
Abstract
Mucopolysaccharidosis type II (MPS II) is a neuropathic lysosomal storage disorder caused by a deficiency of iduronate-2-sulfatase (IDS), which leads to the accumulation of glycosaminoglycans (GAGs). We demonstrated that biochemical alterations in the brains of MPS II mice are not corrected by bone marrow transplantation (BMT) or enzyme replacement therapy, although BMT has been shown to be effective for other neurodegenerative MPSs, such as Hurler syndrome. In this study, we demonstrated that lentiviral isogeneic hematopoietic stem cell (HSC) gene therapy corrected neuronal manifestations by ameliorating lysosomal storage and autophagic dysfunction in the brains of MPS II mice. IDS-transduced HSCs increased enzyme activity both in various visceral organs and the CNS. Decreased levels of GAGs were observed in many organs, including cerebra, after transplantation of IDS-transduced HSCs. In addition, lentiviral HSC gene therapy normalized the secondary accumulation of autophagic substrates, such as p62 and ubiquitin–protein conjugates, in cerebra. Furthermore, in contrast to naive MPS II mice, there was no deterioration of neuronal function observed in transplant recipients. These results indicated that lentiviral HSC gene therapy is a promising approach for the treatment of CNS lesions in MPS II.
Introduction
M
Enzyme replacement therapy (ERT) and hematopoietic stem cell transplantation (HSCT) are currently available to treat patients with MPS II. ERT has been shown to improve some clinical symptoms, such as hepatosplenomegaly and walking capacity 1 –3 but not CNS lesions, because enzymes do not cross the blood–brain barrier (BBB). 4 In addition, repeated administration of ERT is costly and puts further stress on patients with MPS II. HSCT was the only definitive treatment for MPS II before the development of ERT, and has been shown to correct visceral and CNS manifestations of several lysosomal storage diseases including MPS I, globoid-cell leukodystrophy, metachromatic leukodystrophy, and Gaucher disease. 5 Because engrafted donor cells differentiate into both hematopoietic cells and microglial cells that secrete a therapeutic enzyme allowing for cross-correction of neuronal cells in recipients, 6 HSCT seems to be an effective treatment of neuropathic lysosomal storage disorders. However, early studies reported that there were no beneficial effects of HSCT in CNS disease in patients with MPS II. 7 –9 In addition, our previous study demonstrated that bone marrow transplantation (BMT) does not correct biochemical alterations in the brains of MPS II mice, such as the accumulation of GAGs and deficiency of IDS activity. 10
Viral vectors are powerful tools for the transduction of therapeutic genes into various cells. Ex vivo HSC gene therapy with lentiviral vectors has been used as a clinical option for several genetic diseases including adrenoleukodystrophy, metachromatic leukodystrophy, Wiskott-Aldrich syndrome, and β-thalassemia. 11 –14 Lentiviral ex vivo gene therapy has also been shown to be effective for correcting certain aspects of neuronal defects in mouse models of MPS I and MSP IIIA. 15,16 It appears, therefore, that enhanced secretion of IDS from HSCs is an attractive approach to ameliorate CNS lesions in MPS II. However, the effects of lentivirally transduced HSCT on CNS disease in MPS II have not been clarified. In this study, we demonstrated that lentiviral isogeneic HSC gene therapy corrected both the biochemical abnormalities in the brain and the deterioration of neurological function in the MPS II mouse model.
Materials and Methods
Animals
Congenic C57BL/6 female mice, 17 which are heterozygous for the IDS gene on the X-linked allele (IDS+/–), were kindly provided by J. Muenzer (University of North Carolina, Chapel Hill, NC). Female heterozygous mice (IDS+/–) were mated with male wild-type mice to generate IDS knockout (KO) male mice (MPS II mice). Their genotypes were determined by PCR. 10 The animal care and ethics committees of Jikei University School of Medicine (Tokyo, Japan) approved the animal husbandry and other procedures used in the animal experiments.
Lentiviral vector construction and production
Codon-optimized human IDS gene (GenScript, Piscataway, NJ) and enhanced green fluorescent protein (eGFP) gene were inserted into the multiple cloning site of the second-generation lentiviral vector pSMPUR-MCU3-MCS, 18 which was kindly provided by D.B. Kohn (UCLA, Los Angeles, CA). Large-scale production of lentiviral vectors was achieved according to a procedure previously described. 18 Lentiviral titers were determined by enzyme-linked immunosorbent assay of viral p24 proteins. 19
Transduction and transplantation of lineage-negative bone marrow cells
Whole bone marrow cells were harvested from tibias and femurs of donor MPS II mice (8–9 weeks old) and separated into lineage-positive and lineage-negative populations, using a lineage cell depletion kit (Miltenyi Biotec, Auburn, CA). Lineage-negative cells were transduced at a multiplicity of infection of 50 with lentiviral vectors carrying the IDS or eGFP gene for 16 hr on a RetroNectin-coated 6-well plate at a density of 2×106 cells/ml in Dulbecco's modified Eagle's medium (DMEM; Wako Pure Chemicals, Tokyo, Japan) with 10% fetal calf serum, recombinant mouse Flt-3 ligand (rmFlt-3L, 100 ng/ml), recombinant mouse stem cell factor (rmSCF, 100 ng/ml; R&D Systems, Minneapolis, MN), and Polybrene (8 μg/ml; Sigma-Aldrich, St. Louis, MO), as previously described. 20 The infected cells (2×106) were washed with phosphate-buffered saline (PBS; Wako Pure Chemicals) and injected intravenously into IDS-KO recipient mice (n=14) after lethal irradiation (9 Gy; Hitach-MBR-1520-R irradiator), according to a procedure previously described. 10 Blood samples were collected every 4 weeks and mice were perfused and killed 24 weeks after transplantation. Only well-perfused tissues were used in all experiments.
