
Abstract
The main obstacle to viral vector-mediated gene therapy remains the elicitation of an immune response to the vector, resulting in clearance of transgene and resistance to further transgenesis. Specific antibody production contributes to such immune responses. A single class II-restricted epitope of adenovirus serotype 5 (Ad5) vector hexon-6 capsid protein containing a thiol-oxidoreductase motif was used in an attempt to prevent specific antibody production in response to Ad5 vectors. We demonstrate here that such immunization carried out before intravenous administration of Ad5 vectors prevents antibody production to the ensemble of Ad5 vector proteins in both BALB/c and C57BL/6 mice. The antibody response to Ad5 is dependent on innate immune activation, seemingly involving natural killer T (NKT) cells. We observed that immunization with a class II-restricted Ad5 peptide prevents such NKT cell activation. Increased transgenesis and prolonged transgene expression result from such immunization, providing a simple protocol for improving gene therapy.
Introduction
V
Therefore, preventing the immune response to a viral vector remains a significant unmet medical need. Ideally, the method should be strictly antigen-specific and efficient over the long run, allowing the chance to readminister the construct if required. Our laboratory has developed a methodology by which an immune response to a viral vector can be prevented. The principle relies on the reduction of a disulfide bridge in the CD4 molecule, using class II-restricted epitopes flanked with a four-amino acid motif containing thioreductase activity. This results in increased synapse formation and in CD4+ T cells acquiring the capacity to induce apoptosis of antigen-presenting cells with which a cognate synapse is formed, as well as inducing apoptosis of CD4+ T cells activated at the surface of the same antigen-presenting cells. The overall effect is prevention as well as suppression of the production of specific antibodies through elimination of effector CD4+ T cells, even when these are already polarized. 13 The first results obtained in the setting of the prevention of the immune response to an adenoviral vector are presented here, including preliminary data suggesting that such prevention has the potential to improve transgenesis and prolong transgene expression.
We deliberately decided to work with adenoviral vectors, both for the advantages they provide for the practice of gene therapy (and possibly gene vaccination), and for their strong immunogenicity.
Materials and Methods
Peptides
The hexon protein sequence (hexon 6) from adenovirus serotype 5 (Ad5) vector was obtained from UniProt (see
Fluorescence-based assay for thioreductase activity
A fluorescein isothiocyanate (FITC)-NH-Gly-Cys-Asp-COOH peptide was synthesized (Eurogentec) and self-quenched by solubilization in dimethyl sulfoxide (DMSO) [(FITC-Gly-Cys-Asp)ox]. The reduction of 2.5 μM (FITC-Gly-Cys-Asp)ox was monitored on a 96-well plate for 40 min (25°C) after incubation in phosphate-buffered saline (PBS) with peptides (25 μM) or 2 mM dithiothreitol (DTT). Reduction was measured as the increase in fluorescence at 530 nm after excitation at 494 nm on a multimode microplate reader (Synergy HT; BioTek Instruments, Winooski, VT).
Mice
Female C57BL/6 (H2b ) and BALB/c (H2d ) mice were obtained from the animal facility of the University of Leuven (Leuven, Belgium). Female CD1d knockout (KO) mice (C.129S2-Cd1tm1Gru /J) on the BALB/c background were purchased from Jackson Laboratory (Bar Harbor, ME). Throughout the experiments, mice were maintained in the animal facility of the University of Leuven. All experiments were approved by the animal ethics committee and performed in accordance with European regulations.
In vivo studies
Mice (6 to 8 weeks old) received four footpad injections (on days 1, 15, 22, and 29) of peptides (50 μg/mouse) containing either the thioredox motif or the alanine loss-of-function substitute in aluminum hydroxide adjuvant (alum; Life Technologies, Ghent, Belgium) or alum only as control (Ctrl). Seven days after the last immunization, mice were administered intravenously 109 plaque-forming units (PFU) per mouse of either Ad5-GFP or Ad5-null vector (Vector BioLabs, Malvern, PA) via the tail vein, and administration was repeated after 10 days when indicated. Blood samples were collected 10 or 14 days after each administration of Ad5 vector as specified. To analyze the proportions of natural killer and CD8+ T cells in the liver, mice were killed 14 days after the last Ad5 administration.
Humoral response
To quantitate Ad5-specific antibody production, 96-well plates (Greiner Bio-One, Vilvoorde, Belgium) were coated with Ad5 vector at 1.6 × 107 PFU/ml in 1 M NaHCO3/Na2CO3, pH 9.5, overnight at 4°C. After washing, a “blocking” step was included with 2% bovine serum albumin (BSA)–PBS for 1 hr, followed by dilutions of serum samples in PBS and calibration samples for a further incubation of 90 min at room temperature. After washing, plates were probed by addition of horseradish peroxidase (HRP)-conjugated anti-mouse total IgG (Bio-Rad, Brussels, Belgium), anti-IgG1 (BD Biosciences, Erembodegem, Belgium), anti-IgG2a (BD Biosciences), or anti-IgG2b (Santa Cruz Biotechnology, Heidelberg, Germany) for 1 hr. The anti-mouse IgG2a antibody cross-reacts with IgG2c of C57BL/6 mice. 3,3′,5,5′-Tetramethylbenzidine (TMB) substrate (BD Biosciences) was added and absorbance was read at 450 and 650 nm. To determine acute inflammation elicited by innate immunity activation, blood samples were collected before and after (6 and 24 hr) Ad5 administration, and were assessed with ELISA kits for the presence of interferon (IFN)-α (USCN, Schilde, Belgium), IFN-γ (BD Biosciences), and interleukin (IL)-6 (BD Biosciences).
Cell isolation
Spleens were flushed with RPMI medium, and red blood cells were eliminated with ACK (ammonium chloride/potassium) lysing buffer. Lymphocytes were then separated on a Ficoll density gradient. Liver cells were collected by tissue digestion with collagenase buffer containing collagenase (100 U/ml; Sigma-Aldrich, Diegem, Belgium), DNase (50 U/ml), and proteinase inhibitor (Life Technologies) in 3 mM CaCl2. Liver mononuclear cells were then separated by using Percoll (GE Healthcare, Uppsala, Sweden) followed by centrifugation at 1052 g for 20 min at 25°C.
