
Abstract
In this study, we target the hypoxia inducible factor-1 alpha (HIF-1-alpha) pathway by short hairpin RNA interference therapy targeting prolyl hydroxylase-2 (shPHD2). We use the minicircle (MC) vector technology as an alternative for conventional nonviral plasmid (PL) vectors in order to improve neovascularization after unilateral hindlimb ischemia in a murine model. Gene expression and transfection efficiency of MC and PL, both in vitro and in vivo, were assessed using bioluminescence imaging (BLI) and firefly luciferase (Luc) reporter gene. C57Bl6 mice underwent unilateral electrocoagulation of the femoral artery and gastrocnemic muscle injection with MC-shPHD2, PL-shPHD2, or phosphate-buffered saline (PBS) as control. Blood flow recovery was monitored using laser Doppler perfusion imaging, and collaterals were visualized by immunohistochemistry and angiography. MC-Luc showed a 4.6-fold higher in vitro BLI signal compared with PL-Luc. BLI signals in vivo were 4.3×105±3.3×105 (MC-Luc) versus 0.4×105±0.3×105 (PL-Luc) at day 28 (p=0.016). Compared with PL-shPHD2 or PBS, MC-shPHD2 significantly improved blood flow recovery, up to 50% from day 3 until day 14 after ischemia induction. MC-shPHD2 significantly increased collateral density and capillary density, as monitored by alpha-smooth muscle actin expression and CD31+ expression, respectively. Angiography data confirmed the histological findings. Significant downregulation of PHD2 mRNA levels by MC-shPHD2 was confirmed by quantitative polymerase chain reaction. Finally, Western blot analysis confirmed significantly higher levels of HIF-1-alpha protein by MC-shPHD2, compared with PL-shPHD2 and PBS. This study provides initial evidence of a new potential therapeutic approach for peripheral artery disease. The combination of HIF-1-alpha pathway targeting by shPHD2 with the robust nonviral MC plasmid improved postischemic neovascularization, making this approach a promising potential treatment option for critical limb ischemia.
Introduction
C
Therapeutic angiogenesis represents an alternative treatment modality for these patients. However, despite the numerous positive results in animal studies, clinical trials using a single angiogenic factor by either recombinant protein or gene-based formulations have resulted in either negative or conflicting outcomes (Jones and Annex, 2007; Attanasio and Snell, 2009; Sluimer and Daemen, 2009; Muona et al., 2012). To increase the success of gene-based therapeutic angiogenesis in the clinical situation, various gene transfer techniques have undergone intense investigation in recent years. Nonviral plasmid DNA is commonly used in clinical trials because of safety concerns with viral vectors, but these conventional plasmids also may provoke negative side effects that affect the gene expression. For instance, the bacterial backbone may lead to unwanted heterochromatin formation that changes eukaryotic gene expression or induces immune responses to CpG sequences (Jechlinger, 2006; Huang et al., 2009; Kay et al., 2010). To resolve these issues, an attractive gene transfer tool (nonviral minicircle [MC] plasmid) was designed. MCs are supercoiled DNA molecules that are smaller compared with conventional plasmids and lack a bacterial origin of replication or an antibiotic resistance gene, features that greatly improve the transgene expression in terms of both expressed levels of gene expression as well as in the persistence of the gene expression (Jechlinger et al., 2004; Mayrhofer et al., 2009).
As indicated above, the results of clinical trials using a single angiogenic (growth) factor were disappointing (Khurana et al., 2005; Gupta et al., 2009). This suggests that the expression of a single angiogenic factor such as vascular endothelial growth factor alone may not be sufficient to improve neovascularization (Cao et al., 2003). Newer therapeutic approaches designed to circumvent this problem utilize the upstream transcriptional factor, hypoxia inducible factor-1 alpha (HIF-1-alpha). Factors expressed by the HIF pathway include vascular endothelial growth factor, fibroblast growth factor, insulin-like growth factor, erythropoietin, and nitric oxide synthase, among others (Yla-Herttuala and Alitalo, 2003; Rey and Semenza, 2010). However, the HIF-1-alpha protein has a limited biological half-life as HIF-1-alpha is degraded in normoxic conditions by oxygen-dependent prolyl hydroxylase-2 (PHD2) (Lee et al., 2004). PHD2 mediates the interaction of HIF-1-alpha with von Hippel–Lindau protein ubiquitin ligase complex, which leads to proteasomal degradation of HIF-1-alpha (Ho et al., 2006). Inhibition of HIF-1-alpha degradation through short hairpin RNA (shRNA) knockdown of PHD2 has been shown to significantly improve neovascularization, which in turn improved cardiac function in a mouse model of myocardial infarction (Huang et al., 2008). Moreover, inhibition of PHD by shRNA provides a promising proangiogenic therapeutic approach, as in vivo electrotransfer of plasmids with either shPHD2 or shPHD3 has been shown to significantly improve postischemic neovascularization in a mouse model of PAD (Loinard et al., 2009).
In the present study, we combine the advantages of MC with the angiogenic effects of the HIF-1-alpha pathway, through shRNA knockdown of PHD2 in a murine hindlimb ischemia model. We demonstrate that the combination of these two strong concepts offers a promising new avenue for improving postischemic neovascularization in the future.
Materials and Methods
Construction of MC plasmids and MC DNA
The MC plasmids are constructed and produced as described earlier (Huang et al., 2008, 2009). Briefly, for the production of MC, ubiquitin promoter-driving firefly luciferase (Luc) is amplified with Luc forward and Luc reverse primers using pUbiquitin-Luc as a template. The mouse PHD2 gene is cloned, as described earlier, with TGTGAGGAACTTGAGATCT as the short hairpin scramble antisense sequence, and with the fragment No. 2 knockdown site inserted after H1 promoter in the vector pSuper (Huang et al., 2008). The MC DNA plasmid used here (a gift from Dr. Mark Kay, Stanford University) was produced as described previously (Huang et al., 2009; Jia et al., 2010).
