
Abstract
Duchenne muscular dystrophy (DMD) is a devastating muscle disease caused by loss-of-function mutations in DMD encoding dystrophin. No rational therapy is currently available. Utrophin is a paralog of dystrophin and is highly expressed at the neuromuscular junction. In mdx mice, utrophin is naturally upregulated throughout the muscle fibers, which mitigates muscular dystrophy. Protein-anchoring therapy was previously reported, in which a recombinant extracellular matrix (ECM) protein is delivered to and anchored to a specific target using its proprietary binding domains. Being prompted by a report that intramuscular and intraperitoneal injection of an ECM protein, biglycan, upregulates expression of utrophin and ameliorates muscle pathology in mdx mice, protein-anchoring therapy was applied to mdx mice. Recombinant adeno-associated virus serotype 8 (rAAV8) carrying hBGN encoding human biglycan was intravenously injected into 5-week-old mdx mice. The rAAV8-hBGN treatment improved motor deficits and decreased plasma creatine kinase activities. In muscle sections of treated mice, the number of central myonuclei and the distribution of myofiber sizes were improved. The treated mice increased gene expressions of utrophin and β1-syntrophin, as well as protein expressions of biglycan, utrophin, γ-sarcoglycan, dystrobrevin, and α1-syntrophin. The expression of hBGN in the skeletal muscle of the treated mice was 1.34-fold higher than that of the native mouse Bgn (mBgn). The low transduction efficiency and improved motor functions suggest that biglycan expressed in a small number of muscle fibers was likely to have been secreted and anchored to the cell surface throughout the whole muscular fibers. It is proposed that the protein-anchoring strategy can be applied not only to deficiency of an ECM protein as previously reported, but also to augmentation of a naturally induced ECM protein.
Introduction
D
The extracellular matrix (ECM) protein, biglycan, is a leucine-rich proteoglycan and binds to α-dystroglycan, α-sarcoglycan, and γ-sarcoglycan. 5,6 Biglycan is encoded by BGN located on the X chromosome in both rodents and human. In muscle cells, biglycan upregulates the sarcolemmal localization of dystrobrevin, syntrophin, and nNOS. 7 Biglycan-deficient (Bgn –/0) mice display decreased growth rates as well as reduced bone mass, which is first recognized at 6 months, and premature osteoarthritis. 8 Expression of utrophin is also reduced. 9 Recombinant human biglycan protein (rhBGN) injected into the muscle of Bgn –/0 mice becomes stably associated with the sarcolemma and ECM for at least 2 weeks, and upregulates the expression of DAPC component proteins. 7 Similarly, rhBGN injected into the peritoneum of mdx mice upregulates expression of utrophin and other DAPC component proteins on the sarcolemma, and ameliorates muscle pathology as well as motor deficits. 9 However, the injected rhBGN stays on the sarcolemma for only 3 weeks.
Protein-anchoring therapy was previously reported, in which a recombinant ECM protein is delivered to and anchored to a specific target using the proprietary binding domains of the ECM protein. 10 Collagen Q (ColQ) is an ECM protein that makes a triple helix and binds to 12 catalytic subunits of acetylcholinesterase (AChE) to anchor AChE to the synaptic basal lamina at the NMJ. A single intravenous injection of recombinant adeno-associated virus serotype 8 (rAAV8) carrying human COLQ into the tail vein of Colq-knockout mice enabled specific anchoring of the AChE/ColQ protein complex to the NMJ, and almost normalized motor functions by mitigating abnormalities in the NMJ ultrastructure and the electrical signal transmission at the NMJ. In order to validate further the proof-of-concept that the protein-anchoring therapy is applicable not only to a defect of the ECM protein but also to the augmentation of a naturally induced ECM protein, rhBGN was expressed to treat mdx mice. The study shows that a single intravenous injection of rAAV8-hBGN carrying human BGN cDNA to mdx mice upregulates the expression of utrophin in the long-term and markedly improves motor deficits and dystrophic muscle pathologies.
Materials and Methods
Preparation of AAV carrying human BGN
To make rAAV8-hBGN, HEK-293EB cells were co-transfected with pAAV8, pHelper, and pAAV-hBGN, which carried human BGN downstream of the CMV promoter, using the calcium phosphate co-precipitation method. rAAV8-hBGN was purified by cesium chloride gradient ultracentrifugation as previously described. 10 The viral titer was estimated by quantitative polymerase chain reaction (PCR) using LightCycler 480 II (Roche).
Animals and AAV administration
All mouse studies were approved by the Animal Care and Use Committee of the Nagoya University Graduate School of Medicine. The genetic background of mdx mice was C57BL/10, and the strain of wild-type mice was matched to male C57BL/10. Multiple sets of littermates were used for both wild-type and mdx mice in order to increase the number of mice to be analyzed. Each mouse was housed individually in a cage on a 12-h light/dark cycle at 25°C. rAAV8-hBGN (1 × 1012 vg) was injected into the tail vein of 5-week-old male C57BL/10 mdx mice. All mice were examined for motor activities up to 24 weeks of age and were then sacrificed. Sacrificed mice were divided into batches for extracting DNA, RNA, or protein, or for analyzing skeletal muscle morphologies.