Generation of immature dendritic cells from bone marrow
Naive C57BL/6 mice were killed and bone marrow (BM) cells were harvested from femurs and tibias. Progenitors of dendritic cells (DCs) were obtained by depleting CD19+ cells (B cells) and CD90.2+ cells (T cells), using microbeads (Miltenyi Biotec, Leiden, The Netherlands). Cells were then cultured in AIM-V medium supplemented with 50 mM 2-mercaptoethanol and granulocyte-macrophage colony-stimulating factor (GM-CSF, 500 U/ml). Medium was renewed every 2 days, and immature DCs were collected after 1 week and were either stored in liquid nitrogen or matured (with tumor necrosis factor [TNF]-α, 10 ng/ml; overnight).
CD4+ T cell proliferation assay
In a cell proliferation test, T cells (3 × 104 cells/200 μl of AIM-V medium/well) were cocultured at a ratio of 3:1 with CytoRed cell tracer-stained DCs, in the presence of serial concentrations (10× dilution; 0–10 μM) of ccHexBL/6 peptide. The proliferating T cells were then quantitated by fluorescence-activated cell sorting (FACS), using anti-Ki-67–V450 (BD Biosciences, Erembodegem, Belgium) staining of the CytoRed-negative T cell population after 72 hr. T cell surface (using anti-CD3–APC [eBioscience] and anti-CD4–APC-H7 [BD Biosciences] antibodies) and intracellular staining (using anti-Ki-67–V450 antibody) was performed according to the manufacturer's instructions.
Cell phenotyping
Standard protocols for FACS staining were followed after exclusion of dead cells and blockage with anti-mouse CD16/32 antibody (eBioscience) to prevent nonspecific binding. To quantify the proportion of natural killer T (NKT) and CD8+ T cells from the liver, staining included anti-CD8–FITC (eBioscience), anti-CD3–PE (eBioscience), anti-CD19–PerCP (eBioscience), anti-NKG2D–APC (eBioscience), and anti-CD4–APC-H7 (BD Biosciences). To quantify the proportion of effector memory CD4+ T cells, surface staining included anti-CD44–V450 (BD Biosciences), anti-CD62L–PE (eBioscience), anti-CD8–PerCP (eBioscience), anti-CD3–APC (eBioscience), and anti-CD4–APC-H7 (BD Biosciences) antibodies. At least 50,000 events were acquired and analyzed by flow cytometry (MACSQuant; Miltenyi Biotec).
Derivation of cytolytic CD4+ T cell clones
To induce cytolytic CD4+ T cells, C57BL/6 mice were immunized four times (on days 1, 15, 21, and 28) with ccHexBL/6 peptide or aaHexBL/6 peptide intraperitoneally. Two weeks after the last immunization, mice were killed and spleens were removed. CD19+ and CD8+ cells were then depleted from splenocytes and the rest of the cells were stimulated with mitomycin C-pretreated, peptide-loaded DCs. To expand specific cell populations before phenotypic assessment, three consecutive stimulations at an interval of 10 days were performed.
APC cell death assay
DCs were first stained with CytoRed according to the manufacturer's instructions. Peptide aaHexBL/6 (1 μM) was added to 104 DCs before coculturing overnight with 2 × 104 specific CD4+ T cells generated with either ccHexBL/6 or aaHexBL/6 epitope. DCs only, or DCs cocultured with T cells in the absence of peptide, were used as controls. After 18 hr, the cells were stained with annexin-V–APC for 15 min at room temperature in the dark. An additional volume of annexin binding buffer (100 μl/well; BD Biosciences) was added before centrifugation. The cells were resuspended in 0.2% BSA–PBS (100 μl/well) and analyzed by flow cytometry.
Immunohistochemistry
Mice were killed and livers were perfused with PBS via the inferior vena cava before being removed. After fixation with formaldehyde (4%), 8-μm-thick cryosections were prepared. Green fluorescent protein (GFP) expression was then evaluated by immunohistochemistry. To quantitate GFP+ cells in the liver, cryosections were incubated overnight with rabbit anti-GFP (diluted 1:400) and then probed for 45 min with goat anti-rabbit IgG–HRP (diluted 1:300) (both from Life Technologies). Signals were developed by addition of diaminobenzidine (Sigma-Aldrich). To quantitate Kupffer cells in liver sections, cryosections were incubated overnight with rat anti-mouse F4/80 (AbD Serotec, Puchheim, Germany) at a 1:100 dilution and then stained with rabbit anti-rat IgG–HRP (Dako, Heverlee, Belgium) at a 1:300 dilution. After washing, signals were detected with Cy3 tyramide (PerkinElmer, Zaventem, Belgium). GFP+ cells and Kupffer cells were identified and enumerated by microscopy (Axio Imager; Carl Zeiss, Göttingen, Germany).
Statistical analysis
Data are expressed as means ± SEM. The Mann–Whitney U test was applied to compare two experimental groups. When data were not normally distributed or when more than two groups were evaluated, the log10 concentration was used for one-way analysis of variance (ANOVA) with Bonferroni correction. Values of p equal to or less than 0.05 were considered significant.
Results
Peptide design and characterization
The development of an immune response to Ad5 vector is directed primarily against hexon proteins,
5,13
and the presence of anti-hexon antibodies is thought to represent the main hurdle to transgenesis. We selected MHC class II-restricted epitopes of hexon 6 for both C57BL/6 and BALB/c strains and produced peptides encompassing the epitopes plus a thioredox motif located in flanking residues. The enzymatic activity of such peptides was assessed toward a fluorescent substrate. Supplementary Fig. S1 (supplementary data are available online at
MHC class II-restricted hexon epitope containing a thioredox motif elicits antigen-specific cytolytic CD4+ T cells
C57BL/6 mice were immunized with ccHexBL/6 or aaHexBL/6. CD4+ T cells were then isolated from splenocytes and stimulated with mitomycin C-treated DCs loaded with the corresponding epitopes. Stimulation was repeated four times at intervals of 10 days. Conversion to the effector memory cytolytic CD4+ T cell phenotype is shown in Fig. 1A. Ninety percent of the CD4+ T cells exposed to ccHexBL/6 adopted a CD44hiCD62L– phenotype characteristic of effector memory cells, whereas cells exposed to aaHexBL/6 showed only partial conversion (29%; Fig. 1B). CD4+ T cells isolated from the spleen of naive mice or from mice treated only with alum failed to grow in this setting (data not shown). ccHexBL/6-stimulated cells proliferated vigorously in response to ccHexBL/6 epitope exposure (Fig. 1C). In contrast, cells stimulated with the aaHexBL/6 epitope did not expand in response to aaHexBL/6 peptide (Fig. 1C).