Cell culture and transfection
Mouse C2C12 cells were cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (Huang et al., 2008). All cells were maintained in a 5% CO2 incubator. For transfection, cells were seeded with a density of 2×105 cells/well in 6-well flat-bottom microassay plates (Falcon Co.) 24 hr before transfection. At 70%–80% confluence, cells were transfected with 4 μg of plasmids carrying the Luc reporter gene (PL-Luc) or equimolar 2 μg of MC carrying the Luc reporter gene (MC-Luc), using Lipofectamine 2000 (Invitrogen) according to the manufacturer's protocol. C2C12 cells show rapid proliferation with a doubling time of approximately 19 hr in vitro (Pisani et al., 2004). In order to more closely resemble the in vivo conditions of slower proliferating cells, C2C12 cells were exposed to 9,000 rad 3 hr before transfection, resulting in an optimal proliferation pattern (Supplementary Fig. S1; Supplementary Data are available online at
Noninvasive bioluminescence imaging to assess the duration of reporter gene expression
To compare the duration of gene expression in vivo, 25 μg of PL-Luc or equimolar 12.5 μg of MC-Luc was injected into normal gastrocnemius muscles of C57Bl6 mice, with PL-Luc injected on the right side and MC-Luc injected on the left side. Because of the well-known divergent neovascularization capacities of various mice strains (Fukino et al., 2003), the duration of gene expression in C57Bl6 mice in the present study was closely examined. The same mouse strain was used in the hindlimb ischemia experiments as described in the next paragraph. Bioluminescence imaging (BLI) was performed with the Xenogen In Vivo Imaging System on days 1, 3, 5, and 7, and weekly thereafter until the endpoint on day 28, by an investigator blinded to study conditions (E.J.B.), as previously described (Wu et al., 2002; Swijnenburg et al., 2008). BLI signals were quantified in maximum photons per second per centimeter squared per steradian (p/s/cm2/sr). To compare the duration of gene expression in vitro, irradiated C2C12 cells were imaged using BLI at 3, 24, and 48 hr after transfection.
Experimental animals
Ten-week-old C57Bl6 mice (Jackson Laboratory) were used (10 mice per group). All animal experiments were performed after approval of the relevant authorities at both institutes (Stanford Animal Research Committee and the Leiden University Animal Research Committee).
Surgical procedure to induce hindlimb ischemia
C57Bl6 mice possess a well-known fast recovery after hindlimb ischemia induction (Hellingman et al., 2010). Mice injected with MC-shPHD2 were expected to have enhanced blood flow recovery as compared with controls. Double electrocoagulation of both the common femoral artery and the popliteal artery distally was performed because it could provide a larger therapeutic window (Hellingman et al., 2010). This enables a better analysis of the enhancing effects of MC-shPHD2 on arteriogenesis. Before surgery, mice were anesthetized with an intraperitoneal injection of a combination of midazolam (5 mg/kg; Roche), medetomidine (0.5 mg/kg; Orion), and fentanyl (0.05 mg/kg; Janssen). A skin incision was made unilaterally from the left inguinal region to the knee. For a double-coagulation model such as that used in this study, both the common femoral artery and the popliteal artery were dissected from the nerve and vein as described before (Hellingman et al., 2010). An electrocoagulation of the common femoral artery was performed and followed by an electrocoagulation of the popliteal artery. After surgery, the skin was closed with 6–0 Ethilon sutures.
Laser Doppler perfusion imaging
To compare the neovascularization effects with equimolar concentrations, 25 μg of PL-shPHD2 and 12.5 μg of MC-shPHD2 was injected into the left gastrocnemius muscles of C57Bl6 mice (n=10/group) 24 hr after the induction of hindlimb ischemia. Additional mice (n=10) were injected with phosphate buffered saline (PBS; 50 μl) as a control. Measurements of paw perfusion were performed of the mouse hindlimbs before, directly after, biweekly until day 14, and weekly until 4 weeks after the surgical procedure with laser Doppler perfusion imaging (Moor Instruments). Perfusion was expressed as a ratio of the left (ischemic) to right (nonischemic) paw, as previously described (Hellingman et al., 2012).
Histological analysis
Additional animals were sacrificed 10 days after ischemia induction followed by injection of MC-shPHD2, PL-shPHD2, or PBS, with their calf and adductor muscles removed and fixed with 4% formaldehyde and paraffin-embedded. Serial 5 μm cross sections were generated. Sections were re-hydrated, and endogenous peroxidase activity was blocked for 20 min in methanol containing 0.3% hydrogen peroxide. Capillaries and collaterals were visualized using antibodies recognizing CD31 on endothelial cells or α-smooth muscle actin (SMA) in smooth muscle cells, as previously described (Hellingman et al., 2012; van der Bogt et al., 2012). Quantification of labeled tissue sections was performed using ImageJ (9 sections per mouse were analyzed to obtain the mean per animal, and 5 animals per group were measured).
Quantitative polymerase chain reaction
For real-time reverse transcriptase–polymerase chain reaction (RT-PCR), we used inventoried assay on demand primers sets (Applied Biosystems). Oligonucleotides for PHD2 (Applied Biosystems; EGLN1, Mm00459770_m1) were used to analyze PHD2 mRNA levels, and mouse HPRT (Applied Biosystems; HPRT, Mm00446969_m1) was used to normalize sample amplification. Polymerase chain reaction was performed on a 7500 Fast Real Time PCR System (Applied Biosystems). mRNA expression levels of PHD2 were corrected for expression of HPRT and displayed as relative expression values (ΔCt) and analyzed as previously described (van der Laan et al., 2012). RNA extraction was performed as described in the Supplementary Methods section.
