
Abstract
Sustained expression of the CFTR gene is a major challenge to gene therapy with either viral or nonviral vectors with immune response to vector and transgene products. One strategy to achieve sustained CFTR expression is to modulate the host immune system through transient immunosuppression. In this study, we examined cyclophosphamide (cytoxan), dexamethasone (Dex), and a combination of cyclosporin, methylprednisolone, and azathioprine (combination) for their effects on long-term expression of the human CFTR delivered with helper-dependent adenoviral vectors in mouse airways. We found that cyclophosphamide significantly enhanced long-term expression of the transgenic human CFTR and the reporter gene LacZ by reducing host immune responses. Dex administration greatly reduced neutralizing antibody production but had no effect on transgene expression. Treatment with a combination of cyclosporin A, azathioprine, and methylprednisolone affected neither CFTR gene expression nor inflammation. Our data suggest that transient immunosuppression might be a strategy to improve sustained expression in gene therapy.
Introduction
C
Lack of long-term CFTR gene expression is the major challenge to the development of CF gene therapy. The recent report of the U.K. gene therapy trial proves this point. 7 Adenoviral (Ad) vectors are attractive for CF lung gene therapy because they have enough DNA-carrying capacity and relatively high efficiency in airway gene delivery. 8,9 However, early CF gene therapy trials with Ad vectors were not successful 10 –12 because Ad vectors elicit both innate and adaptive immune responses that lead to the elimination of the vector-transduced cells. 13
To minimize the host immune responses, a helper-dependent adenoviral (HD-Ad) vector was developed by deleting all the viral coding sequences. 14,15 The deletion of viral genes allows for a further increase in DNA-carrying capacity of the vectors. More importantly, HD-Ad vectors are much less immunogenic with significant improvement in transgene expression in airway epithelial cells compared with the first-generation Ad vectors. 16 The improvement of transgene expression with HD-Ad vectors is believed to be because of the deletion of viral genes from the vector genome because the leaky expression of viral genes in the first-generation Ad vectors is known to cause host immune responses. We and others have successfully tested these vectors for gene delivery and shown that they are highly efficient in mediating transgene expression in lungs of mice, rabbits, pigs, and baboons via direct instillation or aerosol delivery. 9,17 –19
However, the capsid proteins of HD-Ad vectors elicit innate and adaptive immune responses, which compromise long-term therapeutic gene expression. 20,21 Several strategies, including host immune system modulation and vector modification, have been explored to reduce the immune reaction to viral vector and transgene products by using immunosuppressants and antibodies against CD4 or CD40L, CTLA4, as well as vector PEGylations. 22 –24 Using transient immuosuppression with cyclophosphamide, our group has successfully achieved readministration of HD-Ad vectors to mouse lungs. 25
In this study, we compared several regimens of transient immunosuppression with immunosuppressants and anti-inflammatory reagents, such as cyclophosphamide, dexamethasone, combination of cycolosporin A, azathioprine, and methylprednisolone and analyzed their effects on sustaining the expression of the CFTR gene. We investigated how these reagents modulate host immunity after HD-Ad vector delivery in mouse airways.
Methods
HD-Ad vector preparation and delivery to mice
HD-K18-CFTR and HD-K18-LacZ vectors were prepared as described previously. 16,19 Briefly, a number of serial passages on 116 cells allow high titer production of helper-dependent Ad5 vectors after three rounds of CsCl density gradient centrifugation. HD-Ad preparations were determined by quantified real-time PCR to contain <0.04% helper virus contamination. Ad particle numbers were calculated by absorbance at 260 nm. 26
For vector delivery, 8–10-week-old female C57Bl/6 mice (Charles River Laboratories) were anesthetized by isoflurane inhalation and 20 μl of HD-Ad vector in PBS, containing 40 μg/ml DEAE-dextran and 0.01% L-α-lysophosphatidylcholine (Sigma-Aldrich), was placed in small drops onto mouse nares, from which it was aspirated into the lungs. For transient immunosuppression studies on long-term human CFTR (hCFTR) expression, 5 groups of mice (10 for each group) were used in the experiments. One group without vector and drug treatment served as the control group and the base line for antibody analysis. The other four groups of mice received 1.5 × 1010 HD-K18-hCFTR vectors (vp/mouse); three of these groups were treated with cyclophosphamide (cytoxan; 50 mg/kg), dexamethasone (Dex; 5 mg/kg), or a triple-drug combination of cyclosporin (15 mg/kg), azathioprine (6 mg/kg), and methylprednisolone (25 mg/kg), respectively, at 6 hr before and 48 hr after vector administration by intraperitoneal injection. The group that received HD-Ad vector only was used as a control for transgene expression assays. Four months after vector delivery, the mice were sacrificed and blood samples were collected by cardiac puncture. Bronchoalveolar lavage was performed using 0.9 ml phosphate-buffered saline flushing three times. The lungs were placed in Trizol solution for RNA isolation or in optimal cutting temperature compound for frozen section. For measuring reporter LacZ expression, 2 groups of mice received 1.5 × 1010 of HD-K18-LacZ, and 1 group was treated with cyclophosphamide (50 mg/kg) by i.p. injection 6 hr before and 48 hr after vector administration. Lung tissues were collected and stored in β-galactosidase buffer for activity assay 100 days after lacZ gene delivery. For early data collection, mice were treated as above and lung tissues were collected 3 days after the treatment.