ccHexBL/6 peptide immunization elicits an effector memory phenotype with acquisition of cytolytic properties. C57BL/6 mice were immunized four times with ccHexBL/6 peptide or aaHexBL/6 peptide (50 μg/mouse; intraperitoneal). Fourteen days after the last immunization, CD4+ T cells were isolated from splenocytes and stimulated with mitomycin C-treated dendritic cells (DCs) loaded with the corresponding epitope. Stimulation was repeated three times at intervals of 10 days before fluorescence-activated cell-sorting (FACS) analysis. Contour plot for the expression of CD62L and CD44 on CD4+ T cells generated with ccHexBL/6 epitope
Next, we measured the cytolytic capacity of T cells stimulated with ccHexBL/6 epitope on CytoRed-stained DCs. As IFN-γ production is associated with cytotoxicity properties, supernatants were collected 24 hr after the coculture. As measured by ELISA, the production of IFN-γ by T cells stimulated with ccHexBL/6 was 3.8-fold higher (Fig. 1D: p < 0.05, Mann–Whitney U test) as compared with that of T cells stimulated with aaHexBL/6 peptide.
Further, DC apoptosis was determined, showing 22% where cytolytic CD4+ (cCD4+) T cells stimulated with ccHexBL/6 (Fig. 1E) were used, as compared with 14% by CD4+ T cells stimulated with aaHexBL/6 (p < 0.05, Mann–Whitney U test).
It was therefore concluded that addition of the CxxC motif to the hexon-6 MHC class II epitope converted CD4+ T cells into effector memory cells with cytolytic activity toward antigen-presenting cells.
Immunization with a hexon-specific MHC class II peptide containing a thioredox motif prevents anti-Ad5 antibody production
Previous studies have shown that removing key epitopes from the surface of the viral capsid could, to some extent, circumvent preexisting anti-vector immunity and prevent immunization against the viral vector. 3 This approach remains limited because of the large number of such epitopes, both for antibodies and for CD4+ T cells. Antibody prevention through active elimination of specific CD4+ effector cells could potentially be achieved by eliciting cCD4+ T cells.
The thioredox motif-containing peptides were used to immunize C57BL/6 or BALB/c mice, followed by two intravenous injections of 109 PFU of Ad5 vector (Fig. 2A). Control experiments included mice immunized with alum only for each strain, and with a loss-of-function peptide only for the BALB/c strain. Blood samples were collected after each injection of Ad5 vector for specific antibody evaluation.
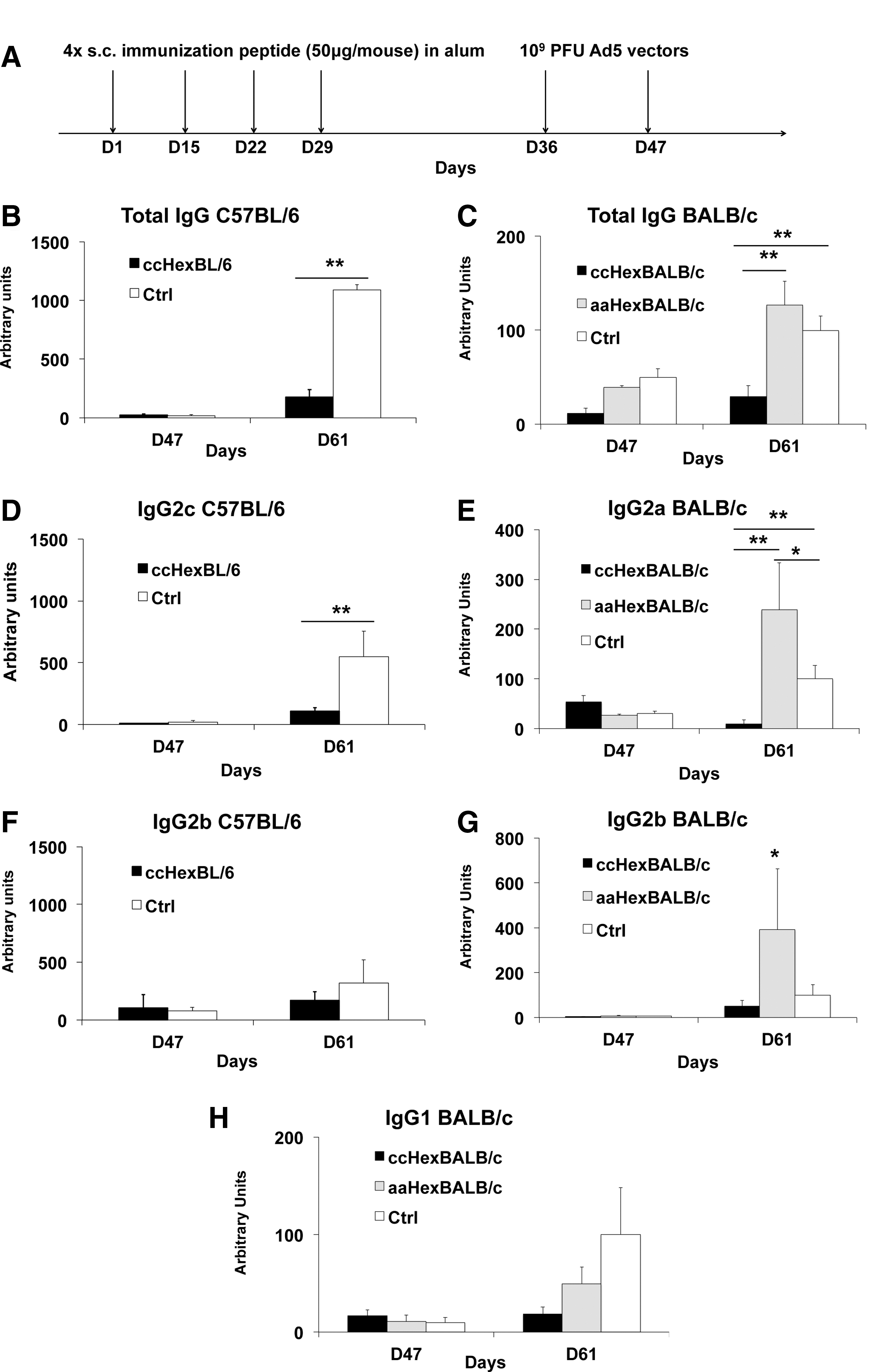
Administration of MHC class II-restricted hexon epitope flanked with a thioredox motif prevents antibody production against the Ad5 vector.