Protein extraction and Western blot analysis
Protein from gastrocnemius muscle (n=14) was extracted and analyzed as previously described (Huang et al., 2008; Velotta et al., 2011). Cellular lysed homogenates in radioimmunoprecipitation assay buffer were isolated for protein concentration determination with a bicinchoninic acid assay (Thermo Scientific). Equal amounts of protein were loaded per well (20 μg of protein per well). Samples were extracted with a sample buffer (Biorad) containing 50 mM dithiothreitol as a reducing agent for 5 min at 95°C, resolved by polyacrylamide gel electrophoresis, and transferred to polyvinylidene fluoride membranes. The membranes were then blocked with 5% milk/Tris-buffered saline-Tween (TBST) for 1 hr at room temperature, incubated with the appropriate primary antibody at 4°C overnight, and washed with TBST. Primary antibodies used were HIF-1-alpha (1:200, NB100-479; Novus) and actin as control (1:1,000, SC 1615; Santa Cruz Biotech). The appropriate horseradish peroxidase-conjugated secondary antibody, diluted in 5% milk/TBST, was applied for 1 hr at room temperature. After washing with TBST, immunoblots were visualized and quantified by the Super Signal West Dura Extended Duration Substrate (Perbio Science), LabWorks 4.6 software, and a luminescent image workstation, as previously described (Lindeman et al., 2008; Mowat et al., 2010). Hif-1-alpha expression levels were corrected for expression of actin and displayed as relative expression values.
Statistical analysis
Results are expressed as mean±SEM. Comparisons between means were performed by an independent T-test. For repeated measures, one-way repeated-measures ANOVA with post-hoc Bonferroni–Holm's correction was used. p-values<0.05 were considered statistically significant. All calculations were performed in SPSS 19.0.
Results
Evaluation of MC vectors versus regular plasmid vectors in vitro
To assess the transfection efficiency, MC-Luc and PL-Luc were used in equimolar amounts to transfect mouse C2C12 cells. Luc was evaluated by BLI. MC-Luc showed a higher Luc expression compared with PL-Luc, with a 4.6-fold higher BLI signal at 24 hr and a 4.4-fold higher BLI signal at 48 hr in irradiated C2C12 cells (Fig. 1). As a control, in nonirradiated cells MC-Luc showed 2.5-fold higher Luc expression compared with PL-Luc at 24 hr after transfection (Supplementary Fig. S2).

In vitro BLI of irradiated C2C12 cells after transfection with MC-Luc or PL-Luc.
Comparison of MC versus regular plasmids in vivo
Equimolar amounts of MC-Luc and PL-Luc were injected into the left or right mouse gastrocnemius muscles, respectively, to determine expression levels in C57Bl6 mice in vivo. Gene expression was followed by BLI from day 0 until day 28. MC-Luc injection showed a significantly higher Luc activity compared with PL-Luc from day 3 until the end point at day 28, with BLI of 6.9×103±1.7×103 versus 3.6×103±0.46×103 p/s/cm2/sr at day 1 (p=ns); 3.2×105±2.4×105 versus 0.4×105±0.3×105 p/s/cm2/sr at day 7 (p=0.002); 3.5×105±2.8×105 versus 0.5×105±0.3×105 p/s/cm2/sr at day 21 (p=0.004); and 4.3×105±3.3×105 versus 0.4×105±0.3×105 p/s/cm2/sr at day 28 (p=0.016) (Fig. 2).
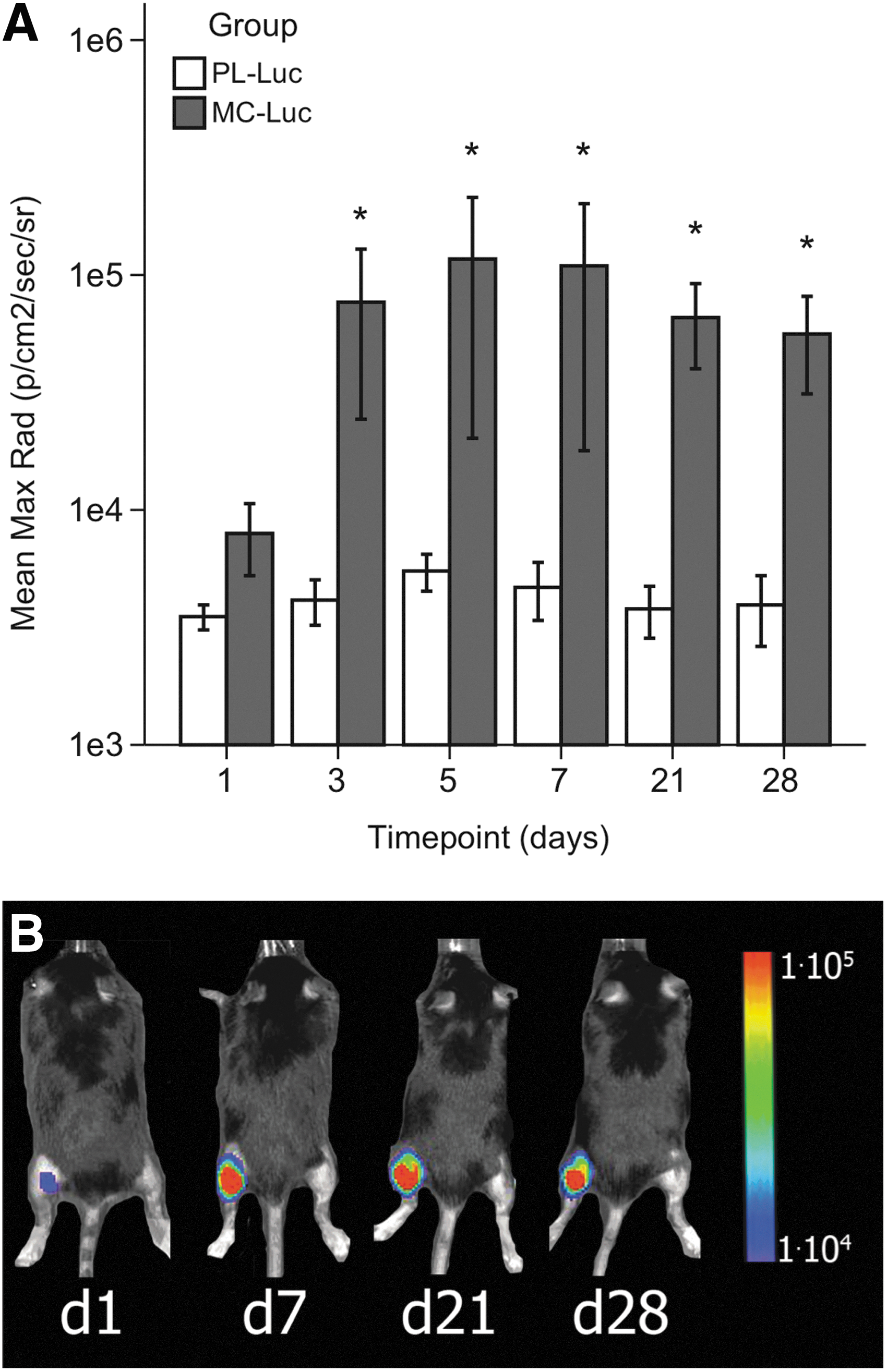
In vivo BLI of the transfection efficiency of MC-Luc compared with PL-Luc in C57Bl6 mice.