β-Galactosidase activity assay
Mouse lungs were homogenized in ice-cold buffer consisting of 100 mM K-phosphate pH 7.8, 0.2% Triton X-100, 0.5 mM dithiothreitol, and proteinase inhibitor cocktail (Roche Diagnostics) and total protein was determined by BCA Protein Assay Kit (Pierce). The homogenate was cleared by centrifugation at 4°C, and the supernatant was incubated at 48°C for 50 min to inactivate endogenous enzymes and assayed for β-galactosidase activity using a chemiluminescence kit (Galacto-Light; Applied Biosystems) with microplate luminometer (E.G.&G. Berthold LB96V). Values, expressed as relative light units, were normalized for total protein content.
RNA isolation and qRT-PCR
Total RNA from mouse lungs was isolated using RNAeasy kit (Qiagen) according to the manufacturer's instruction (Invitrogen), followed by DNase digestion. Reverse transcription was performed with 1 μg total RNA using random hexamers and SuperScript II reverse transcriptase following the manufacturer's protocol. For real-time RT-PCR, 10 ng cDNA was used as template in PCR with SYBR Green (ABI Prism 7500; Applied Biosystems). Primers for vector-specific CFTR were designed containing vector sequence. For relative quantification, PCR signals were compared among groups after normalization using 18S ribosomal RNA (Control Reagents; ABI) as an internal reference.
Western blot
Proteins isolated from IB3-1 cells 3 days after cyclophosphamide treatment and HD-K18-CFTR transduction were separated on a 7% SDS-PAGE and probed with anti-hCFTR (clone: 13-1; R&D) antibody followed by HRP-conjugated antimouse secondary antibodies (Vector Laboratories).
Determination of antibody titer
A pan-specific (IgA, IgE, IgGs, and IgM) ELISA for mouse antihuman Ad5 antibodies was performed as previously described. 26,27 A 96-well ELISA plate (Corning Costar) was coated with 5 × 109 vp of human Ad5 per well overnight at 4°C in 100 mM bicarbonate buffer pH 9.6. The plate was then washed with Tris-buffered saline, pH 7.5 (TBS) and blocked with 3% BSA in TBS. For titering of total Ig antibodies, mouse bronchoalveolar lavage fluid (BALF) was diluted in 1:20, or serum at 1:2000 in TBS, and added to the wells for overnight incubation. After washing with TBS, the plate was incubated with anti-mouse-Ig-biotin (BD Pharmingen) at 1:5000 dilution in TBS for 3 hr at room temperature. The plate was washed again and incubated with avidin-alkaline phosphatase (Sigma-Aldrich) diluted 1:50,000 in TBS, for 2 hr at room temperature. The plate was subsequently washed and incubated with 1 mg/ml p-nitrophenyl phosphate (Sigma-Aldrich) in 100 mM diethanolamine buffer pH 9.8, containing 0.5 mM MgCl2, for 10 min at room temperature. The reaction was stopped by the addition of 25 μl of 0.2 M EDTA and optical density was read at 405 nm.
Neutralizing Ab assay
The ability of mouse BALF or heat-inactivated serum to block Ad5 infection of HeLa cells was measured as described previously. Serum and BALF were heated at 56°C for 30 min. Diluted serum or BALF were incubated with 2.5 × 106 vp of HD-LacZ for 1 hr and then HeLa cells infected. After 3 days, cells were fixed and stained with X-gal solution. The highest dilution that resulted in a minimum 50% reduction in blue cells was recorded. When no reduction was observed the lowest dilution was conservatively assigned.