Motor activity tests
Muscle strength and fatigability were measured with the running wheel, rotarod, and grip strength tests every 2 weeks from 4 to 24 weeks of age. Running-wheel tests and rotarod tests were for quantifying voluntary and forced exercises, respectively.
For the running-wheel test, each mouse was housed in an individual counter-equipped running-wheel cage (Ohara Medical). The running distance was recorded over 72 h, and the average distance in 24 h was calculated and analyzed.
For the rotarod test, mice were examined on a rotarod apparatus (Ugo Basile) every 2 weeks. Mice were trained on the rotarod for 2 days before the initial measurement. Three rotarod protocols were used: accel, fix, and step. With the accel protocol, the rod was linearly accelerated from 5 to 45 rpm over 500 s. 11 With the fix protocol, the rod was gradually brought from 0 rpm to a constant speed of 30 rpm, and the mouse was monitored up to 300 s. 12 With the step protocol, the rotation was accelerated in four consecutive steps: (1) 5 rpm for 120 s, (2) 10 rpm for 60 s, (3) acceleration up to 40 rpm over 30 s, and (4) 40 rpm for 150 s. 13 The first 5-rpm step was for acclimatization of the mouse and was not included in the dwell time on the rod. Each mouse was run on each protocol three times with a 10-min rest between each trial, and the dwell times on the rod were averaged. The rotarod test with the step protocol was performed on the following day.
The grip strengths of fore-, hind-, and all limbs were measured using the DFE II Digital Force Gauge (Chatillon, Ametek, Inc.). 13 Mice were acclimatized to grasp meshes with fore-, hind-, and all limbs for two consecutive days before data collection. To measure the strength of the fore- and all limbs, mice were allowed to hold the horizontal mesh with the fore- and all paws, and then the tail was gently pulled back until the grip was broken. Each measurement was repeated three times within 2 min. To measure the strength of the hindlimbs, an angled mesh assembly was used. The mouse's tail was pulled toward the meter and also parallel to the mesh assembly. The average of three scores was calculated. The grip strengths were measured by a single researcher.
Morphological studies
Mice were sacrificed at 24 weeks of age. Quadriceps muscles were isolated and flash-frozen in liquid nitrogen–cooled isopentane, and sectioned at 8 μm thick with a cryostat (CM3050; Leica) at –20°C. The frozen quadriceps sections were stained with hematoxylin and eosin. Sections were dehydrated with ethanol three times, and were permeated with Lemosol (Wako) and were enclosed with Softmount (Wako). The frozen sections of gastrocnemius and triceps brachii were used for immunofluorescence staining. The frozen sections were fixed by acetone at 4°C for 10 min, blocked with 5% horse serum in phosphate-buffered saline for 1 h, and incubated with the primary antibody overnight. The primary antibodies were utrophin (1:100; BD Biosciences), α1-syntrophin (1:1000; GeneTex), γ-sarcoglycan (1:100; Novusbio), or α-dystrobrevin (1:150; Abcam). Sections were incubated with the secondary antibody of anti-goat-FITC (1:100; Vector), anti-rabbit-FITC (1:100; Vector), anti-rat-FITC (1:100; Open Biosystems), or anti-mouse-FITC (1:100; Vector) for 1 h. If the host of the primary antibody was mice, a M.O.M Immunodetection Kit (Vector) was used according to the manufacturer's protocols. Signal intensities of utrophin, γ-sarcoglycan, α-dystrobrevin, and α1-syntrophin were analyzed by the MetaMorph imaging software (Molecular Devices). After immunostaining plasma membrane with laminin α2 antibody (1:100; Enzo), sizes of 300–500 myofibers in 10 non-overlapping fields in each mouse were estimated using the NIS-elements AR (Nikon). Morphometric analyses were blindly performed using three or four mice in each group.
Plasma creatine kinase
Mice were deeply anesthetized with isoflurane, and blood samples were obtained by cardiac puncture. Blood was collected in a 1.5 mL heparinized tube, and centrifuged at 1200 g for 10 min. The activity of creatine kinase (CK) was assayed by the standard method by Oriental Yeast Co. Ltd.