Antibodies
Anti-LC3 antibody was obtained from Novus Biologicals (Littleton, CO). Anti-ubiquitin monoclonal antibody was purchased from Medical and Biological Laboratories (Nagoya, Japan). Anti-ubiquitin polyclonal antibody was obtained from Dako (Glostrup, Denmark). Anti-p62 antibody and anti-GFP antibody were purchased from Abcam (Cambridge, MA). Anti-actin antibody was obtained from Sigma-Aldrich. Anti-SCMAS (subunit C of mitochondrial ATP synthase) antibody was gifted by J. Ezaki (Juntendo University School of Medicine, Tokyo, Japan). Anti-Iba1 antibody was obtained from Wako Pure Chemicals.
Quantitative analysis of proteins and IDS activity
Protein concentrations were estimated with a bicinchoninic acid (BCA) protein assay kit (Thermo Fisher Scientific, Rockford, IL) standardized with bovine serum albumin. IDS activity was assayed with artificial substrate 4-methylumbelliferyl-α-iduronide-2-sulfate, according to methods previously described. 21
Total GAG assay and pathological iduronic acid assay
Total GAG assays were performed with a Wieslab sulfated GAG (sGAG) quantitative Alcian blue-binding assay kit (Euro-Diagnostica, Malmö, Sweden), as previously described. 21 The pathological iduronic acid (PIA) assay was performed according to the procedure described by Shimada and colleagues. 22 Briefly, GAGs in cerebral homogenates were isolated by passage through an anion-exchange column and digested with recombinant iduronidase and recombinant IDS (a gift from Genzyme Japan). Disease-specific iduronic acid from the nonreducing terminal of heparan sulfate and dermatan sulfate was derivatized with ethyl 4-aminobenzoate (ABEE) by reductive amination, and measured against standard curves of the authentic compound, using high-performance liquid chromatography (HPLC).
SDS-PAGE and Western blot analysis
Cerebral tissues were homogenized in 9 volumes of 50 mM Tris-HCl, pH 7.5 (Tris buffer), containing 1% Triton X-100 and protein inhibitor cocktail (PIC; Roche Diagnostics, Indianapolis, IN), followed by sonication and centrifugation at 18,000×g for 1 hr at 4°C. Residual pellets were lysed with 7 volumes of Tris buffer containing 2% sodium dodecyl sulfate (SDS) and PIC, followed by sonication and centrifugation, as previously described. The supernatants were identified as the detergent-insoluble fractions. Alternatively, whole tissue extracts from cerebra were obtained according to a procedure previously described. 23,24 Samples containing equal amounts of proteins were resolved by SDS-PAGE on 4–20% acrylamide gradient gels and transferred onto nitrocellulose membranes. The membranes were dipped in blocking buffer (Tris buffer containing 150 mM NaCl, 0.1% gelatin, 0.1% casein, and 0.05% Tween 20) and incubated with each primary antibody. After a brief wash, membranes were immunostained with peroxidase-labeled secondary antibody (Nichirei, Tokyo, Japan) and detected with ImmunoStar LD (Wako Pure Chemicals).
Flow cytometry
We used antibodies specific for CD11b–phycoerythrin (PE) (clone M1/70), Ly-6G-PE (clone RB6-8C5), CD45R-PE (clone RA3-6B2), and CD3e-PE (clone 145-2C11) (eBioscience, San Diego, CA). Peripheral white blood cells (WBCs) were obtained from recipient mice transplanted with eGFP-overexpressing HSCs. Cells were incubated with Fc blocker (CD16/32; eBioscience) and stained to obtain the expression of surface markers. 25 Data were obtained with a MACSQuant analyzer (Miltenyi Biotec, Bergisch Gladbach, Germany) and analyzed with MACSQuantify software (Miltenyi Biotec).
Immunohistochemistry
Preparation of brain sections and immunostaining were performed according to procedures previously described. 21,26 Sliced sections were incubated with the primary antibodies and visualized with Alexa 555-conjugated anti-rabbit IgG, Alexa 488-conjugated streptavidin (Invitrogen, Carlsbad, CA), and biotinylated anti-chicken IgY (H+L) (GeneTex, Irvine, CA) secondary antibodies.