Injection of MC encoding shPHD2 improves postischemic blood flow recovery
To examine whether MC-shPHD2 could improve postischemic neovascularization in vivo as compared with PL-shPHD2 or PBS, hindlimb ischemia was performed in C57BL6 mice followed by injection of MC-shPHD2, PL-shPHD2, or PBS, respectively. After double electrocoagulation of both the common femoral artery and the popliteal artery, blood flow decreased to less than 5% in all mice. Mice injected with MC-shPHD2 showed significantly improved blood flow recovery, up to 50% from day 3 until day 14 after ischemia induction as compared with mice injected with PL-shPHD2 or PBS (Fig. 3). Injection of PL-shPHD2 did not improve blood flow recovery significantly as compared with PBS injection.

Paw perfusion as measured by LDPI.
Increase in collateral and capillary density in postischemic skeletal muscle after MC-shPHD2 injections
To study collateral artery formation at the tissue level, ischemic skeletal muscles were harvested 10 days after hindlimb ischemia induction, and immunohistochemical staining was performed. Mice injected with MC-shPHD2 showed a significant increase in collateral density, as measured by SMA expression, in the postischemic adductor muscle as compared with mice injected with PL-shPHD2 or PBS (3.5±0.5 vs. 1.9±0.2 and 1.0±0.1 collaterals per mm2) (Fig. 4A). Furthermore, collaterals were significantly larger in mice injected with MC-shPHD2 than in mice injected with PL-shPHD2 or PBS (850.5±397.9 μm2 vs. 258.8±91.7 μm2 and 196.2±33.6 μm2) (Fig. 4B). In