Real-time PCR
Gastrocnemius muscles and the liver were isolated from 24-week-old male mice, and were incubated at 4°C overnight in RNAlater solution (Ambion). Total RNAs from the gastrocnemius muscles and the liver were extracted using the RNeasy Mini Kit (Qiagen) with DNase I- and proteinase K-treatment according to the manufacturer's protocols. Purified RNA was converted to cDNA using the ReverTra Ace reverse transcriptase (Toyobo). Quantitative reverse transcription real-time PCR (qRT-PCR) reactions were performed using the SYBR Premix Ex Taq II (Takara Bio., Inc.) on the LightCycler 480 II (Roche). Gapdh was also quantified for normalization. Primer sequences are shown in Supplementary Table S1 (Supplementary Data are available online at
Western blotting
Total protein was extracted from the gastrocnemius muscle by homogenization in RIPA buffer (Thermo Scientific) with the cOmplete Protease Inhibitor Cocktail (Roche). Homogenates were centrifuged at 10,000 g for 15 min, and supernatants were collected. The first and second antibodies were anti-biglycan antibody (1:1000; Abcam) and anti-rabbit-horseradish peroxidase (1:2000; GE Healthcare), respectively. GAPDH in a loading control was detected by anti-GAPDH HRP-DirecT (MBL).
Transduction efficiency
To quantify the transduction efficiency, total DNA was extracted from gastrocnemius muscles and the liver using the QIAamp DNA Mini Kit (Qiagen). The primers were designed on the CMV promoter, and qPCR reactions were performed using the SYBR Premix Ex Taq II (Takara) on the LightCycler 480 II (Roche). A gene for telomerase reverse transcriptase (Tert) was also quantified for normalization. Primer sequences are shown in Supplementary Table S1.
Statistical analysis
Statistical analysis was performed with Prism v6.0f (GraphPad). All data were analyzed using one-way analysis of variance with the Tukey multiple comparison test.
Results