Quantitative RT-PCR
Vector copy number in livers and cerebra were determined by analysis of lentiviral packaging sequences in genomic DNA, using quantitative real-time PCR as previously described. 19
Y-maze test
The behaviors of wild-type, MPS II, and transplanted MPS II mice (9 and 33 weeks old) were assessed by Y-maze testing as previously decribed. 21
Statistical analysis
Statistical analysis was performed with JMP statistical software version 10 (SAS Institute, Cary, NC). The Student t test was used in this study. A p value less than 0.05 was considered statistically significant.
Results
Lentiviral ex vivo HSC gene therapy increases IDS activities in MPS II tissues
To investigate the effect of lentiviral gene delivery to HSCs, we transfected lineage-negative cells derived from IDS-deficient bone marrow cells with a lentiviral vector encoding IDS and analyzed the activities of the enzyme. Lentiviral gene transduction drastically increased IDS activity in lineage-negative cells derived from MPS II mice (nontransduced, 45.7 nmol/4 hr/mg protein vs. transduced, 7418.9 nmol/4 hr/mg protein).
Next, we transplanted lentiviral IDS-transduced MPS II HSCs into lethally irradiated recipient IDS-KO mice and analyzed the activities of the enzyme in samples from several tissues including liver, heart, cerebrum, and serum (n=4–8, respectively). From 2 weeks after transplantation, the levels of IDS activity in serum were increased from 0 to 4767.1 nmol/4 hr/mg protein and supernormal levels (approximately 15-fold higher than wild type) were sustained throughout the 24 weeks of treatment (Fig. 1A). Supernormal IDS activity was also observed in liver (approximately 11-fold higher than wild type) and heart (approximately 3-fold higher than wild type) derived from recipient MPS II mice (Fig. 1B). Furthermore, transplantation of IDS-overexpressing HSCs into MPS II mice significantly increased the activities of the enzyme in cerebral tissues (approximately 2.9% of wild-type activity) (Fig. 1B). These results indicated that lentiviral ex vivo HSC gene therapy increased IDS activity both in peripheral tissues and the CNS in MPS II mice.

Lentiviral ex vivo hematopoietic stem cell (HSC) gene therapy increases iduronate-2-sulfatase (IDS) activity in mucopolysaccharidosis type II (MPS II) tissues.
Lentiviral ex vivo HSC gene therapy corrected the accumulation of GAGs in MPS II tissues
To evaluate the therapeutic outcome of IDS-overexpressing isogeneic HSCT, we analyzed the amounts of GAGs in livers, hearts, and cerebra of recipient MPS II mice (33 weeks old, n=5–8, respectively) by Alcian blue staining. The levels of total GAGs in liver and heart tissue of MPS II mice were normalized by lentiviral HSC gene therapy treatment (Fig. 2A). In contrast, whereas the levels of total GAGs in cerebra were slightly increased in MPS II mice compared with wild type, no significant difference was observed between untreated and transplanted mice (Fig. 2A). We previously demonstrated that Alcian blue staining is not sensitive or specific enough to precisely evaluate the accumulated levels of GAGs in MPS II mouse cerebra; however, this can be achieved using the pathological GAG assay. 10,27 In this study, we analyzed PIA derived from the nonreducing end of disease-specific GAGs in the cerebra of MPS II mice (each group, n=5) because the PIA assay could quantify the amount of disease-specific GAGs as accurately as the pathological GAG assay. 22 The PIA assay showed a drastic increase in the levels of PIA in the cerebra of MPS II mice and a significant reduction in diseased mice transplanted with lentivirally transduced HSCs (Fig. 2B). These results indicated that lentiviral ex vivo HSC gene therapy reduced the accumulation of GAGs both in peripheral tissues and the CNS in MPS II mice.

Lentiviral ex vivo HSC gene therapy corrects the accumulation of glycosaminoglycans (GAGs) in MPS II tissues.
Lentiviral ex vivo HSC gene therapy rescued autophagic dysfunction in MPS II cerebra
Defective autophagy is a prominent feature of lysosomal storage disorders. 28 In MPS, lysosomal enzyme deficiency has been shown to induce retardation of autophagic flux in the brain of the mouse model of MPS IIIA, 29 which results in heparan sulfate accumulation, similar to the effects in MPS II. To investigate whether impaired autophagy can be observed in the brains of MPS II mice, we analyzed the levels of autophagic marker LC3 in cerebra, using Western blot analysis (each group, n=3). The levels of LC3-II, a major component of autophagosomes, were higher in MPS II mice than in the wild type (Fig. 3A). We then analyzed autophagy substrates such as p62, detergent-insoluble ubiquitin–protein conjugates, and SCMAS. The levels of both p62 and Triton X-100-insoluble ubiquitin–protein conjugates were increased in MPS II mouse cerebra compared with the wild type (Fig. 3A and B). Moreover, immunohistochemical analysis revealed neuronal accumulation of ubiquitin–protein conjugates and SCMAS in cerebral tissues from MPS II mice (Fig. 3C), indicating that autophagic flux is impaired in the MPS II mouse brain.