Analysis of airway inflammation
To analyze the infiltration of inflammatory cells in the airway, BALF was collected with 0.9 ml PBS and cells were collected for flow cytometry assay. CD45+ cells in BALF were stained with antibodies directly conjugated with fluorochrome, anti-CD45.2 (BD Biosciences). CountBright Absolute Counting Beads for flow cytometry (Invitrogen) were added into each sample before analysis on a FACS Calibur instrument (BD).
Cell culture
BEAS-2B and IB3-1 cells 28 were cultured in DMEM with 10% FBS and LHC-8 medium with 5% FBS, respectively, and treated with cyclophosphamide at 0, 15, 30, 80, and 200 μg/ml followed by HD-Ad-hCFTR vector transduction at 50 MOI (infectious vector particles per cell). RNA was isolated from cells 2 days after treatment, and vector-derived hCFTR mRNA was analyzed by RT-qPCR. Protein samples were prepared 3 days after treatment, and the hCFTR protein was detected with anti-CFTR antibody by Western blot.
Statistical analysis
There are 10 mice for each group. Student's t-test or one-way ANOVA followed by Holm's multiple comparison was used to compare data pairs or sets, respectively. Data are presented as mean ± SEM. p < 0.05 was considered significant (*p < 0.05, **p < 0.01).
Results
Long-term expression of the human CFTR and reporter LacZ in mouse lungs
To examine whether transient administration of immunosuppressants or anti-inflammatory reagents could improve transgene expression, three groups of mice were treated, respectively, with cyclophosphamide, dexamethasone, or the triple-drug combination of cycolosporin A, azathioprine, and methylprednisolone via i.p. injection 6 hr before and 48 hr after vector delivery. Human CFTR mRNA from mouse lung tissues was measured by real-time qPCR four months later. To determine exogenous CFTR with real-time PCR, the primers were designed to include vector sequences that do not recognize the endogenous mouse CFTR gene. We found that the level of human CFTR mRNA was 2.7 times higher in lungs of the mice treated with cyclophosphamide compared with the no-treatment control group. This result suggests that transient administration of cyclophosphamide significantly prolonged the transgene expression (Fig. 1). Interestingly, cyclophosphamide was found to be the most efficient among all drugs tested in this study for improving long-term transgene expression because the mRNA level of CFTR in this group was significantly higher than that in Dex and the drug combination groups (p < 0.01). Anti-inflammatory reagent dexamethasone did not affect the long-term expression of the human CFTR gene in mouse airways. There was no difference in mRNA expression of hCFTR between the Dex-treated and control mice. The mice treated with the triple-drug combination did not improve hCFTR gene expression and the expression level was lower, even though the difference was not significant compared with that of the control group.