A single intravenous injection of rAAV8-hBGN improves motor functions in mdx mice
In this study, 1 × 1012 vector genomes (vg) of rAAV8-hBGN was intravenously injected into 5-week-old dystrophin-deficient mdx mice (AAV-mdx mice). The voluntary exercise was estimated using a counter-equipped running wheel every 2 weeks from 4 to 24 weeks of age. The spontaneous movement of AAV-mdx mice barely exceeded that of mdx mice after 8 weeks of age (Fig. 1A). Grip strengths of fore-, hind-, and all limbs of AAV-mdx mice were 10.3–28.6% higher than those of mdx mice throughout a range of ages from 6 to 24 weeks (Fig. 1B–D). Balance and cooperative ambulatory exercise was tested with the accel, fix, and step protocols of the rotarod apparatus. With all the protocols, the dwell time on the rod was much better than that of mdx mice after 8 weeks of age (Fig. 1E–G).
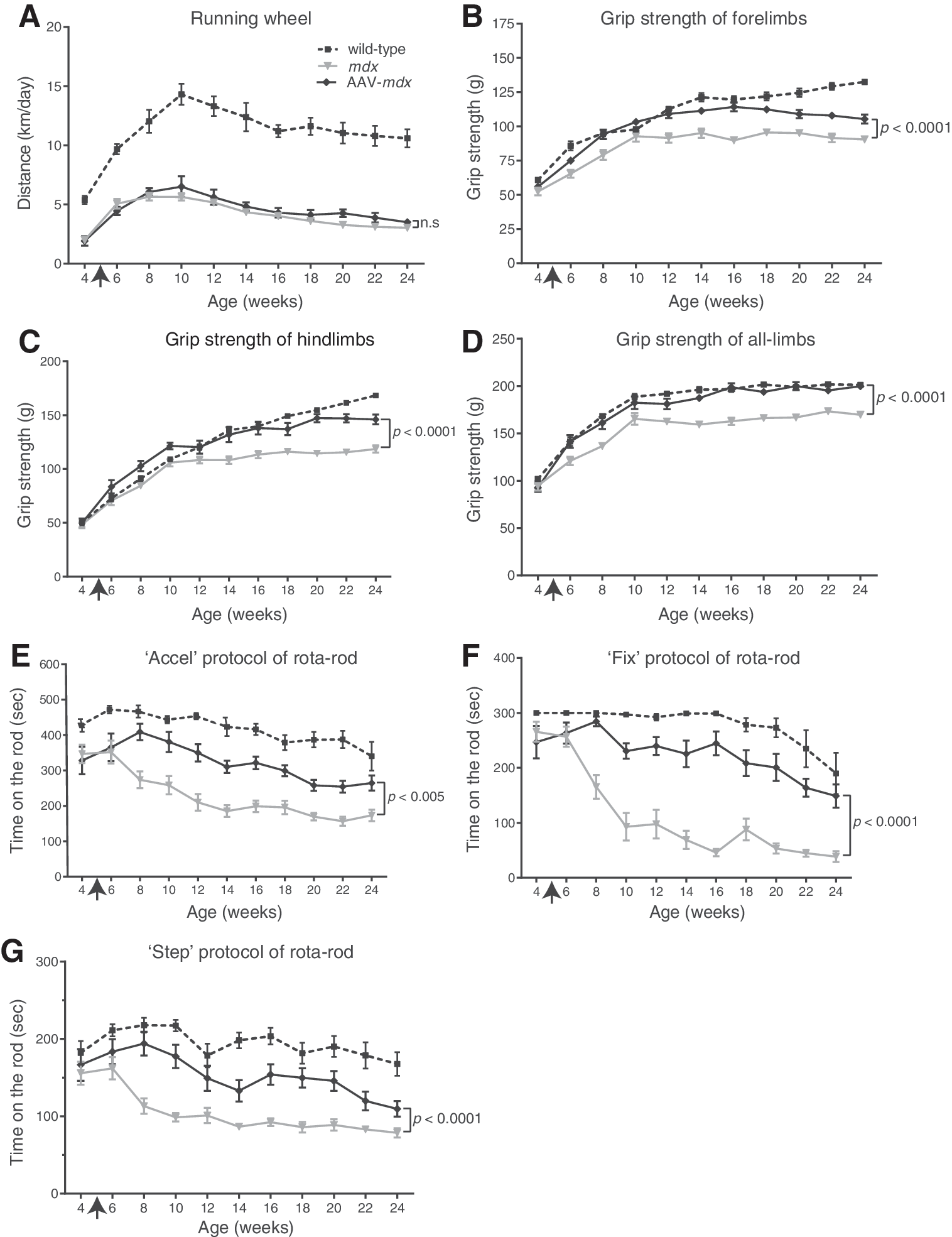
Motor functions of wild-type, untreated mdx, and treated mdx (AAV-mdx) mice. AAV-mdx mice had a single intravenous injection of rAAV8-hBGN (1 × 1012 vg) at 5 weeks of age (arrow). Each group is comprised of 10–16 mice, and the mean and standard error (SE) are plotted. Statistical differences are calculated with two-way repeated measures analyses of variance (ANOVA). n.s., not significant. (
Central nuclei are reduced in AAV-mdx mice
Pathological features of quadriceps muscles were analyzed. Muscle fibers that have been injured, repaired, and regenerated contain centrally located myonuclei, which serve as a persistent morphological indicator for muscle degeneration and regeneration. 14 The effect of rAAV8-hBGN on muscle fiber regeneration was examined by measuring the proportion of muscle fibers containing central nuclei in wild-type, mdx, and AAV-mdx mice at 24 weeks of age. The ratios of central nuclei-positive fibers in wild-type, mdx, and AAV-mdx were 2.1%, 53.8%, and 28.3% (Fig. 2A and B). The treatment thus markedly reduced the number of central nuclei.