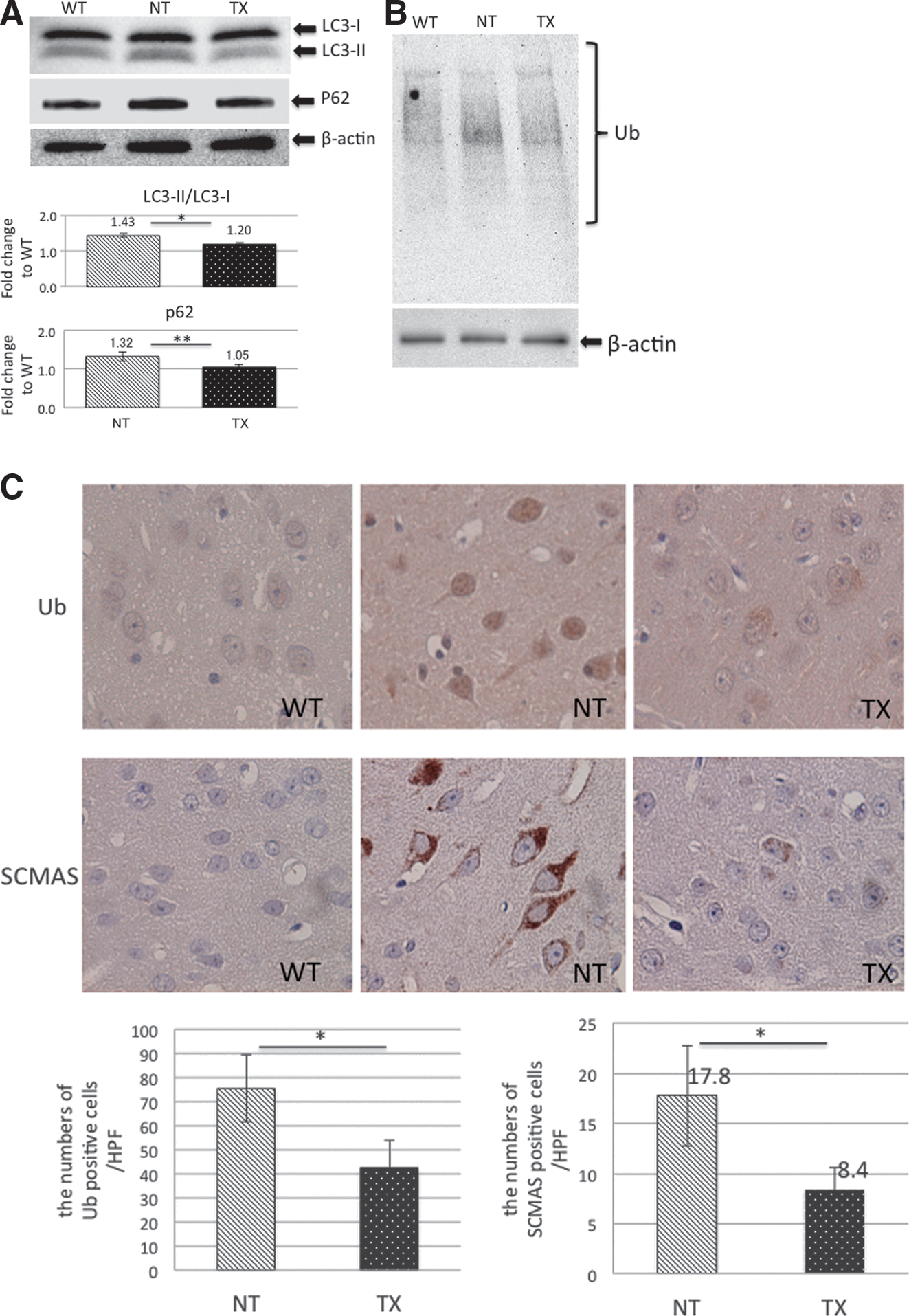
Lentiviral ex vivo HSC gene therapy rescues autophagic dysfunction in the MPS II brain.
We subsequently investigated the effects of IDS-overexpressing isogeneic HSCT on autophagic dysfunction in the brains of MPS II mice. The levels of LC3-II, p62, and detergent-insoluble ubiquitinated proteins in MPS II mouse were reduced by lentiviral HSC gene therapy treatment (Fig. 3A and B). Furthermore, a positive effect of genetically modified HSCT on neuronal accumulation of ubiquitin–protein conjugates and SCMAS was observed in MPS II mouse cerebra (Fig. 3C). These results indicated that lentiviral ex vivo HSC gene therapy rescued autophagic flux in the MPS II mouse cerebrum.