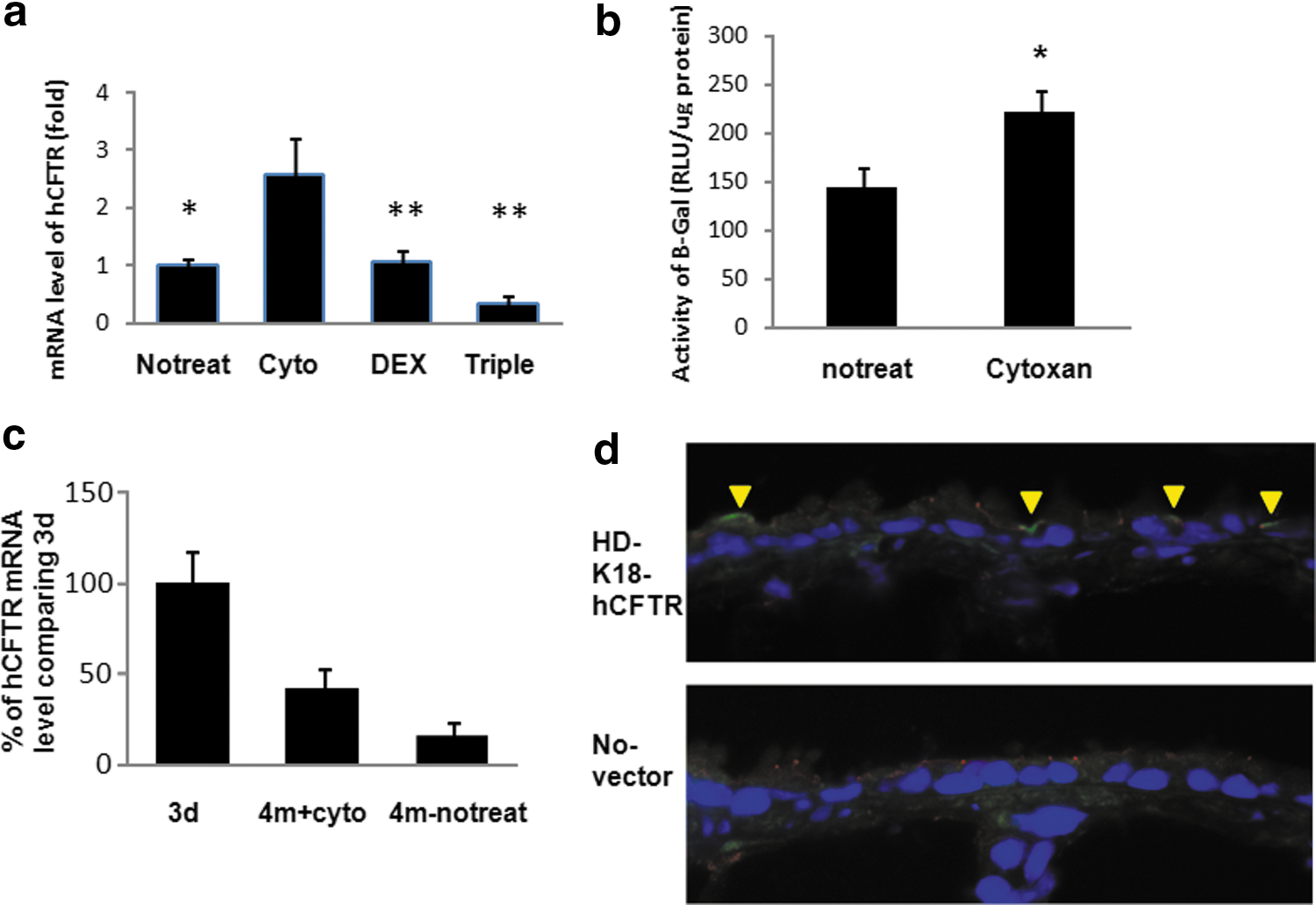
Expression of transgene in mouse lungs.
To further investigate whether the effect of cyclophosphamide on sustained expression of the CFTR transgene can be extended to other genes, we examined the long-term expression of the LacZ reporter gene in mouse lungs with similar treatments. Using chemiluminescent assays, we showed that the activity of β-galactosidase in mice treated with cyclophosphamide after 100 days was significantly higher than that of the group without treatment (p < 0.05) (Fig. 1b). This indicates that transient administration of cyclophosphamide can significantly improve the long-term expression of transgenes in mouse airways and the effect is not gene specific.
Because airway epithelial cells turn over and our gene therapy vector does not integrate into the genome of host cells, expression of transgenes in the airway is expected to decrease over time. To determine how much transgene expression remains 4 months after vector delivery, we compared the level of hCFTR mRNA between 4 months and 3 days after vector transduction. Assuming the level of expression at day 3 is the maximal (100%), we found that the human CFTR mRNA expression level at 4 months after cyclophosphamide treatment remained around 42% (Fig. 1c).
To investigate whether cyclophosphamide affects prolonged transgene expression or vector transduction, we collected lung tissues 3 days after gene delivery and determined the level of hCFTR expression. We found no significant difference in vector CFTR mRNA levels between mice with and without cyclophosphamide treatment (Supplementary Fig. S1; Supplementary Data are available online at
Anti-Ad antibodies and neutralizing antibodies in mouse BAL and serum
To investigate how cyclophosphamide enhances long-term transgene expression, we examined antivector antibody production in mice treated with different immunosuppressants.
The capsid proteins of HD-Ad vectors are known to induce innate and adaptive immune responses even though all viral genes are deleted. We measured anti-Ad antibodies in BALF and serum of groups of mice transiently treated with cyclophosphamide, dexamethasone, and the triple-drug combination by ELISA four months after vector delivery. We used a pan-specific antibody that detects multiple mouse Ig isotypes. As expected, levels of antibodies in BALF and serum of wild-type mice with no vector delivery were hardly detectable. Mice from the control group showed a significantly higher level of anti-Ad antibodies at this time point. However, Ab production in mice treated with cyclophosphamide was reduced dramatically in both BALF and serum compared with that of the control group (Fig. 2). Comparatively, the triple-drug combination also partially blocked antibody production in BALF, but not in serum. There was no difference in levels of anti-Ad antibodies between dexamethasone-treated mice and control.