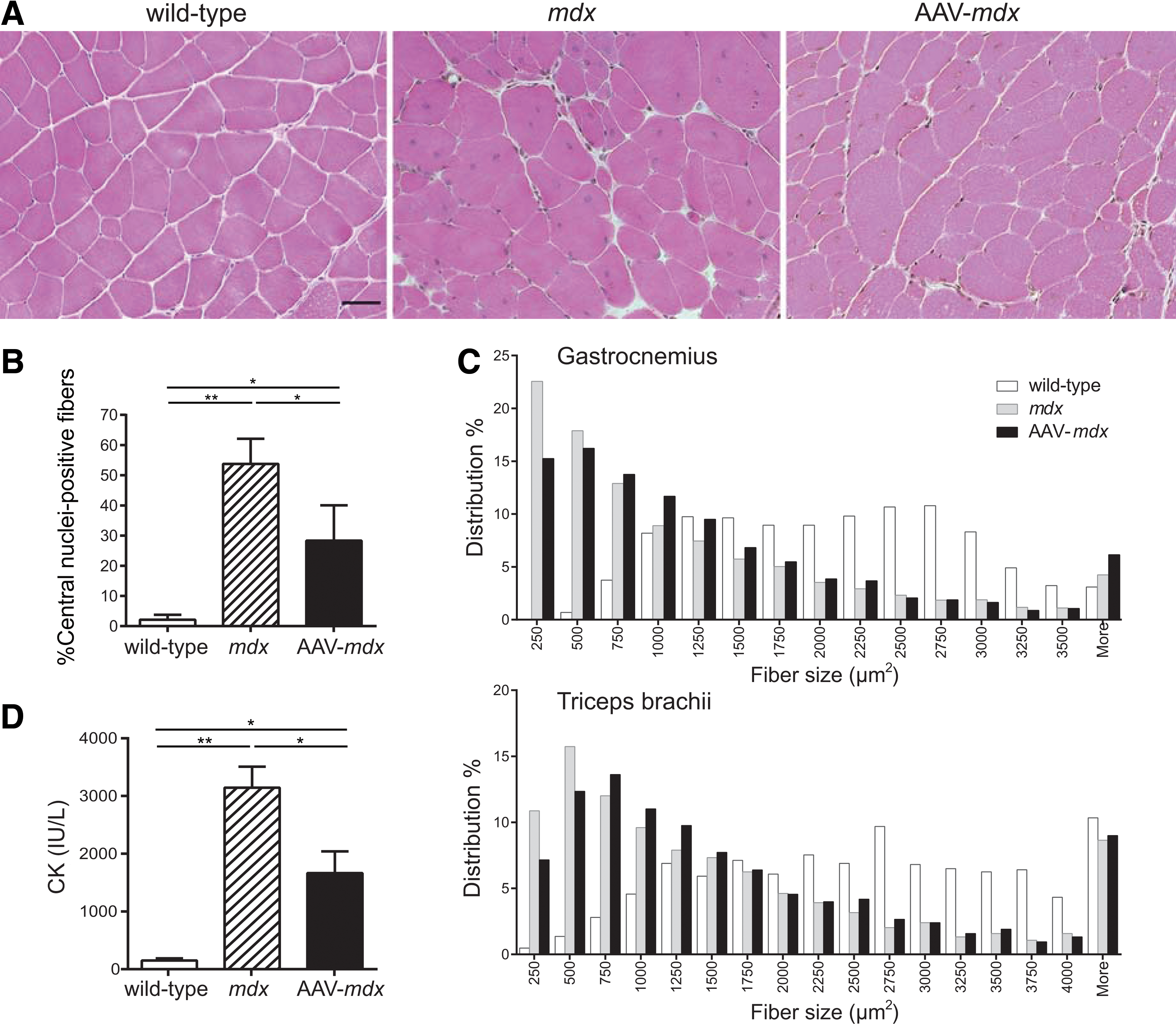
Intravenous administration of rAAV8-hBGN reduces central nuclei in muscle fibers in mdx mice. (
Myofiber sizes are improved in AAV-mdx mice
The myofiber cross-sectional area was measured in gastrocnemius and triceps brachii. In AAV-mdx mice, the percentage of myofibers ≤500 μm2 in size were decreased and that of myofibers >500 μm2 in size were increased compared with untreated mdx mice, rendering a shift of the distribution of myofiber sizes toward larger values (Fig. 2C).
Plasma CK activity decreases in AAV-mdx mice
Untreated mdx mice had 20.6-fold higher plasma CK activities compared with wild-type mice at 24 weeks of age (Fig. 2D). rAAV8-hBGN treatment decreased the plasma CK activities to 10.9-fold of wild-type mice, suggesting that rAAV8-hBGN ameliorated sarcolemmal vulnerability of mdx mice.
Utrophin and DAPC component proteins are upregulated in AAV-mdx mice
Next, we examined whether rAAV8-hBGN upregulates utrophin in mdx mice. Immunostaining of gastrocnemius muscles showed that the sarcolemmal expression of utrophin was increased in mdx mice, and that rAAV8-hBGN treatment further increased the sarcolemmal expression of utrophin (Fig. 3A and B).
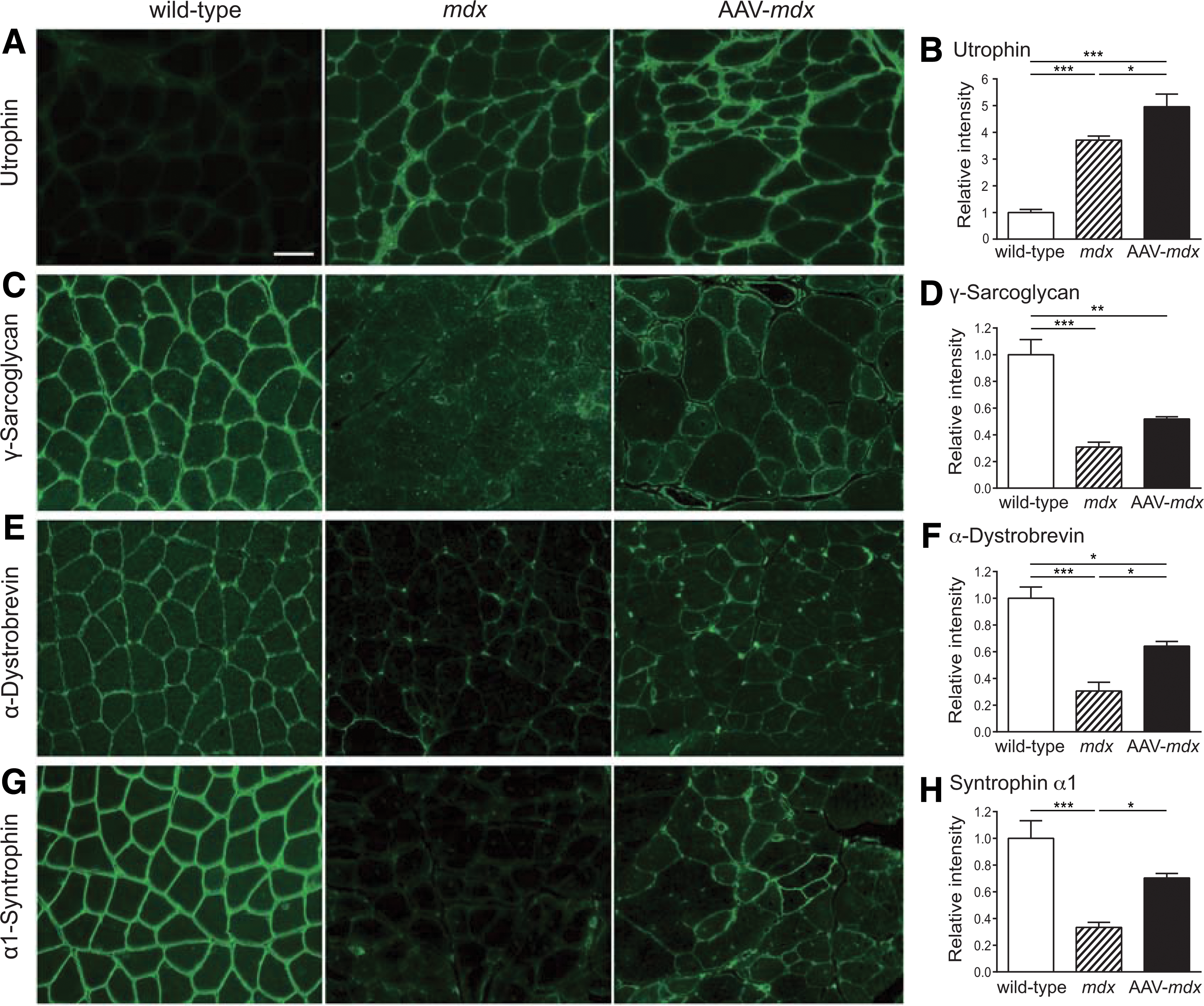
Histological analyses of gastrocnemius muscles in each group of mice. Gastrocnemius muscle sections of 24-week-old wild-type, mdx, and AAV-mdx mice are stained with utrophin (
The expressions of DAPC component proteins were compared in wild-type, mdx, and AAV-mdx mice. Immunostaining of the gastrocnemius muscle revealed that the sarcolemmal expressions of γ-sarcoglycan, α-dystrobrevin, and α1-syntrophin were lower in mdx mice compared with those in wild-type mice (Fig. 3C–H). However, the sarcolemmal expressions of these proteins in AAV-mdx mice were upregulated. Moreover, expressions of these proteins were continuous and uninterrupted on the sarcolemma in AAV-mdx mice. These results indicate that rAAV8-hBGN markedly ameliorated defective expressions of DAPC component proteins in mdx mice.