addition, injection of MC-shPHD2 significantly increased the CD31+ capillary density of the postischemic calf muscles of mice as compared with injection of PL-shPHD2 or PBS (40.3±4.9 vs. 18.1±5.2 and 11.8±3.4 capillaries per mm2) (Fig. 4D). To illustrate collateral growth in the postischemic hindlimb, angiographs were made 10 days after hindlimb ischemia induction (see Supplementary Methods section). Collaterals were observed in the postischemic adductor muscles of all mice, indicating that C57Bl6 mice do have the capacity to form collaterals after hindlimb ischemia. Mice injected with MC-shPHD2 showed more typical corkscrew-like collateral arteries in the ischemic adductor muscle area as compared with mice injected with PL-shPHD2 or PBS. These angiographic results confirm the results of the SMA staining (Fig. 5).
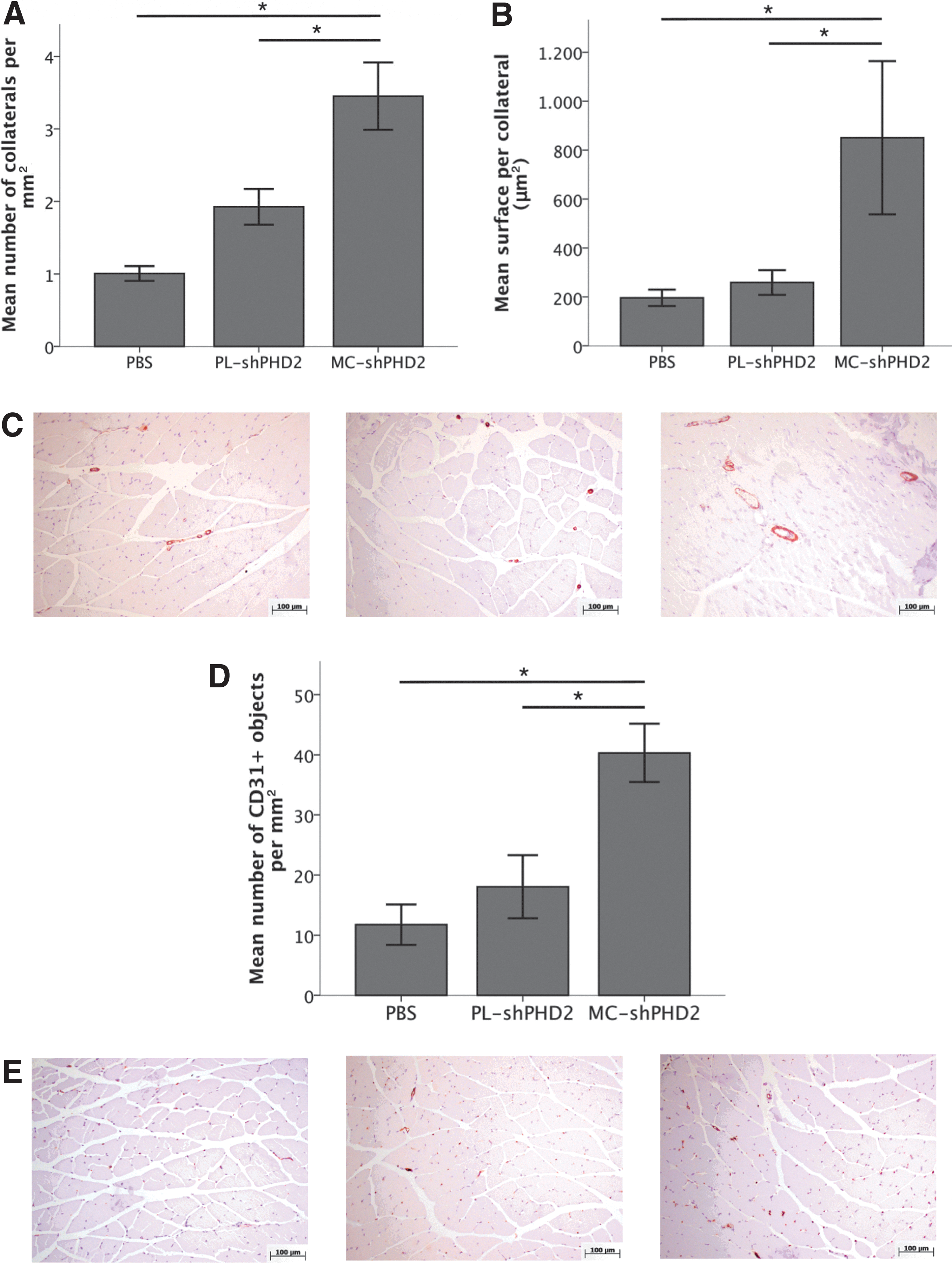
Immunohistochemical analyses of the adductor (SMA) and calf (CD31+) muscles harvested 10 days after surgery.
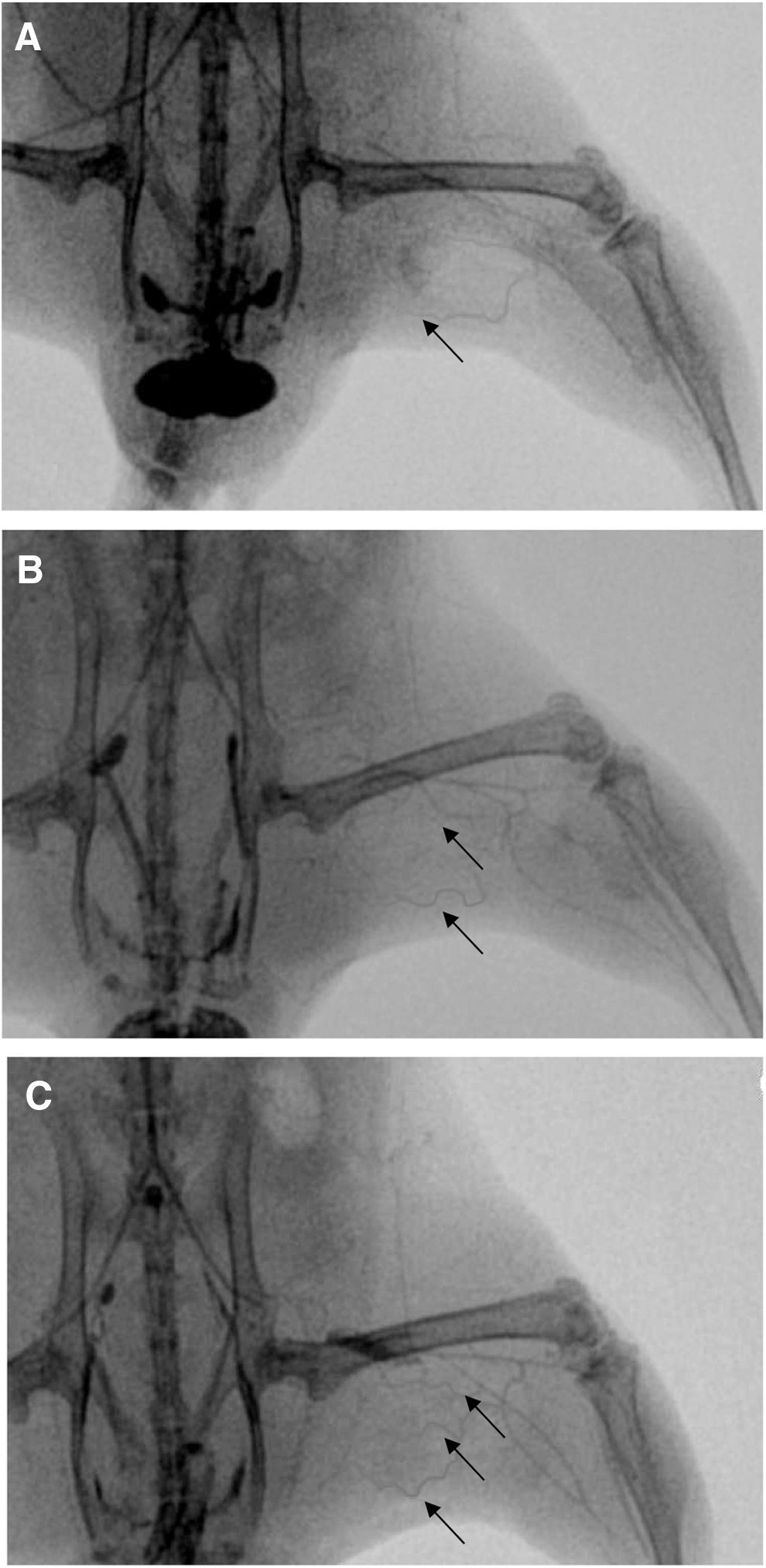
Representative example of angiographies showing collateral formation after
Quantitative PCR analysis of PHD2 mRNA expression
Additional C57BL6 mice were subjected to hindlimb ischemia followed by injection of MC-shPHD2, PL-shPHD2, or PBS, to assess PHD2 mRNA and HIF-1-alpha protein expression at 7 days after surgery. MC-shPHD2 injection significantly decreased the mean PHD2 mRNA expression in gastrocnemius muscles, with ΔCt levels of 0.28±0.022, compared with PL-shPHD2 and PBS (0.35±0.016 and 0.36±0.026, respectively; p<0.05) (Fig. 6).