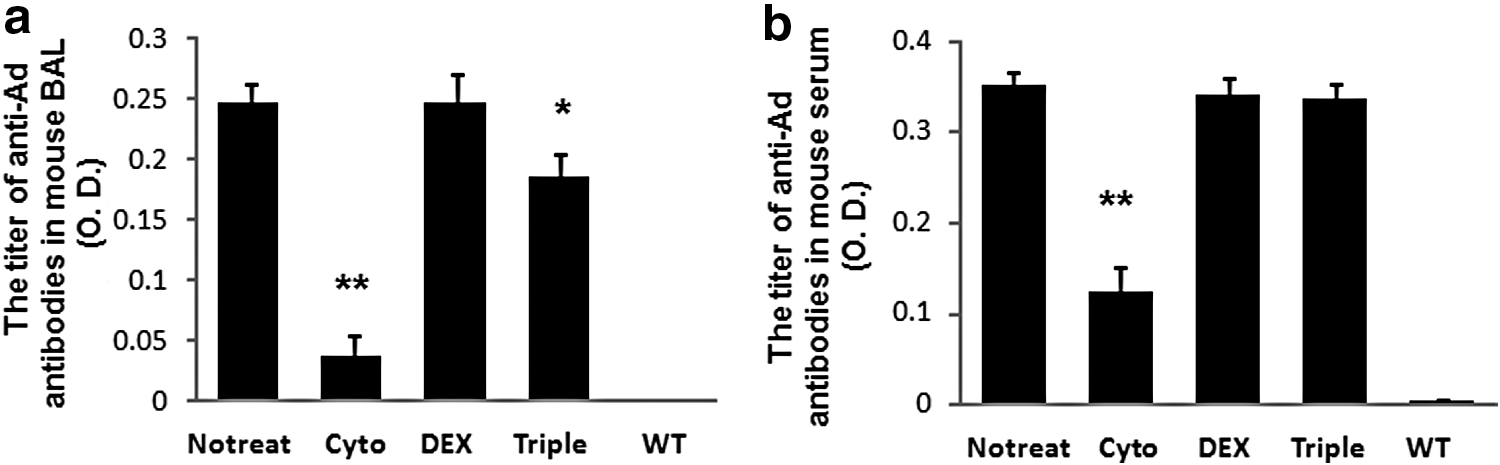
The titer of antiadenoviral (anti-Ad) antibodies in mouse serum and BALF. Serum
The presence of neutralizing antibodies, which inactivate Ad vectors, in mouse BALF and serum was also measured in all groups. Similar to the production of total anti-Ad antibodies, cyclophosphamide significantly inhibited neutralizing antibody production in both BALF and serum compared with the group with no treatment. Dexamethasone treatment partially decreased neutralizing antibodies in airway and blood system. However, the triple-drug combination did not affect neutralizing antibody production (Fig. 3).
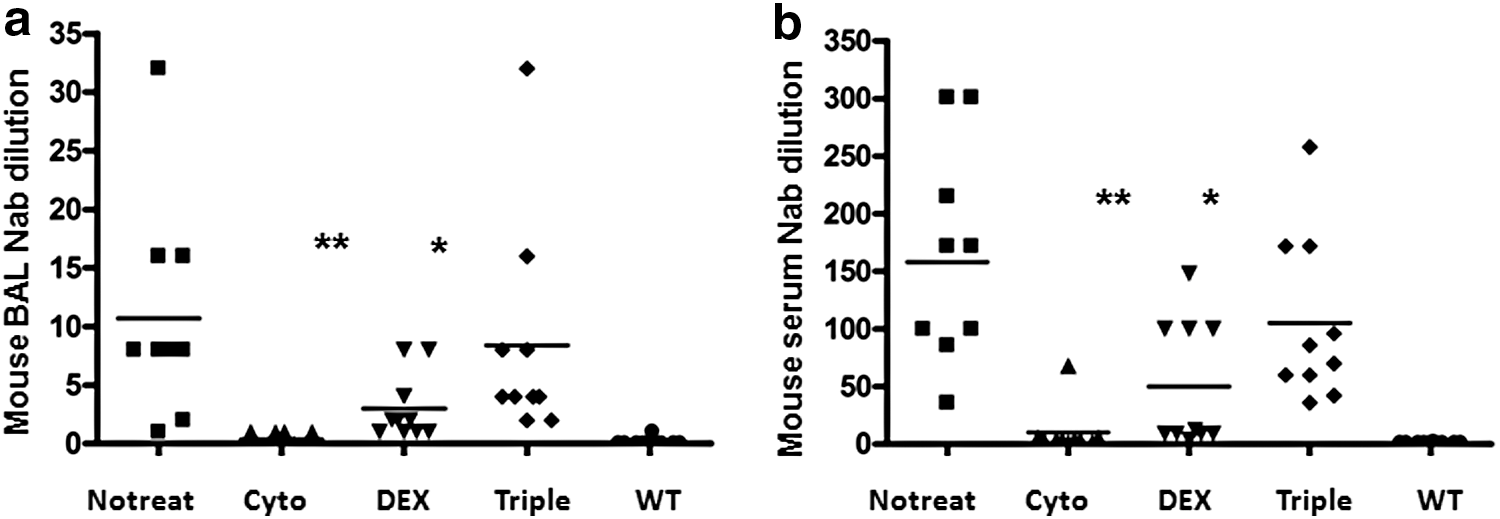
Neutralizing antibodies in mouse serum and BALF. Serum
Analysis of immune cells in the lung
To further understand the effect of transient immunosuppression and anti-inflammation on transgene expression, we analyzed the presence of immune cells in the lungs. We performed BALF collections 4 months after vector delivery and counted the total number of CD45 cells in BALF fluids of mice treated with different immunosuppressants. There was no difference in CD45+ cells and lymphocytes four months after vector administration among the groups of mice treated with cytoxan or Dex compared with the control