Lentiviral ex vivo HSC gene therapy prevented neurofunctional deterioration in MPS II mice
MPS II mice were analyzed by the Y-maze test, which evaluates short-term memory performance, to investigate the effect of IDS-overexpressed HSCT on behavioral abnormalities (each group, n=3–5). 21,30,31 Although spontaneous alternation behavior did not differ between MPS II mice and wild-type mice at 9 weeks of age, rates were impaired in a time-dependent manner in MPS II mice (Fig. 4). On the other hand, transplantation of IDS-overexpressed HSCs inhibited the deterioration of alternation behavior in MPS II mice (Fig. 4). These results indicated that lentiviral ex vivo HSC gene therapy prevented neurofunctional impairments in MPS II mice.
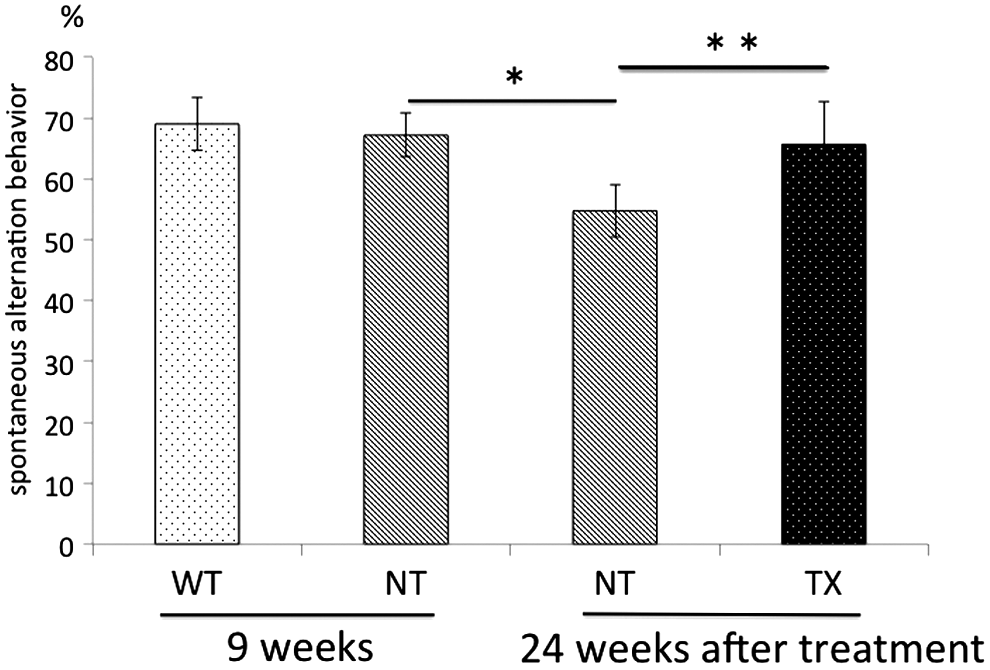
Lentiviral autologous HSC gene therapy prevents neurofunctional deterioration in MPS II mice. Spontaneous alternation behaviors of wild-type (WT), MPS II (NT), and HSC-transplanted MPS II (TX) mice were analyzed by a Y-maze test before (n=3, respectively) and 24 weeks after treatment (n=5, respectively). Data are expressed as means±SD. Statistical analyses of data were performed by Student t test (*p=0.01, **p<0.05).
Gene-modified HSCs differentiated into hemocyte fractions in peripheral blood
We analyzed GFP expression in the peripheral blood of recipient mice 12–14 weeks after transplantation to confirm whether the transplanted gene-modified HSCs differentiated into hemocyte fractions (n=4). We detected GFP expression in 29.5% of total peripheral WBCs, 31.6% of monocytes (CD11b positive), 52.2% of granulocytes (Ly-6G positive), 20.4% of B cells (GD45R), and 27.6% of T cells (CD3e positive). These results indicated that the transplanted gene-modified HSCs differentiated into cell lineages.
Some gene-modified HSCs translocate to the brain and differentiate into microglial cells
To investigate the therapeutic mechanisms of IDS-overexpressing HSCT in the MPS II mouse brain, we analyzed the lentiviral copy number in genomic DNA from cerebral tissues of transplanted mice, using quantitative RT-PCR (each group, n=5). Whereas 0.045 copy per cell was observed in the livers, the average copy number was 0.0063 copy per cell in the cerebra of MPS II mice treated with IDS-transduced HSCs, which indicated invasion of HSCs into the cerebra (Fig. 5A).