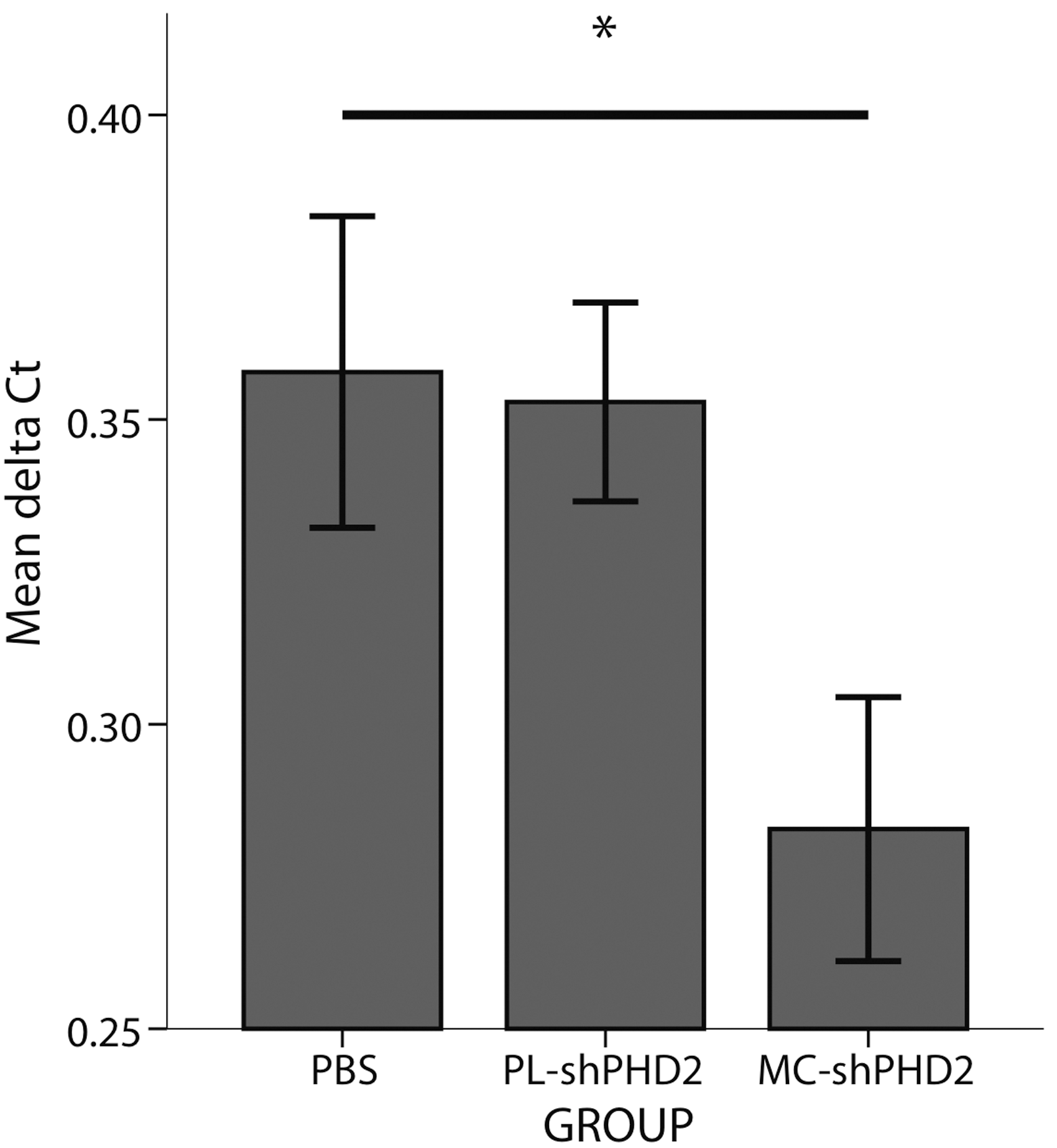
Graphical representation of the mean relative PHD2 mRNA levels (mean ΔCt) in the gastrocnemius muscles of mice 7 days after surgery and treated with MC-shPHD2, PL-shPHD2, or PBS, respectively (*p<0.05).