no-treatment group and wild-type mice (Fig. 4a). Intriguingly, the number of CD45+ cells from the group of mice treated with the triple-drug combination was significantly higher than that from the other three groups (p < 0.05) (Fig. 4a and Supplementary Fig. S3). But the lymphocyte number in the triple-drug-treated mice was not significantly higher compared with any of the other three groups (Fig. 4b). We also detected granulocyte marker Gr-1 expression in mouse lung tissue four months after vector delivery; there was no difference among the groups (Fig. 4c).
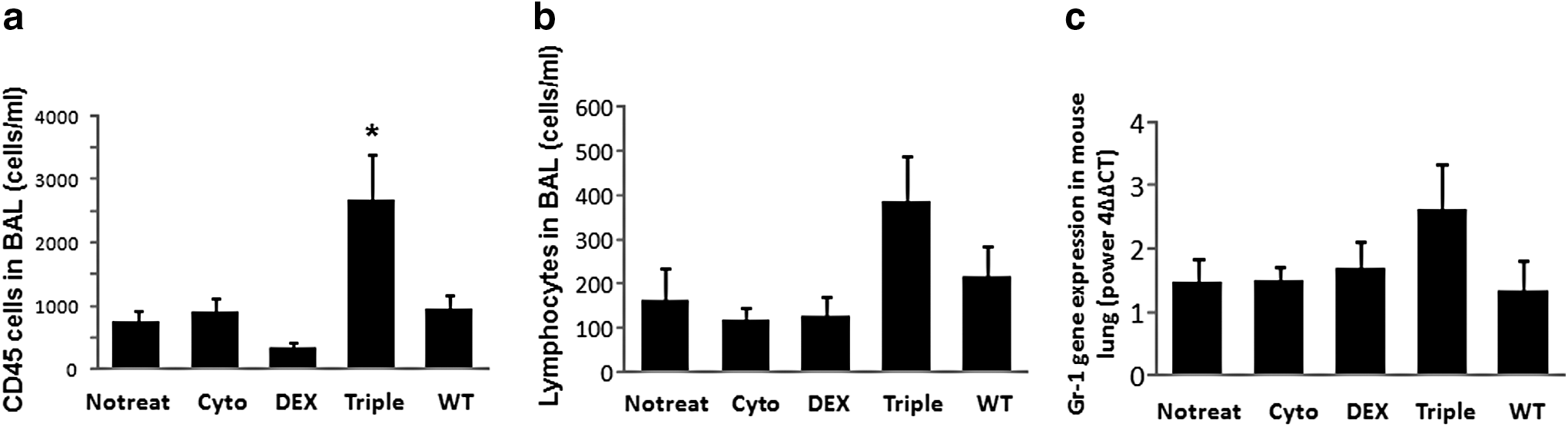
Infiltration of inflammatory cells and Gr-1 gene expression in mouse airways. CD45+ cells
CD4 and CD8 cells in the lung
To examine the cell-mediated adaptive immune reaction in the lung, CD8 and CD4 T-cells were analyzed with anti-CD8 and anti-CD4 antibodies by flow cytometry. However, the CD8+ and CD4+ cells were too low to be detected in mouse airway 4 months after vector transduction in all groups regardless of immunosuppression status (data not shown). Thus, we analyzed CD4 and CD8 mRNA expression in the lung issues with real-time RT-PCR. As shown in Fig. 5, we did not find any significant difference in CD3, CD4 and CD8 expression among the groups (Fig. 5).