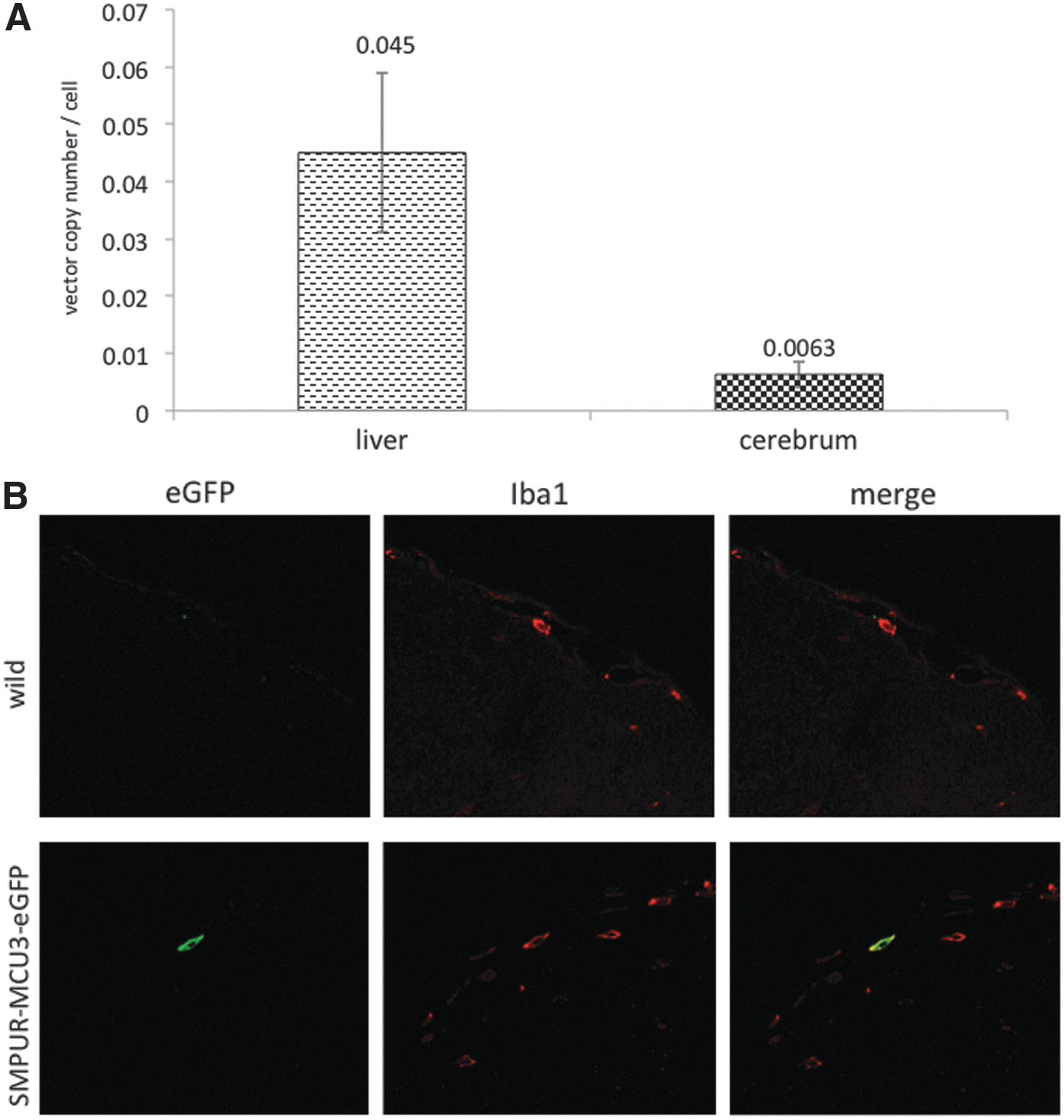
Gene-corrected HSCs translocate to the brain and differentiate into microglial cells in MPS II mice.
Next, to determine whether transplanted HSCs differentiate into microglial cells, we transplanted HSCs overexpressing eGFP into MPS II mice, and analyzed the localization of eGFP in cerebral tissues by fluorescence immunohistochemistry. eGFP fluorescence was observed in cerebra of recipient mice, and was colocalized with some microglial marker Iba1-positive cells (Fig. 5B). These results strongly suggest that lentiviral ex vivo HSC gene therapy imparts a therapeutic effect in CNS disease in MPS II mice through the differentiation of HSCs into microglial cells, which allows for enzymatic cross-correction.
Discussion
We have shown that lentiviral ex vivo HSC gene therapy improves the biochemical abnormalities in diseased tissues of MPS II mice, including the cerebrum (Fig. 1). In addition, we found that gene-corrected HSCs rescued autophagic flux retardation and neurofunctional impairments in the CNS of MPS II mice. To our knowledge, this is first report to show that ex vivo gene therapy with lentiviral vectors can correct CNS disease in MPS II.