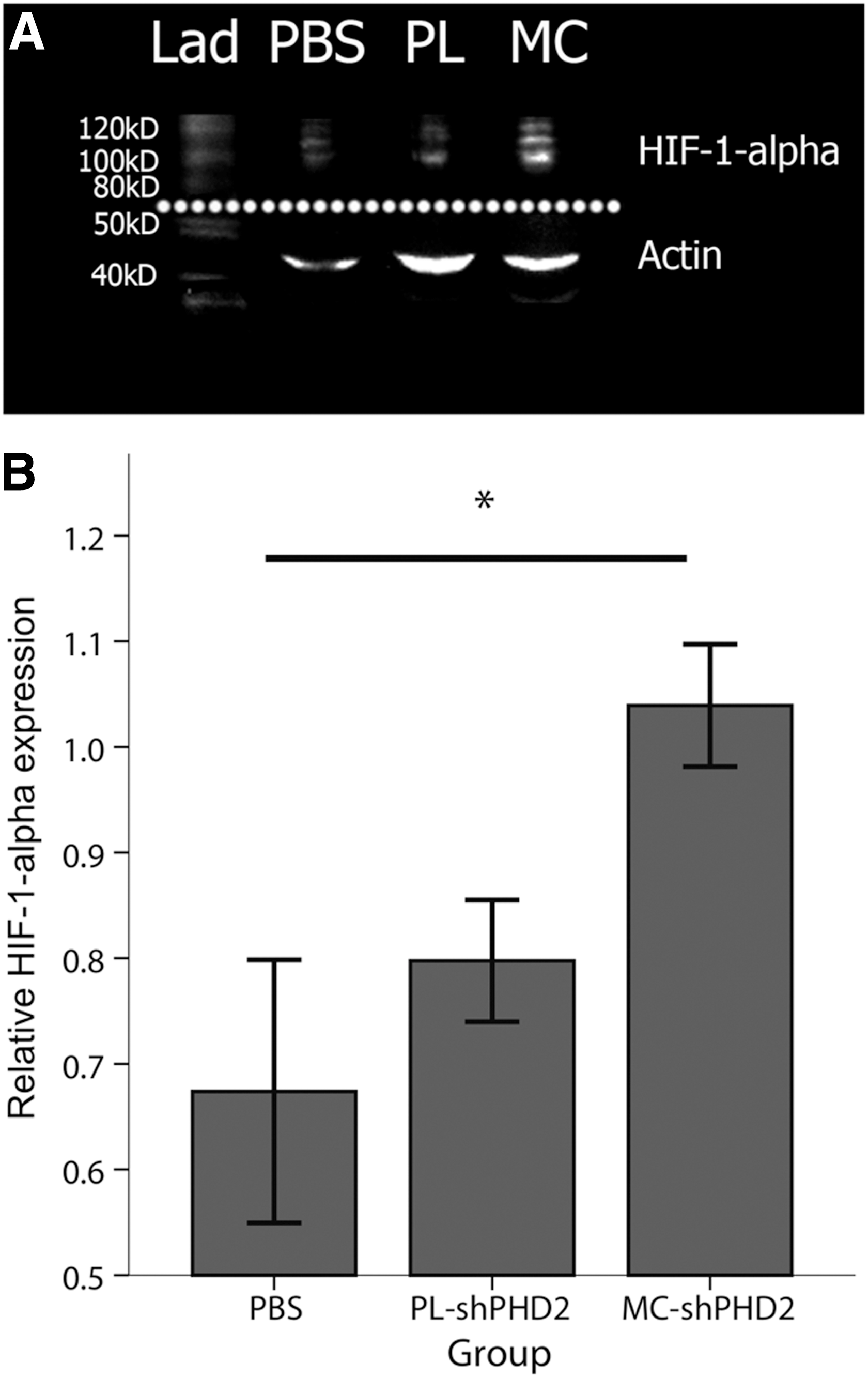
Western blot analysis of HIF-1-alpha protein expression
Western blot analysis of HIF-1-alpha protein expression in the gastrocnemius muscles at day 7 after surgery showed a significantly increased mean relative HIF-1-alpha protein level in the MC-shPHD2-treated group of 1.0±0.058, compared with PL-shPHD2 treatment (0.80±0.058) and PBS treatment (0.67±0.12) (p<0.05 for both) (Fig. 7).
Discussion
To our knowledge, this is the first study that combines the advantages of stimulating the angiogenic potential of the HIF-1-alpha pathway by shRNA interference of PHD2 with the nonviral MC vector technology. Our results indicate that MC-shPHD2 improves postischemic neovascularization in a hindlimb ischemia mouse model, making this approach an attractive potential treatment option for CLI.
Functional revascularization capacity of MC-shPHD2 was studied by repeated blood flow perfusion measurements after hindlimb ischemia induction in mice treated with MC-shPHD2, PL-shPHD2, or PBS. Mice treated with MC-shPHD2 showed significantly improved blood flow recovery up to 50% as compared with treatment with PL-shPHD2 or PBS. A significant increase in collateral and capillary density in the postischemic skeletal muscles of mice treated with MC-shPHD2 confirmed its beneficial effects on neovascularization. These results demonstrate the potential of MC-shPHD2 as a valuable target for therapeutic neovascularization.
Therapeutic neovascularization is a promising strategy for treatment of ischemic vascular disease that uses angiogenic growth factors or genes encoding these proteins to stimulate neovascularization. Single recombinant growth factors such as fibroblast growth factor (Baffour et al., 1992) and vascular endothelial growth factor (Takeshita et al., 1994) have been studied in preclinical studies, but their use seemed to be restricted by the limited half-life. For instance, phase I clinical trials achieved promising results using plasmid DNA encoding human growth factors in patients with PAD (Isner et al., 1996; Baumgartner et al., 2009; Gupta et al., 2009), but phase III clinical trials have failed to demonstrate unambiguous success that a single growth factor could benefit patients with CLI (Belch et al., 2011). Newer approaches based on upregulation of an upstream transcriptional factor as HIF-1-alpha may provide better alternatives.