mRNA levels for utrophin and β1-syntrophin are upregulated in AAV-mdx mice
Next, we examined whether expressions of DAPC component proteins were upregulated in AAV-mdx mice. qRT-PCR analysis revealed that the treatment upregulated the gene expressions of Utrn (utrophin), and Sntb1 (β1-syntrophin) 1.32- and 1.14-fold, respectively (Supplementary Fig. S1). In contrast, the expression of Nos1 (nNOS) was downregulated, and the expressions of Dtna (α-dystrobrevin) and Musk (muscle-specific receptor tyrosine kinase, MuSK) remained unchanged.
rAAV8-hBGN induces expression of human BGN preferentially in the liver rather than in skeletal muscle
Next gene expression of the human recombinant biglycan (hBGN) delivered by rAAV8-hBGN in skeletal muscle and the liver was examined. As human BGN cDNA (hBGN) and mouse Bgn cDNA (mBgn) are identical at 981/1110 (88%) nucleotides, we failed to design PCR primers or TaqMan probes that specifically amplify hBGN expressed by rAAV8-hBGN. Thus, the total expression level of hBGN + mBgn transcripts was quantified by placing PCR primers on segments shared by hBGN and mBgn, and the expression level of mBgn transcript was subtracted. mBgn was specifically amplified by placing a forward primer on the 5′ UTR of mBgn.
The total expression levels of hBGN and mBgn were quantified in the gastrocnemius muscle of wild-type, mdx, and AAV-mdx mice. In mdx mice, the expression of the biglycan gene was 5.85-fold higher than that in wild-type mice (Fig. 4A). In AAV-mdx mice, the expression of the biglycan gene was 7.85-fold higher than that in wild-type mice. As the expression level of mBgn was not changed in AAV-mdx mice, rAAV8-hBGN increased the expression of the biglycan gene 1.34-fold (7.85/5.85).