Figure 2B and C show, respectively, 80% (p < 0.01, Mann–Whitney U test) and 71% (p < 0.01, one-way ANOVA) reduction of the concentration of anti-Ad5 total IgG in the thioredox peptide-treated group as compared with the alum-treated group for C57BL/6 mice (Fig. 2B) and BALB/c mice (Fig. 2C). Further, a 77% reduction of total IgG was observed in the ccHexBALB/c group as compared with the aaHexBALB/c group (Fig. 2C).
We further evaluated antibody subclasses, in particular IgG2c and IgG2a antibodies, as they are reported to be the dominating isotype against adenovirus for C57BL/6 and BALB/c, respectively. 14 In C57BL/6 mice, the ccHexC57BL/6-immunized group showed an 80% reduction in IgG2c antibody concentration as compared with the alum-treated group (Fig. 2D: p < 0.01, Mann–Whitney U test). In BALB/c mice, the ccHexBALB/c-immunized group showed a ≥90% reduction in IgG2a concentration as compared with the alum-treated group, and a 95% reduction as compared with the aaHexBALB/c group (Fig. 2E: p < 0.01, one-way ANOVA). A 50% reduction in IgG2b concentration (Fig. 2F and G) was observed in both C57BL/6 and BALB/c mice, although not reaching statistical significance because of the large variations among individuals. No IgG1 antibodies were produced in C57BL/6 mice, as reported. 15 By contrast, in BALB/c mice, a trend consisting of an 80% reduction of IgG1 (Fig. 2H: p = 0.10, one-way ANOVA) was observed as compared with the control groups. Interestingly, preimmunization with the loss-of-function peptide actually increased by 2.4-fold the concentration of anti-Ad5 IgG2a (Fig. 2E) and by 3.8-fold the concentration of IgG2b (Fig. 2G: p < 0.05, Mann–Whitney U test).
It is important to stress that the assay system used here was designed so as to evaluate the production of antibodies in response to the ensemble of proteins constituting the vector, indicating that the use of a single epitope of a single protein was enough to reduce the overall production of antibodies against the vector.
Immunization with a hexon 6-specific MHC class II peptide containing a thioredox motif induces sustained suppression of antibody production even on readministration of the viral vector
One of the key issues in gene therapy is to determine whether or not a vector can be readministered over time, that is, if methods used to suppress antibody production are efficient enough to prevent responses boosted by renewed exposure to the vector. Researchers have tried to circumvent this by making use of vectors in which, for instance, capsid proteins are replaced with those of an alternative adenovirus serotype. 16 This interesting approach is, however, limited by the small number of such alternative serotypes and by the fact that they are immunogenic on their own.
We therefore went on to determine whether the prevention of specific antibody production, and in particular that of the IgG2a isotype, was sustained over time. In BALB/c mice, Ad5 vectors administered three times led to detectable levels of all antibody isotypes; Ad5-specific antibody production was monitored in this strain (Supplementary Fig. S2A). The kinetics of total IgG, IgG2a, IgG2b, and IgG1 production over time are shown in Supplementary Fig. S2B–E, respectively.
Total IgG to Ad5 vector peaked on day 88 (open histogram; Supplementary Fig. S2B). The concentrations of IgG2a and IgG2b antibodies peaked, however, on day 57 (open histograms; Supplementary Fig. S2C and D), suggesting an alternative mechanism of antibody production (see below).
In mice immunized with the ccHexBALB/c peptide, a significant increase in antibodies was observed on day 88 (Supplementary Fig. S2B), seemingly due to an increase in IgG1 (Supplementary Fig. S2E). Strikingly, however, the IgG2a response was suppressed over the entire observation period (Supplementary Fig. S2C: p < 0.01, Mann–Whitney U test), indicating effective, sustained suppression over time.
Production of anti-Ad5 vector antibodies is dependent on innate immunity activation
Data presented in Supplementary Fig. S2B–E suggest earlier kinetics in the production of IgG2a as compared with IgG1 antibodies. The production of the IgG2a (BALB/c) or IgG2c isotype (C57BL/6) is IFN-γ dependent, whereas that of IgG1 is IL-4 dependent. 17 We therefore hypothesized that the source of IFN-γ could be from early activation of innate immunity. Ad5 vectors are indeed known to be strong activators of innate immunity, 18 and challenges with a large dose of Ad5 vector trigger a cytokine storm leading to shock syndrome and fatalities. 19 One obvious source candidate for early IFN-γ production is the NK/NKT cell. 20 NKT cells, in contrast to NK cells, are abundant in the liver, 21 which is the main organ affected by intravenous injection of Ad5 vector.
To investigate the possible role of NKT cells in the response against Ad5 vector administration, BALB/c mice and H2d -restricted CD1d KO mice (BALB/c mouse background with no NKT cells) were treated according to the same protocol (Fig. 3A). Briefly, mice were distributed into three groups (n = 6 per group): BALB/c mice were immunized with either ccHexBALB/c peptide or alum only (WT-BALC/c Ctrl) and CD1d KO mice received alum only (CD1d KO Ctrl). Then, all mice received two injections of Ad5 vector. Figure 3B–E shows that CD1d KO mice did not mount significant antibody responses to Ad5 as compared with BALB/c mice treated with alum only (a reduction of 91 and 98% for total IgG and IgG2a, p < 0.001, and 91% for IgG1, p < 0.05, one-way ANOVA), respectively. Interestingly, IgG2a antibodies were undetectable in CD1d KO mice.