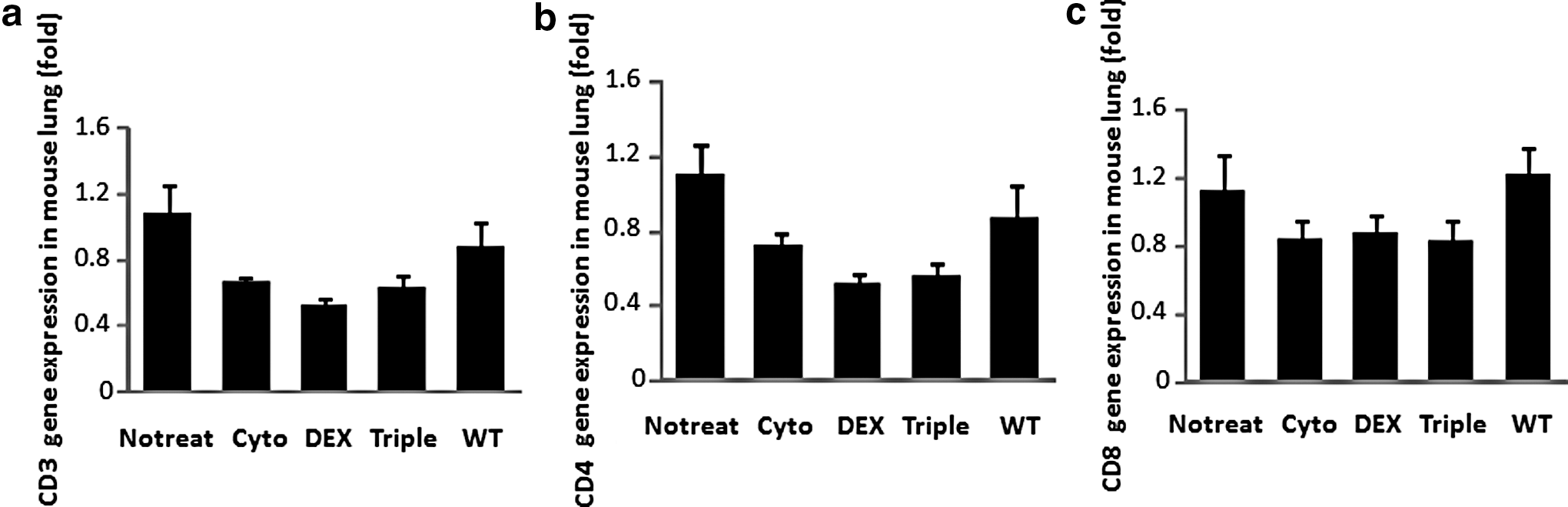
T-cell gene marker expression in mouse lungs. RNA was isolated from lung tissues 4 months after vector transduction, and real-time RT-PCR was performed with total mRNA from lung tissues.
Discussion
Immune response to vectors and transgene products is one of the major challenges for lung gene therapy with viral vectors. HD-Ad vectors have been shown to be highly efficient in gene delivery to lung airways of small 19 and large 17,18 animal models. However, because HD-Ad vectors share the same capsid proteins as the first-generation vectors, 9,14 host immune responses to the capsid proteins are expected to occur. Because HD-Ad do not contain viral coding sequences, there is no leaky expression of viral genes, which can cause host immune reactions, as in the case of the first-generation Ad vectors. Here we show that transient immunosuppression can be used to reduce the host immune reactions and subsequently to improve long-term transgene expression in vivo. Cyclophosphamide, dexamethasone, and the combination of cyclosporin A, methylprednisolone, and azathioprine have all been used for immunosuppression in animal studies 29 or clinical applications. 30 –32 In this study, we asked whether transient immunosuppression can enhance long-term CFTR expression, and if so, which drug is more effective. Because the triple-drug combination has been used in organ transplantation to suppress the immune reactions of recipients 30 –32 and in animal studies for enhancing gene delivery with the first-generation Ad vectors, 33 we tested the combination in our study as well. To our surprise, cyclophosphamide turns out be more efficient in improvement of both CFTR and LacZ expression in mouse airway by suppressing antibody production locally and systemically. Cyclophosphamide as an immunosuppressant is widely used in autoimmune disease and organ transplantation. 34 Cyclophosphamide also prolonged transgene expression from first-generation Ad vectors. 33 We previously also reported that cyclophosphamide affects CD4+ and CD8+ infiltration in airway in addition to preventing antivector antibody production. 25 Together with the previous published data, our results suggest that cyclophosphamide can be used to enhance sustained transgene expression mediated through HD-Ad vector delivery.