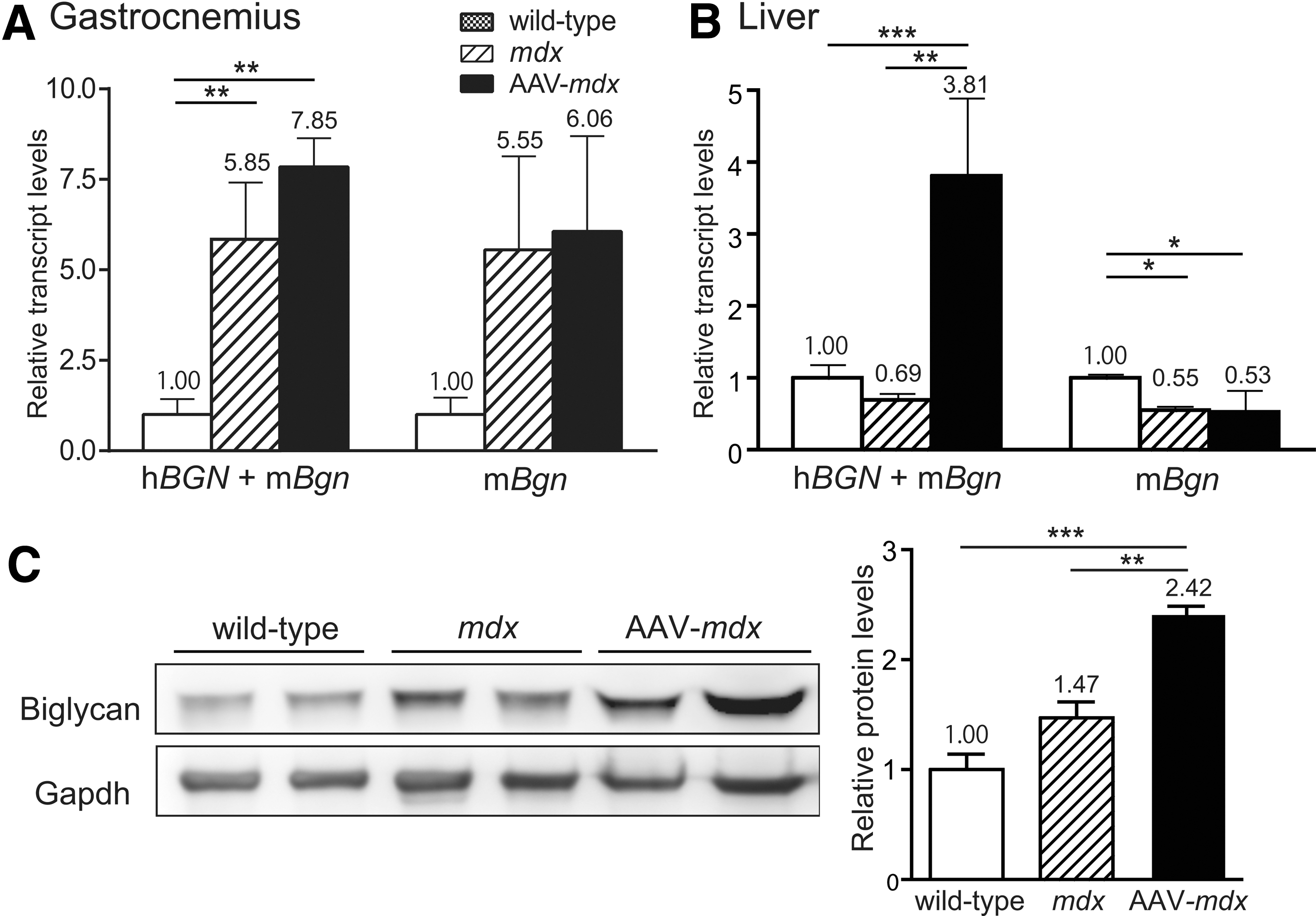
Expression of total biglycan (recombinant human biglyan and native mouse biglycan) is quantified at the gene and protein levels. Expression levels of hBGN + mBgn mRNA and mBgn mRNA in the gastrocnemius muscles (
Because AAV8 has tropism for hepatocytes in addition to skeletal muscle, 15 next the expression of hBGN in the liver was quantified (Fig. 4B). In mdx mice, the expression of the biglycan gene in the liver was reduced to 0.69 of that in wild-type mice. Thus, unlike the skeletal muscle, the expression of biglycan was decreased in the liver in mdx mice. On the other hand, in AAV-mdx mice, the expression of the biglycan gene in the liver was 3.81-fold higher than that in wild-type mice. As the expression level of mBgn was not changed in AAV-mdx mice, rAAV8-hBGN increased the expression of the biglycan gene 5.49-fold (3.81/0.69) in the liver.
Next, the protein level of biglycan in the gastrocnemius muscle was examined in wild-type, mdx, and AAV-mdx mice (Fig. 4C). In mdx mice, the expression of biglycan was 1.47-fold higher than that in wild-type mice. In AAV-mdx mice, the expression of biglycan was 2.42-fold higher than that in wild-type mice. Thus, the expression of biglycan in AAV-mdx mice was 1.65-fold (1.47/2.42) higher than that in mdx mice.
As the expression of hBGN in the skeletal muscle was lower than expected, the transduction efficiency of rAAV8-hBGN in the gastrocnemius muscle and the liver was estimated. DNA was extracted, and the copy numbers of the CMV promoter included in rAAV8-hBGN were estimated by qPCR. The copy number of the telomerase reverse transcriptase gene (Tert) was also estimated for normalization. The copy number of rAAV8-hBGN was divided by the copy number of Tert twice to calculate the number of rAAV8-hBGN per cell. As a result, the number of rAAV8-hBGN per cell was 0.164 ± 0.119 (M ± SD; n = 4) in the skeletal muscle, whereas it was 11.6 ± 9.07 (M ± SD; n = 4) in the liver. Therefore, rAAV8-hBGN was more efficiently transduced into the liver rather than into the skeletal muscle.