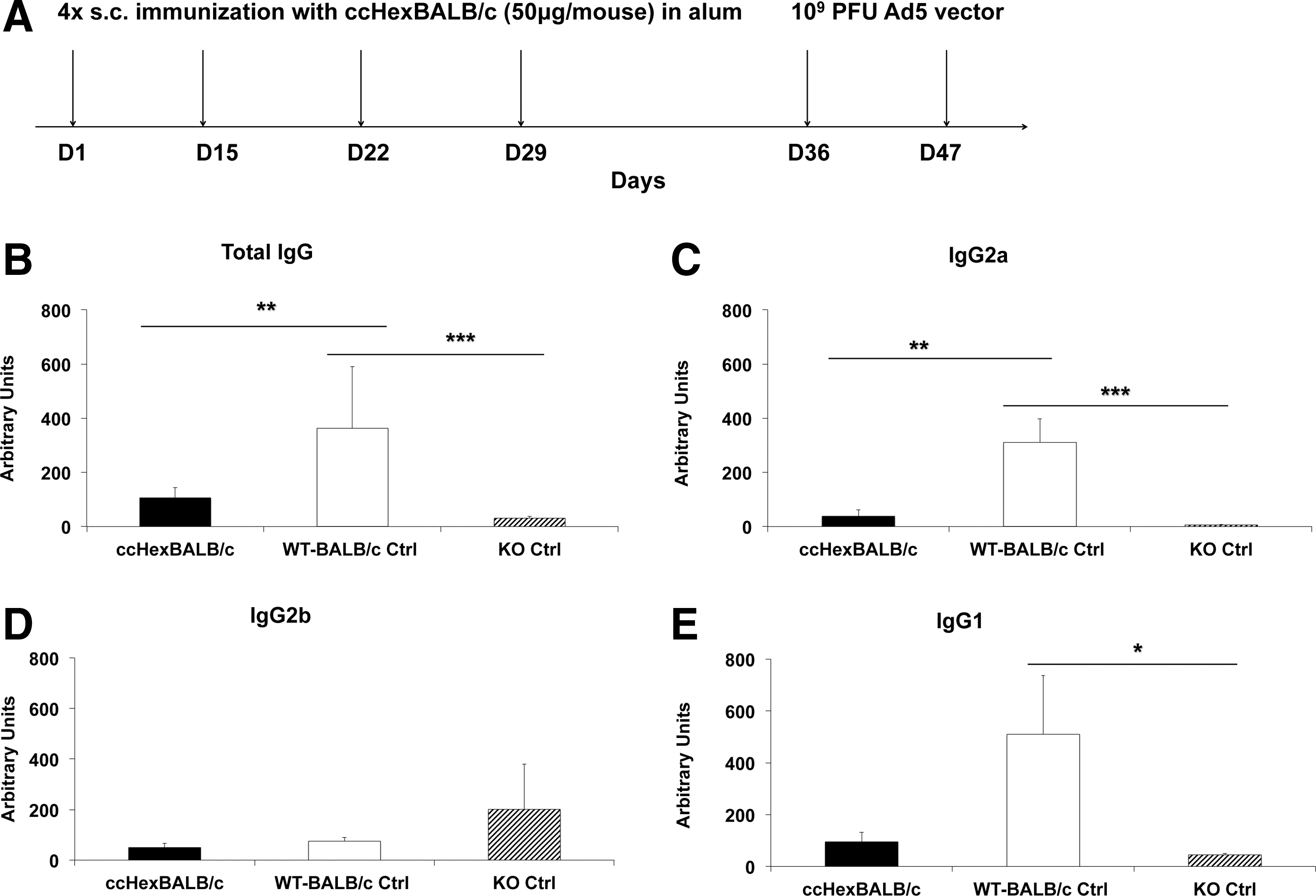
Immunization with ccHexBALB/c peptide prevents antibody production by altering natural killer T (NKT) cell activation.
Overall, the concentration of antibodies produced in CD1d KO mice paralleled closely that of ccHexBALB/c peptide-immunized wild-type BALB/c mice compared with the alum-immunized control group (a reduction of 71 and 90% for total IgG and IgG2a, one-way ANOVA, p < 0.01; Fig. 3B and C, respectively).
Immunization with an Ad5-specific MHC class II peptide containing a thioredox motif specifically prevents early innate immunity activation on Ad5 vector administration
To explore the involvement of innate immunity against Ad5 vectors, we first established that injection of 109 PFU of Ad5 vector elicited such innate immunity activation in the present experimental setting. In Fig. 4A–C (Ctrl group), control mice receiving Ad5 vector showed a brisk increase in the concentrations of IFN-α, IFN-γ, and of IL-6 occurring within 6 hr of vector administration. In mice preimmunized with ccHexBALB/c peptide (ccHexBALB/c group in Fig. 4A–C), a significant reduction in IFN-α (59%, p < 0.05) and IL-6 (49%, p < 0.05), and a complete prevention of IFN-γ production (≥90%, p < 0.01, Mann–Whitney U test), were observed, as compared with the control group.
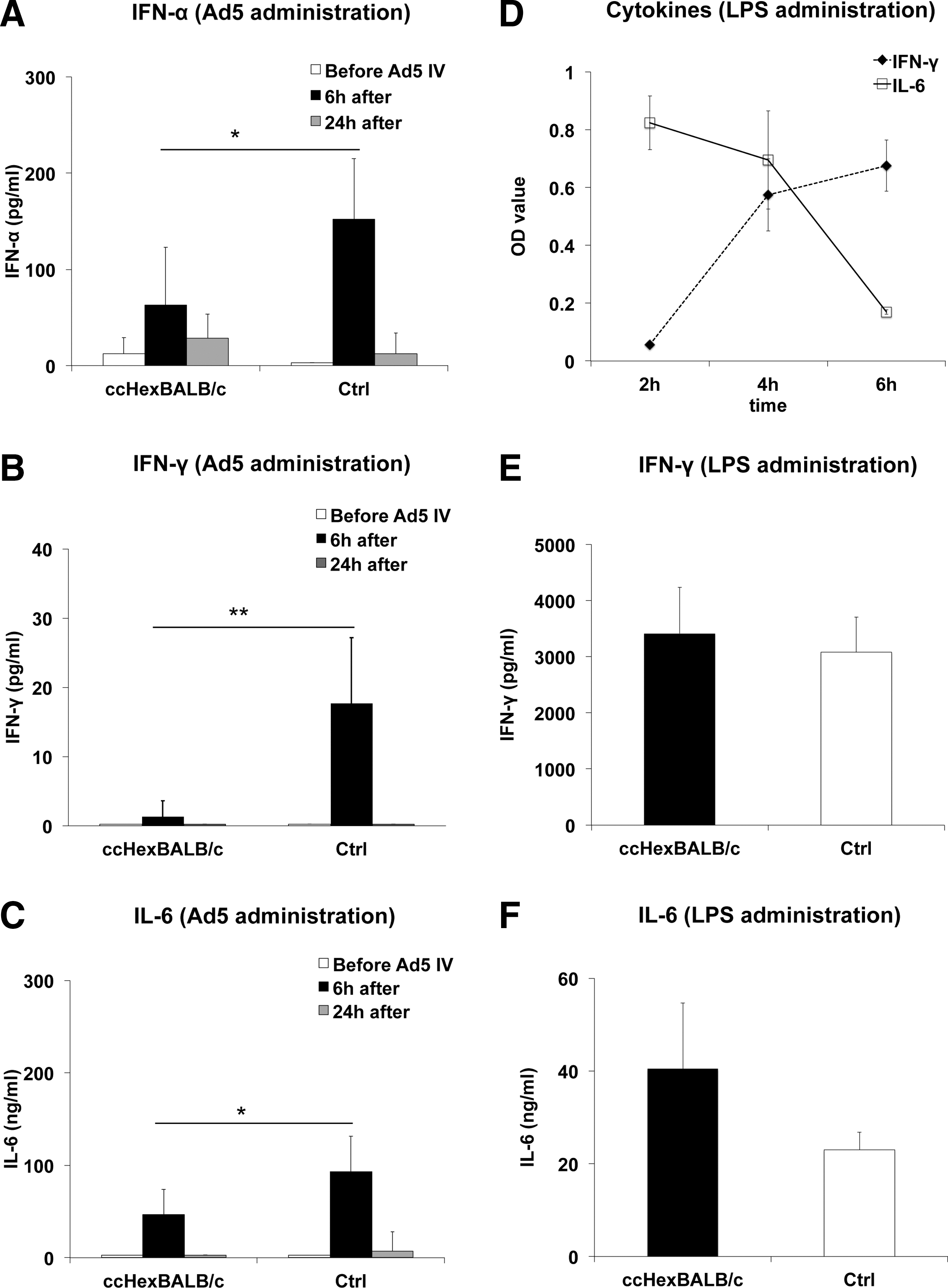
ccHexBALB/c peptide immunization prevents innate immune response after Ad5 injection into BALB/c mice.
This finding prompted us to check whether such an effect on cytokine production associated with early innate immunity activation was specific for Ad5 vector. To this end, we evaluated the production of IFN-α, IFN-γ, and IL-6 after intravenous injection of lipopolysaccharide (LPS), a canonical activator of innate immunity, in two groups of BALB/c mice: mice immunized with ccHexBALB/c or naive BALB/c mice. Although IFN-α was not detected, in keeping with reported data, 22 significant increases in IFN-γ and IL-6 were observed (Fig. 4D) in naive BALB/c mice, with peak time at 4 hr for the detection of both cytokines. Figure 4E and F indicates that immunization of mice with ccHexBALB/c peptide did not alter LPS-induced production of either IFN-γ or IL-6, establishing the antigen specificity of the innate immunity suppression shown previously.
