and homing of primitive BM-derived cells delivered to the infarct, with subsequent directed differentiation into cardiomyocytes and vascular cells (Orlic et al., 2003). Yamamoto and colleagues (2001) found that fibroblast-derived SCF upregulated monocyte chemoattractant protein-1 (MCP-1) expression in mast cells. MCP-1 is a potent chemotactic factor for monocytes. Along these lines, we observed more than a 2-fold increase in CD11b+ cell (monocytes/macrophages) accumulation in LV/mSCF-transduced mouse hearts 3 days post-MI. Monocytes/macrophages persist for days in the infarct zone (Nahrendorf et al., 2008); they accelerate vascularization in ischemic tissue (Herold et al., 2004) and promote infarct healing (Minatoguchi et al., 2004; Dewald et al., 2005), myocyte protection (Trial et al., 2004), and possibly regeneration (Eisenberg et al., 2003; Minatoguchi et al., 2004).
Neovascularization of the infarct border typically begins during the reparative phase of healing (Ren et al., 2002). It is clear that enhancement of angiogenesis on the infarct border post-MI improves heart function (Yau et al., 2001; Engel et al., 2006). In the present study, the number of CD31+ blood vessels in aged LV/mSCF-transduced animals was increased compared with nontransduced mice 3 days post-MI. Increased vessel density early after infarction may potentially increase blood supply to cardiomyocytes on the infarct border; however, it is important that early capillaries mature so that the blood supply is maintained. As the scar heals, the early vessels are either pruned or matured by gaining pericyte coverage. In the hearts of LV/mSCF-transduced mice, we showed that increased vessel density compared with nontransduced control mice was retained at 21 days post-MI, by which time many of the vessels had become coated with smooth muscle. Correspondingly, we showed that cardiac function in LV/mSCF-transduced mice was similar to that in control mice early after infarction, but by 21 days post-MI, it was significantly better in the LV/mSCF-transduced mice.
The infarcted myocardium forms the recipient environment for stem cell-mediated repair. Reports showed that SCF improved myocardial homing of c-Kit+ cells to the infarcted heart (Lutz et al., 2008; Fujii et al., 2009). In this study, we did not observe a difference in c-Kit (CD117) expression in the hearts of LV/mSCF-transduced and nontransduced groups post-MI. One possible explanation is that the c-Kit+ cell homing might be impaired in aged mice compared with young mice, as has been revealed in a rat MI model (Lehrke et al., 2006).
We also compared the expression of CD11b, a marker of monocytes and proangiogenic cells, on cells from the heart, BM, and peripheral blood of aged LV/mSCF-transduced and nontransduced mice and found a significant difference only in the heart. Because α-MHC is specifically expressed in the heart but not in other organs (Lee et al., 2011), this regional cardiac-specific SCF expression due to our α-MHC proviral promoter may be a key factor in monocyte infiltration into the heart post-MI, without affecting other organs. It has been shown that in the presence of SCF, CD11b upregulation induced by eotaxin, MCP-2, and MCP-1 was enhanced, and the sensitivity of basophils to chemoattractants increased, whereas SCF alone did not induce considerable changes in CD11b expression (Heinemann et al., 2005). A possible reason why we did not observe a difference in CD11b expression between LV/mSCF-transduced and nontransduced mouse hearts before ligation may be that transgene-produced mSCF did not recruit CD11b+ cells into the heart until certain environmental conditions were met, such as inflammation, ischemia, or ischemia–reperfusion.
In summary, this is the first study to demonstrate improved survival and cardiac function in aged mice after MI, using a unique gene therapy strategy—LV/mSCF transduction of neonates. Preparing the endogenous healing system of the heart by gene therapy, like preparing the immune system through immunization, protected against myocardial ischemia that occurred later in life. The benefit of this strategy is that there is no time delay between the occurrence of MI and the administration of treatment. Another potential clinical application of LV/mSCF gene therapy is to systemically deliver it to post-MI patients, with the potential for long-term expression and protection from downstream complications of MI. This study provides a novel concept for the development of potential treatments of MI and the prevention of heart failure.
Footnotes
Acknowledgments
This work was supported by a grant from the Canadian Institutes of Health Research (MOP102535 to R.K.L.). R.K.L. holds a Canada Research Chair in Cardiac Regeneration.
Author Disclosure Statement
No competing financial interests exist.