HSCT was shown to ameliorate disease symptoms of MPS I in the 1980s, and is now a standard treatment for the disorder. 32 In particular, HSCT alleviates several manifestations including neurological deterioration in patients with MPS I when performed at an early age (<2 years old). 33 HSCT became a treatment option for patients with MPS II, because MPS II is similar to MPS I in terms of the accumulation of heparan sulfate and neurodegenerative disease, and a retrospective study of Japanese patients with MPS IIB showed efficacy in the treatment of CNS and valvular lesions. 34 However, some studies have reported that transplantation of HSCs is not effective for CNS disease in patients with MPS II 7 –9 and this notion is supported by our study, which showed that BMT of wild-type donor cells cannot correct biochemical abnormalities, including GAG accumulation and enzyme activity defects, in the brains of MPS II mice. 10 The efficacy of BMT in patients with MPS II remains controversial. On the other hand, this study demonstrated that lentiviral isogeneic HSC gene therapy improved biochemical and neuropathological alterations in the cerebra of MPS II mice (Figs. 1B, 2, and 3), suggesting that the levels of IDS secreted by naive HSCs are not sufficient to rescue CNS lesions in MPS II. Indeed, IDS-overexpressing HSCT leads to supraphysiological enzyme activity in the sera of MPS II mice (Fig. 1A), whereas transplantation of allogeneic normal bone marrow cells achieves only 20% of the enzymatic activity of wild-type enzymes. 10 It appears, therefore, that ex vivo gene therapy with lentiviral vectors can overcome the limits of HSCT in MPS II.
Our study showed that lentiviral ex vivo HSC gene therapy increased the levels of IDS activity in the cerebra as well as in liver and heart tissues of MPS II mice (Fig. 1B). Some transplanted HSCs pass through the BBB and differentiate into microglial cells; hence HSCT has been shown to cure several neuropathic lysosomal storage diseases including metachromatic leukodystrophy (MLD). 5 Indeed, we found the presence of vector sequences in genomic DNA from cerebral tissues of MPS II mice treated by IDS-corrected HSCT (Fig. 4A). In addition, we detected the colocalization of eGFP with microglial marker Iba1 in the cerebra of MPS II mice transplanted with eGFP-overexpressing HSCs (Fig. 4B). Therefore, it seems that the increased levels of IDS activity in the cerebra of MPS II recipients might be attributed to cross-correction via HSC-derived microglial cells. Furthermore, another contributing factor to the increase in enzyme levels in the CNS could result from enforced passage through the BBB because of high levels of enzymes in the serum. Our study showed that despite donor chimerism in total peripheral blood being approximately 30%, the levels of IDS activity in serum of MPS II after lentiviral HSC gene therapy were 15-fold higher than that of wild type, indicating that the levels of enzyme produced per IDS-overexpressed cell might be approximately 50-fold higher than that of normal blood cells. A previous study showed that 5-fold higher enzyme levels in the sera of wild-type mice, achieved by AAV2/5 in vivo gene therapy, could pass through the BBB in MPS II mice 35 and that systemic administration of high-dose recombinant IDS could transiently raise enzyme levels in the brain. 36 Our treated mice showed more than 15-fold greater enzyme activity continuously in serum, which may have contributed to the increased enzyme levels in the CNS.
Our results showed that transplantation of IDS-overexpressing HSCs improved enzyme activity in the cerebra of MPS II mice to approximately 2.9% of the activity found in the wild type, although clearly this is not a dramatic increase (Fig. 1B). Consistent with the results of the enzyme activity assays, we confirmed that small numbers of lentiviral vector copy and eGFP-positive microglial cells were present in the cerebra of MPS II recipient mice (Fig. 5). These observations suggested that only a few transplanted HSCs were recruited to the CNS in our MPS II mice. In wild-type mice, HSCT is reported to replace less than 5% of cerebral microglial cells with cells of HSC origins. 37 Moreover, the recruitment of HSCs to the brain has been shown to be promoted by neuronal injury and neurodegeneration accompanied by the production of cytokines and chemokines. 6,37 –39 It seems that the low recruitment of HSCs into the MPS II brain may be due to insufficient neurodegeneration for triggering the replacement of microglial cells in recipient mice. Our results support this notion because we found no obvious differences in neuronal function between wild-type and pretransplanted MPS II mice (Fig. 4). In this study, we detected GFP expression in 31.6% of CD11b-positive cells from MPS II mice after transplantation of gene-corrected HSCs. Biffi and colleagues reported the expression of GFP in approximately 80% of CD11b-positive cells and in approximately 25% of microglial cells derived from MLD mice transplanted with lentivirally GFP-transduced HSCs. 40 They also showed that approximately 10% of wild-type enzyme activity was observed in the cerebrum of MLD mice treated by lentiviral HSC gene therapy. 41 Therefore, the efficacy of transduction of our vector to HSCs may be another contributing factor to relatively low IDS activity in the brains of MPS II mice transplanted with IDS-overexpressing HSCs. In addition, a previous report showed that lentiviral ex vivo gene therapy using the CD11b promoter achieved high GFP expression in CD11b-positive cells and more than 10% of wild-type N-sulfoglucosamine sulfohydrolase (SGSH) activity in treated MPS IIIA mouse brains. 42 This method could contribute to an improvement in our results.