Taken together, the data suggested that immunization with class II-restricted peptides from Ad5 vectors eliminates the early activation of innate immunity in a seemingly antigen-specific manner.
Immunization with hexon-specific MHC class II peptide containing thioredox motif prevents accumulation of NKT cells in the liver
The reduced antibody production in CD1d KO mice (Fig. 3B–E) and the prevention of IFN-γ production by immunization with ccHexBALB/c (Fig. 4B) reinforced the presumption that NKT cells could be central in these events. NKT cells are activated by (glyco)lipids presented in the context of the nonpolymorphic CD1d molecule. 23 As hepatocytes and Kupffer cells (KCs) express CD1d, we assumed that Ad5 vector injection would increase the expression of lipids by such cells, activating NKT cells, and thereby providing a source of IFN-γ production.
It is known that administration of (glyco)lipids activates NKT cells within minutes, followed by an accumulation of cells in the liver where interaction with Kupffer cells occurred. 24 –27 We administered Ad5 vector three times to alum-treated control mice and to a group of ccHexBALB/c-immunized mice (Fig. 5A). Two weeks after the last administration of Ad5 vector, mice were killed and livers were removed, and mononuclear cells (MNCs) were harvested and analyzed by FACS for the proportion of NKT and CD8+ cells in the liver. Liver cryosections were prepared to assess the percentage of KCs by immunohistochemistry. Figure 5B shows a significant increase in NKT cells (CD3+NKG2D+ cells) in control mice (2.6-fold) as compared with naive mice (one-way ANOVA, p < 0.01), whereas NKT cell accumulation was completely prevented in mice preimmunized with the ccHexBALB/c peptide. A small yet significant reduction was also observed in liver accumulation of CD8+ T cells (15%) and KCs (23%) in ccHexBALB/c-immunized mice (Fig. 5C and D: one-way ANOVA, p < 0.05).