Discussion
We treated mdx mice modeling for DMD by injecting rAAV8-hBGN carrying human BGN cDNA into the tail vein. An improvement in motor function, reduction in the number of central myonuclei and plasma CK activities, improvement in the distribution of myofiber sizes, and upregulation of the expressions of biglycan and DAPC component proteins were observed. In contrast to marginal improvements in voluntary exercise measured by the running wheel (Fig. 1A), forced exercises measured by the grip strength tests (Fig. 1B–D) and the rotarod tests (Fig. 1E–G) were markedly improved. At 4 weeks of age when examinations of motor functions were started, the voluntary exercise activities of mdx mice were about half of those of wild-type mice (Fig. 1A). In contrast, the forced exercise performances in mdx mice were comparable to those of wild-type mice (Fig. 1B–G). This may represent habitual preference of mdx mice, but the underling mechanisms remain unknown. Improvement in muscle strength by rAAV8-hBGN was not sufficient to change their habitual preference.
Intravenous injection of rAAV8-hBGN increased the expression of the biglycan gene in the gastrocnemius muscle only 1.34-fold (Fig. 4A). Indeed, qPCR of the vector genome revealed that only 16.4% of skeletal muscle nuclei were transduced with rAAV8-hBGN. However, rAAV8-hBGN increased the expression of the biglycan protein in the gastrocnemius muscle 1.65-fold, which was higher than predicted from the expression of the biglycan gene. In gene therapy for intracellular proteins, a low transduction efficiency and a minimal increase in gene expression generally exhibit no effect. 10,16 In contrast, ECM proteins carry proprietary binding domains that enable specific anchoring of the transgene product to the target organ. 17 We show that intramuscular injection of rAAV1-COLQ into the right hindlimb, as well as intramuscular injection of purified AChE/ColQ protein complex into the bilateral gluteus maximus, enabled hanging on the wire for ≥2 min. 10 The rAAV1-COLQ-injected and AChE/ColQ-injected mice indeed showed specific anchoring of AChE/ColQ at all the examined NMJs in the uninjected forelimbs. Low transduction efficiency of rAAV8-hBGN and marked improvement in motor function in mdx mice suggest that biglycan expressed in a small number of muscle fibers were released and anchored to the cell surfaces throughout the entire muscle fibers, and improved motor function as well as muscle pathology by upregulating the expression of DAPC proteins. It was also observed that the liver had a high transduction efficiency of rAAV8-hBGN (1163.6%), and mRNA expressions of hBGN + mBgn in the liver were increased 5.49-fold (Fig. 4B). In a previous study, 10 the liver was similarly highly transduced with rAAV8-COLQ. However, as the liver does not produce AChE, AChE/ColQ cannot be produced by the liver. As biglycan requires no binding partner, biglycan produced by the liver might have partly contributed to anchoring of biglycan on the cell surface of muscle fibers. Amenta et al. previously reported that intraperitoneal injection of recombinant human biglycan reduced muscle pathology in mdx mice, 9 which is in accordance with the notion that biglycan can be anchored to the target organ using its proprietary binding domains.
rAAV8-hBGN was intravenously injected only at 5 weeks of age, and the improvement in motor function lasted at least up to 24 weeks of age. The effect of the protein-anchoring therapy with a single intravenous injection of rAAV8-COLQ similarly lasted at least 48 weeks, and the treated Colq-knockout mice, which usually die before 6 months of age, survived up to 18–20 months. 10 More than 99.5% of the AAV vector genome stays episomal in mouse muscles, even at 57 weeks after injection, 18,19 and expression of the transgene in skeletal muscle lasts for 1–1.5 years without decline. 20,21 These studies again underscore the long-lasting expression of the transgene delivered by AAV.
The authors previously reported the marked effects of the protein-anchoring therapy by intravenous injection of rAAV8-COLQ to Colq-knockout mice. 10 In contrast to the protein-anchoring of ColQ, mdx mice are not deficient for biglycan and rather overexpress biglycan. The current studies support the notion that the protein-anchoring strategy can be applied not only to deficiency of an ECM protein, but also to augmentation of a naturally induced ECM protein.
Footnotes
Acknowledgments
We thank Dr. James M. Wilson at the University of Pennsylvania for providing the chimeric helper plasmid pRC8 (identical to p5E18-VD2/8), and Dr. Takashi Okada at Nippon Medical University for productive discussion on this project. We also acknowledge the Central Institute for Experimental Animals for providing C57BL/10 mdx mice. This work was supported by Grants-in-Aid from the MEXT, MHLW, and AMED of Japan.
Author Disclosure
All authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.