Additionally, the inconsistent benefits of gene therapy using a single growth factor might also be attributable to the lack of ideal vectors and limited transfection efficiencies. To assess transfection efficiency in vitro, MC-Luc and PL-Luc were used in equimolar amounts to transfect irradiated mouse C2C12 cells. To correct for the rapid division of nonirradiated cells, we used irradiated cells that more closely resemble in vivo situation of slower proliferating cells. The present study reported up to 4.6-fold higher gene expression, which was even higher than the transfection efficiency shown in our control experiment with nonirradiated cells. Transfection efficiency in vivo was determined by the injection of MC-Luc and PL-Luc in the gastrocnemius muscles of C57Bl6 mice. Up to a 10-fold higher gene expression of MC-Luc during 28 days as compared with PL-Luc in the mouse hindlimb was reported in this study. This was in line with a recent report that compared gene expression of MC-Luc with PL-Luc in gastrocnemius muscles of FVB/N mice (Huang et al., 2009). These results confirm the same beneficial transfection efficiency of MCs in two different mice strains, which were found to have different neovascularization reactions on hindlimb ischemia (Chalothorn and Faber, 2010b). The mechanism of enhanced transgene expression of MCs is unclear, but may result from the elimination of the unnecessary plasmid sequences, in particular CPG islands, which can lead to transcriptional gene silencing in vivo. In addition, the smaller size of MCs may confer better extracellular and intracellular bioavailability and therefore improve gene delivery properties (Kay et al., 2010).
Previous studies on therapeutic angiogenesis have demonstrated regression of neovessels as time progressed because of a drop in expression after gene transfer (Dor et al., 2002; Tang et al., 2004; Gounis et al., 2005; Olea et al., 2009). By using MC as a vector, we and others have shown that gene expression levels are high up to 4 weeks after transfection (Huang et al., 2009). Additionally, previous studies have shown that the therapeutic window ranges from 1 to 3 weeks, with complete blood flow recovery in untreated control animals after these time points (Hellingman et al., 2010). Therefore, we used a single administration to analyze the effect on neovascularization. No significant neovessel regression was seen in our study. MC-shPHD2 treatment resulted in 100% blood flow recovery 3 weeks after ischemia induction. A nonsignificant decrease in blood flow was seen at 28 days in this group. This might be explained by the relative decrease in HIF-1-alpha levels in the normoxic paws. This is in concordance with the previously mentioned studies in which gene expression tends to vanish in the absence of an angiogenic stimulus. However, we believe that restoring the blood flow to the downstream tissue, and thus resolving the ischemia in the affected areas, is also an important factor that may explain the small decline in perfusion. After formation of sufficient new collaterals and consequently repair of the blood flow to the ischemic tissue, part of the newly formed collaterals will become redundant and will disappear, a process called pruning, whereas other collaterals will fully mature (Chalothorn and Faber, 2010a; LeBlanc et al., 2012). For future larger animal studies that require a longer therapeutic window, repeated transfection using MC vectors is feasible with robust transgene activities, with lower host cellular and humoral immune responses as seen with repeated injections of viral vectors (Huang et al., 2009).
In this study, we selected PHD2 as the target gene by downregulation of the mouse PHD2 gene by MC-mediated shRNA interference, which leads to activation of downstream angiogenic genes and proteins. In line with our results, a recent report showed that downregulation of PHD2 by shRNA enhanced neoangiogenesis in a mouse model of myocardial infarction (Huang et al., 2008). A different study evaluated the effects of shPHD1, shPHD2, and shPHD3 on neovascularization in a mouse hindlimb ischemia model (Loinard et al., 2009). Electroporation of plasmid vectors was used and silencing of PHDs triggered postischemic neovascularization, with shPHD2 and shPHD3 having the most robust effects. In the current study, we chose PHD2 as a target because it is the most abundant isoform (Wong et al., 2013). Both in vivo reports have provided a better understanding of PHD2 downregulation with shRNA in human subjects. Importantly, recent evidence suggests that downregulation of PHD2 promotes tumor growth (Bordoli et al., 2010). In this study, Bordoli et al. showed that PHD2 functions as a tumor suppressor in xenografted tumors derived from breast carcinoma. In this respect, as PHD inhibitors have been developed for the treatment of PAD, there are concerns over possible side effects of these inhibitors in tumor progression. More encouragingly, others have shown that genetic inactivation of PHD2 increased the delivery of chemotherapeutics to the tumor and thereby their antitumor and antimetastatic effect (Leite de Oliveira et al., 2012). This study also showed that PHD2 haplodeficiency prevented oxidative damage, organ failure, and tissue loss. In addition, a randomized, double-blind, placebo-controlled study using adenoviral HIF-1-alpha showed no evidence of promotion of tumor growth (Rajagopalan et al., 2007).
Finally, a recent randomized study studied the effect of adenoviral administration of HIF-1-alpha in a group of 289 patients with claudication (Creager et al., 2011). No significant differences in claudication onset time, ankle-brachial index, or quality-of-life measurements between the placebo and HIF-1-alpha groups were found. Possible explanations for the negative results include the limited biological activity of HIF-1-alpha in patients with claudication and the possible limited efficacy of gene transfer with an adenovirus vector, caused by immunological responses. Hence, shPHD2 treatment in combination with the MC vector might circumvent these issues. Although PAD is a chronic progressive disease, triggering the collateral formation and neovascularization might be very beneficial in restoring the blood flow to the ischemic limbs (Helisch and Schaper, 2003; Makino et al., 2012). MC-shPHD2 would therefore be an interesting alternative treatment strategy for therapeutic angiogenesis in the future.
In conclusion, the present study provides encouraging initial evidence for a new potential therapeutic approach for triggering the collateral formation and neovascularization in PAD. Using a nonviral MC vector carrying shPHD2, HIF-1-alpha is upregulated through PHD2 knockdown, resulting in an improved postischemic neovascularization in a hindlimb ischemia mouse model.
Footnotes
Acknowledgments
This work was supported in part by grants from the NIH R01 HL095571, NIH R01 HL093172, Burroughs Wellcome Foundation (J.C.W.), AHA Beginning Grant in Aid (M.H.), and The Netherlands Organization for Health Research and Development (M.A.L.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.