For the triple-drug combination, Cassivi et al. reported that they could consistently suppress immune reaction with this combination and increase transgene expression from first-generation Ad vectors by reducing inflammation and cellular immune responses in rat airways. 29 Surprisingly, this combination neither reduced anti-Ad antibodies production nor improved transgene expression in this study. Furthermore, there were even more CD45+ cells in airways of the treated mice after 4 months. Although it is not clear why this triple-drug combination did not work in our model, there are major differences between these studies. First, in our study, mice were treated with these drugs only twice during vector delivery, whereas Cassivi et al. administered them daily for the whole transgene expression period up to 35 days. Second, we assessed the long-term effects of the drug combination of vector delivery up to 4 months, whereas Cassivi et al. examined a relative short-term effect. Third, the first-generation Ad vector with viral gene E1 deleted induces a much stronger acute toxicity and immune responses as well as inflammation compared with helper-dependent vector. 16,35,36 Again, the immune reactions were suppressed transiently in this study, and so inflammation might come back again after immunosuppression was discontinued. 24,37 Finally, administration of the drug combination causes some acute epithelial cell injury in rat airways, 29 which may partially explain the less sustained transgene expression in our study.
Dexamethasone has been previously reported to induce increased expression of CFTR in cell lines within 24 or 48 hr of induction. 38,39 Dex, as an anti-inflammatory reagent, was also used to suppress inflammation and immune reaction in gene transfer with the first-generation Ad vector. However, it has no effect on transgene expression with transient administration even though cytokine secretion and antibody production significantly reduced in mice. 40 This is the first time that Dex was used for enhancing airway gene delivery with HD-Ad vectors. We found that Dex significantly inhibited neutralizing antibody production in airway and serum, but did not improve CFTR gene expression. The discrepancy in the effect of dexamethasone on CFTR expression between our in vivo study and those in vitro using cell lines is likely because of the difference in the experimental design. Our study was designed to examine the long-term effects, including the immune reaction, whereas the in vitro studies were to look into the immediate effect on CFTR expression. Although our observation is interesting, more studies are needed to investigate why Dex reduces airway immune responses but does not improve transgene expression.
There are limitations in this study. We did not investigate the immunosuppressant effect on innate immune reaction because cyclophosphamide did not reduce cytokine production. 25 To investigate why cyclophosphamide is more effective in transient immunosuppression, we assessed the antibody production and the presence of immune cells at the time when long transgene expression was determined. The design was to correlate the levels of transgene expression with the immune status of the mice. However, this was not the ideal time frame for assessing immune responses of the mice. Ideally, different time points after vector delivery should be assessed and each with enough numbers of mice. This aspect was not included in the original design of experiments because we did not know whether transgene expression could last for 4 months. However, since our goal was to achieve long-term transgene expression, the study has met our goal. Further studies of the host immune responses at different time points will be useful. For the same reason, the analysis of immune cell infiltration may underestimate the real degree of immune reactions produced by the vectors in our study.
Nevertheless, our results demonstrated that transient immunosuppression can be used to enhance long-term transgene expression in mouse lungs when HD-Ad vectors are used for gene delivery. This may be because HD-Ad vectors do not contain viral genes, which prevents them from continuously expressing viral antigen. Because the viral capsid proteins induce the early phase of immune responses peaked within 24 hr after vector delivery only if there is no viral gene transcription, 41 transient immunosuppression is sufficient to avoid the host innate immune response. This strategy may be effectively used to improve current lung gene therapy. Although cyclophosphamide has been clinically used, it is toxic. 42 It is possible that better drugs can be identified for this application. For example, mycophenolate mofetil may have a better safety profile for human applications. 43 In fact, mycophenolate mofetil together with tacrolimus has been used in clinical studies of cell transplantation to suppress host immune reactions. 44
Considering lung epithelial cell turnover, it is very impressive to observe transgene expression lasting for more than 4 months. Because the half-life of mouse ciliated airway epithelial cells is about 6 months, our result is not too surprising. However, for avoiding repeated gene delivery, it is necessary to integrate the gene expression cassette into a safe genome site in airway progenitor/stem cells. With the major progress made in the development of artificially engineered site-specific endonucleases, such as transcription activator-like effector nucleases (TALENs) 45 and clustered regularly interspaced short palindromic repeats (CRISPRs), 46 it is possible to make CF lung gene therapy a reality.
Footnotes
Acknowledgments
This study was partially supported by the Canadian Institutes of Health Research Grant MOP 125882 and a grant from the Cystic Fibrosis Canada to J.H.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.