We found that approximately 2.9% of wild-type IDS activity decreased the amounts of PIA by about 65% in cerebra of MPS II mice, but did not result in complete normalization (Fig. 2). In several lysosomal storage diseases such as GM1 gangliosidosis and MPS IVB, at least 10% of normal enzyme activity is required to clear the accumulated substrates and prevent the onset of disease. 43,44 Moreover, the above-described level of sulfamidase activity normalized the accumulation of heparan sulfate in the brains of MPS IIIA mice transplanted with lentiviral gene-modified HSCs. 16 It appears, therefore, that nearly 10% of normal IDS activity is required to achieve complete clearance of PIA in MPS II, which is similar to other lysosomal storage diseases. On the other hand, our results revealed that a small amount of IDS activity improved the accumulation of autophagy-related proteins such as LC3-II, p62, detergent-insoluble ubiquitin–protein conjugates, and SCMAS in the cerebra of MPS II mice (Fig. 3). Excessive lysosomal accumulation of undegraded substrates has been shown to induce defective lysosome–autophagosome fusion and block autophagic flux in many lysosomal storage diseases, including MPS IIIA and multiple sulfatase deficiency. 29 Secondary accumulation of autophagic substrates, rather than the primary accumulation of lysosomal substrates, is regarded as a key determinant of cell death. Consistent with the above model of disease pathogenesis, we found that IDS activity prevented not only secondary accumulation of p62, detergent-insoluble ubiquitin–protein conjugates, and SCMAS but also deterioration in the alternation behavior of MPS II mice treated with HSC gene therapy (Figs. 3 and 4). Hence, it seems that even if IDS does not completely eliminate GAG accumulation, the enzyme may contribute to suppression of disease progression in MPS II through maintaining autophagic flux.
Overexpression of galactocerebrosidase in HSCs is known to be toxic and results in functional impairment and apoptosis. 45 However, IDS overexpression in HSCs did not cause these problems and we observed reconstitution of each cell lineage. Moreover, we also measured arylsulfatase A and arylsulfatase B in IDS-overexpressing 293A cells, because the activity of other sulfatases can be reduced due to excessive consumption of formylglycine-generating enzyme (FGE). 46,47 The activities of these two sulfatases did not reduce this effect (data not shown).
The therapeutic potential and safety of lentiviral HSC gene therapy has been reported for several animal models of lysosomal storage disorders, such as globoid cell leukodystrophy and Farber disease. 48 In particular, the efficacy of ex vivo HSCT as a lentiviral vector for patients with adrenoleukodystrophy and metachromatic leukodystrophy has already been demonstrated in clinical trials, 11,12 indicating that lentiviral ex vivo HSC gene therapy is a realistic therapeutic strategy for lysosomal storage diseases. In this study, we showed that lentiviral gene correction of autologous HSCs from MPS II mice improved the manifestations of neuronal disease including biochemical and functional abnormalities, which are resistant to other therapeutic approaches including allogeneic normal HSCT and ERT. Our results have provided evidence that lentiviral autologous HSC gene therapy has the potential to become an encouraging option for the treatment of CNS disease in patients with MPS II.
The limitations of this study included gene toxicity and the confirmation of transduction in stem cells. Gene toxicity is a serious problem because of the possibility of inducing malignant tumors via gene therapy in patients with X-linked severe combined immunodeficiency (X-SCID) and chronic granulomatous disease (CGD). 49,50 In addition, clonal expansion was observed in β-thalassemia treated by lentiviral ex vivo gene therapy. 51 We will be conducting further evaluation and analysis of the integration sites in our future studies.
Footnotes
Acknowledgments
T.W., Y.S., H.K., and T.O. designed and conducted research; T.W., Y.S., T.H., T.F., and H.K. performed research and analyzed the data; T.W. wrote the manuscript; and Y.S., H.K., Y.E., H.I, and T.O. discussed data and edited the paper. The authors are indebted to Joseph Muenzer (UNC Health Care) for providing MPS II mice, to Donald B. Kohn (UCLA) for providing lentiviral vector, and to Dr. Junji Ezaki (Juntendo University School of Medicine, Japan) for providing anti-SCMAS antibody. The authors also thank Sayoko Iizuka and Eiko Kaneshiro (Jikei University School of Medicine) for excellent technical assistance, and members of Laboratory Animal Facility (Jikei University School of Medicine) for helping with animal studies. This research was supported by a Health and Labor Sciences Research Grant.
Author Disclosure Statement
T.O. and Y.E have active research support from Genzyme Japan Co., Ltd. and Shire Japan Co., Ltd. H.I. has active research support from Genzyme Japan Co., Ltd. and Dainippon Sumitomo Co., Ltd. These activities have been fully disclosed and are managed under a Memorandum of Understanding with the Conflict of Interest Resolution Board of the Jikei University School of Medicine. T.W., Y.S., K.A., T.H., T.F., and H.K have no potential COI to disclose.