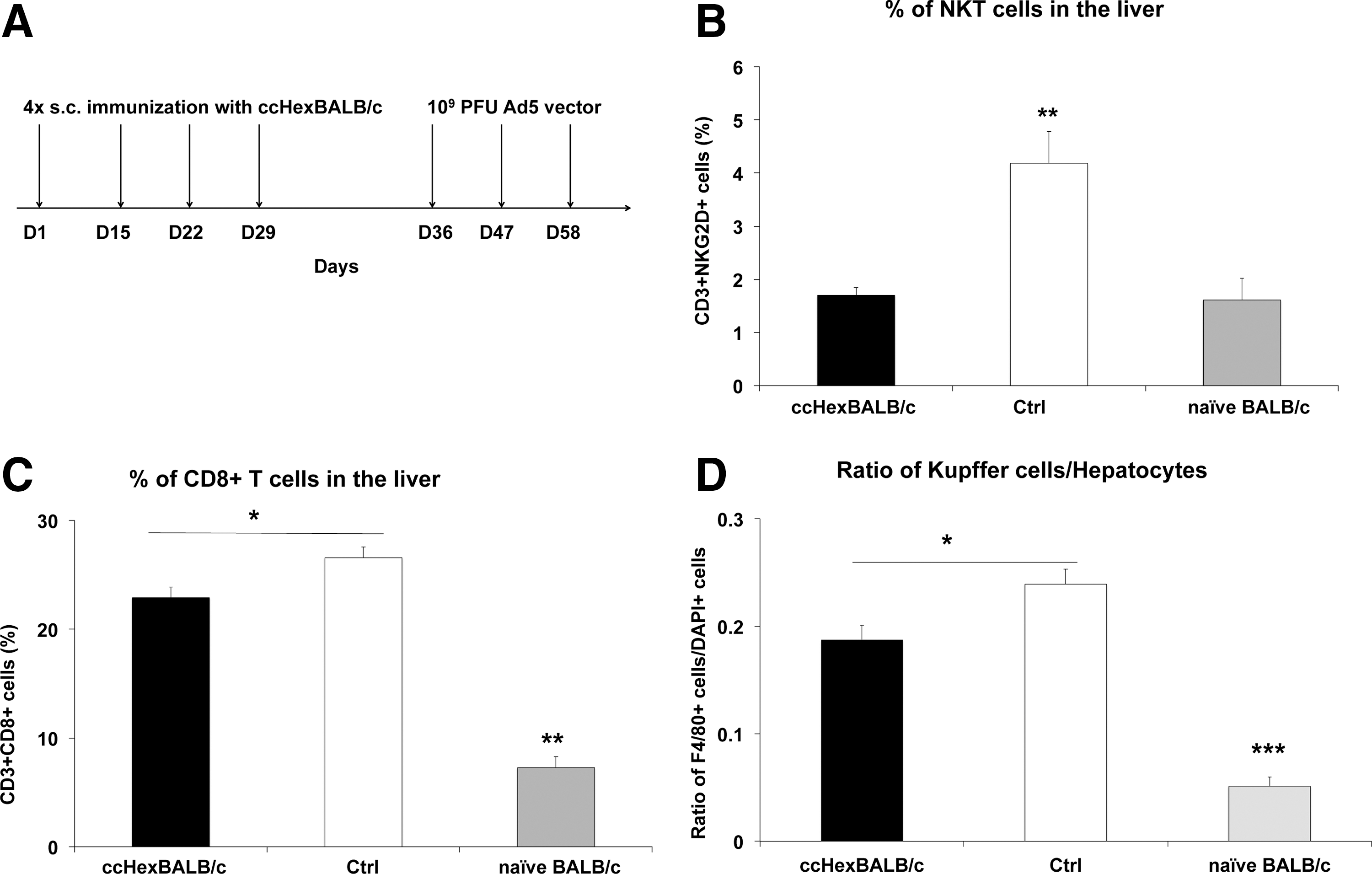
Immunization with the ccHexBALB/c peptide prevents accumulation of NKT, CD8+ T, and Kupffer cells on Ad5 vector administration.
It was therefore concluded that administration of such peptide significantly reduced the accumulation of inflammatory cells in the liver, and in particular NKT cells.
Mice immunized with thioredox motif-containing peptide show increased transgenesis and prolongation of transgene expression
We next questioned whether preventing both innate immunity activation as well as adaptive response to the Ad5 vector would be sufficient to increase the efficiency of transgenesis and to prolong transgene expression. It is known that the neutralizing antibody against Ad5 vectors is the determining factor preventing Ad5-mediated transgenesis. We therefore set up an experiment in which GFP was used as a reporter gene for assessing these two parameters. Because of the significant differences in the immune response to Ad5 observed in C57BL/6 and BALB/c mice (Fig. 2), we carried out these experiments in each of the two genetic backgrounds. However, considering that a loss-of-function peptide elicited increased antibody production (Fig. 2), the likelihood of having any effect on NKT cell accumulation in the liver was deemed to be remote, and therefore not tested in the present experiment.
Naive mice were challenged by a single administration of Ad5-GFP construct (109 PFU), and GFP expression in the liver was assessed on days 1, 7, and 42. On day 1, both strains showed significant GFP uptake, yet with 4-fold higher expression in C57BL/6 mice than BALB/c mice (Fig. 6A and D, day 1). As expected, GFP fluorescence gradually decreased over time and disappeared by day 42 in C57BL/6 mice (Fig. 6A, day 42) and by day 7 in BALB/c mice (Fig. 6D, day 7).

Thioredox motif-containing peptide immunization increases transgenesis and prolongs GFP expression in hepatocytes. Mice (n = 6 per group) were immunized four times with a thioredox motif-containing peptide [
We next immunized mice with thioredox motif-containing peptides before administration of the Ad5-GFP construct. An increase in transduction efficiency was not observed in C57BL/6 mice relative to the control group (Fig. 6A and B, day 1), both reaching maximal detection for the quantitation method, with 820 transduced cells/mm2 (Fig. 6C). In BALB/c mice, a 2.5-fold increase was observed in preimmunized mice (Fig. 6D and E, day 1, Mann–Whitney U test, p < 0.05), with 490 transduced cells/mm2 as compared with 210 cells/mm2 in the control group (Fig. 6F).
We then determined whether preimmunization could prolong expression of the GFP transgene over time. Figure 6C shows that, in C57BL/6 mice, the percentage of GFP+ cells remained at its initial value up to day 7, and then declined but maintained a residual value of 11% on day 42 (94 transduced cells/mm2). In BALB/c mice, thioredox motif-containing peptide immunization maintained 25% of GFP transgene expression, with 120 transduced cells/mm2 on day 7 (Fig. 6F).
Immunization with the thioredox motif-con-taining peptide therefore increased transgenesis at least in the BALB/c strain, and significantly prolonged GFP transgene expression in each of the two mouse strains (Mann–Whitney U test, p < 0.05).
Discussion
Main obstacles to the practice of gene therapy include the elicitation of an immune response to the viral vector and the difficulty in overcoming the presence of preexisting anti-vector antibodies. 28 It is thought that control of vector immunogenicity is essential for transduction efficiency and maintenance of transgene expression over long periods of time. We therefore explored the possibility of preventing such immunogenicity by using methodology that has been shown to be efficient in the control of alternative adaptive responses, such as in type 1 diabetes and multiple sclerosis. We show in the present paper that immunization with a peptide encompassing a class II-restricted peptide of the hexon-6 Ad5 capsid protein and a thioredox motif located within flanking residues results in significant suppression of the polyclonal antibody response elicited by Ad5 injection at doses envisioned for gene therapy.
Obviously, the traditional method involving nonspecific immunosuppressants would be impractical when gene vaccination is contemplated. However, vector-specific immunosuppression can greatly facilitate the efficacy of gene vaccination.
The methodology used in the present paper is antigen-specific, or more precisely T cell epitope-specific. Thanks to the acquisition of cytolytic properties toward antigen-presenting cells, the induced cCD4+ T cells keep under control immune responses even to complex antigens, provided the suppression activity is exerted at the surface of the antigen-presenting cells where an immune synapse is formed. Data on further characterization of the phenotype of elicited cCD4+ T cells can be found in a paper by Carlier and colleagues. 29
Worth mentioning, however, is the fact that CD4+ T cells as induced by our current technology constitute a separate subset as compared with regulatory T cells. cCD4+ T cells are best characterized as effector cells with activation of the phosphatidylinositol-3-kinase (PI3K)–Akt pathway, and mitogen-activated protein kinase (MAPK) and extracellular signal-regulated kinase (ERK) phosphorylation. They do express CD25 as do regulatory T cells (Tregs), but not Helios or neuropilin, the latter being characteristic of thymus-derived Tregs. At the transcriptome level, they show coexpression of Tbet and GATA3, yet not FoxP3. 29 Interestingly, however, we have observed that apoptotic bodies obtained by target cell lysis do generate Tregs as a consequence of presentation by immature dendritic cells. Overall, it is likely that the effects of cCD4+ T cells involve a downstream participation of Tregs (our unpublished data).
One of the consequences of such selective immune suppression is an increased efficacy of transgenesis. This was clearly observed in BALB/c mice, yet not detectable in C57BL/6 mice, likely because a maximal level of transgenesis was already obtained in naive mice with the number of PFUs used. However, in both strains, it was obvious that there was some degree of prolongation in transgene expression. We believe these data establish the proof of concept that control of the anti-vector response at the adaptive immunity level is sufficient to significantly modify the efficacy of gene therapy, extending the duration of transgene expression and allowing readministration of the vector. We are attempting to verify whether it is possible to obtain the same results in the presence of an ongoing response to adenoviral vector.
The implication of innate immunity in the elicitation of response to viral vectors is not new. 8,30 –34 However, whether this represents a separate, independent response to the vector, or whether this is an integral part of the overall vector immunogenicity, is not clear. The present experiments amply confirm the involvement of innate immunity, with peaks of IFN-α and IL-6 occurring early after intravenous administration of the vector.
We also show here that, in the absence of NKT cells, not only the early increase in cytokines is absent, but also the capacity to mount an adaptive response is significantly prevented. Such an adaptive response is thought to depend on the production of IL-21 and activation of follicular helper T cells. This aspect is under investigation, yet preliminary data indicate that NKT cells could constitute the main source of IL-21 in this setting, the elimination of which would prevent the activation of follicular helper T cells. NKT cells, at least as activated by α-galactosylceramide (invariant NKT cells), do produce copious amounts of IL-21. 35
There are some minor controversies about conservation of a full adaptive response in CD1d KO mice, 36 yet the extent of prevention observed in the present study indicates that NKT cell activation is essential to mount an adaptive response. This raises the intriguing possibility of controlling the overall immune response to viral vectors by acting at the NKT cell level. However, NKT cells are activated by lipids or glycolipids, which are nonspecific for a given response, thereby offering no direct possibility of specific prevention without affecting the overall capacity to activate such cells.
Taking into account the sequence of events leading from innate immunity activation to adaptive forms of response, it was unexpected to observe that the immunization procedure used here, strictly oriented toward class II-restricted adaptive immunity, profoundly affected innate immunity activation, both in terms of blunting early cytokine production and preventing accumulation of NKT cells in the liver, an observation associated with activation of such cells. The very fact that NKT cells still reacted to unrelated stimuli such as LPS administration discards a nonspecific effect of class II-restricted CD4+ T cells on innate immunity. The seemingly specific elimination of NKT cells in this setting points to direct contact between cCD4+ T cells and NKT cells. In fact, we have demonstrated (our unpublished data) that cCD4+ T cells can eliminate NKT cells if direct cell–cell contact can be established, such as at the surface of an antigen-presenting cell. Kupffer cells or macrophages within liver sinusoids are likely candidates for fulfilling the conditions for such an effect, yet this has still to be formally demonstrated. Worth noting, however, is our establishment that cCD4+ T cells, class II restricted, could indeed induce apoptosis of activated bystander CD4+ T cells 13 by a cell–cell contact-dependent mechanism mediated by the Fas–Fas ligand (FasL) interaction and granzyme B secretion. We therefore hypothesize that NKT cells, activated by presumed exposure to endogenous glycolipids in response to cellular stress resulting from viral transduction, are eliminated by CD4+ T cells activated at the surface of the same cells.
Whatever the precise mechanism linking these cells belonging to separate lineages, our findings, on the one hand, illustrate the sequential events by which an immune response to Ad5 vector is established and, on the other hand, provide the possibility to prevent the dual, innate, and adaptive response to Ad5 vectors by active immunization with a single class II-restricted peptide in a format eliciting acquisition of cytolytic properties by CD4+ T cells.
An important issue for translational application of the present technology is to determine whether it would be possible to suppress an existing immune response to Ad5, as a significant proportion of people are already immunized by natural exposure to the virus. This issue is not settled; yet, in several alternative applications of the technology (our unpublished data), such as experimental autoimmune encephalomyelitis (a model of multiple sclerosis), type 1 diabetes, and Dby-dependent skin transplant rejection, we showed that ongoing disease could be suppressed by direct vaccination with peptides containing a relevant autoantigen-derived class II-restricted T cell epitope and a thioreductase motif. On the basis of these results, we expect that vaccination of individuals already immunized to Ad5 would lead to suppression of the immune response to proteins contained in the viral vector.
It is our intention to apply the present immunization methodology to alternative viral vectors, and in particular to adeno-associated viral (AAV) vectors, often a first choice for clinical application. Considering the high and fast immune reaction to Ad5 vectors, including innate immunity, the situation with AAV, which contains far fewer viral proteins, is, however, predicted to be easier to control. In fact, AAV elicits less innate reactivity and less class II-restricted adaptive responses. However, it is well described that AAV vector-specific CD8+ T cell elicitation leads to lysis of transduced hepatocytes. 37,38 Current experiments are therefore examining to what extent the elicitation of cCD4+ T cells could indirectly control activation of CD8+ T cells, by virtue of deprivation of “help” provided by effector CD4+ T cells.
Footnotes
Acknowledgments
This work was supported by both the University of Leuven and ImCyse SA. The authors thank Dr. Evelien Gebruers for support in statistics.
Author Disclosure
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.