
Abstract
Gene therapy holds promise to cure a wide range of genetic and acquired diseases. Recent successes in recombinant adeno-associated viral vector (rAAV)-based gene therapy in the clinic for hereditary disorders such as Leber's congenital amaurosis and hemophilia B encouraged us to reexplore an rAAV approach for pulmonary gene transfer. Only limited clinical successes have been achieved for airway gene transfer so far, underscoring the need for further preclinical development of rAAV-based gene therapy for pulmonary disorders. We sought to determine the preclinical potential of an airway-tropic serotype, rAAV2/5, encoding reporter genes when delivered to mouse airways. Although several groups have assessed the stability of gene transfer using a nonintegrating rAAV in mouse airways, long-term stability for more than a year has not been reported. Additionally, an extensive quantitative analysis of the specific cell types targeted by rAAV2/5 using cell-specific markers is lacking. We obtained sustained gene expression in upper and lower airways up to 15 months after vector administration, a substantial proportion of the lifespan of a laboratory mouse. In addition, we demonstrated that readministration of rAAV2/5 to the airways is feasible and increases gene expression 14 months after primary vector administration, despite the presence of circulating neutralizing antibodies. Finally, identification of transduced cell types revealed different subpopulations being targeted by rAAV2/5, with 64% of β-galactosidase-positive cells being ciliated cells, 34% club cells in the conducting airways, and 75% alveolar type II cells in the alveoli at 1 month postinjection. This underscores the therapeutic potential of a nonintegrating rAAV vector to develop a gene therapeutic drug for a variety of pulmonary disorders, such as cystic fibrosis, primary ciliary dyskinesia, and surfactant deficiencies.
Introduction
G
rAAV have emerged as promising gene transfer agents because of their favorable safety profile characterized by low immunogenicity, nonintegrating nature, and absence of viral genes. 9 Novel rAAV serotypes with distinct tissue tropism broaden the use of this vector for different gene therapy applications. 10 –12 We and others have shown efficient transduction of murine airway epithelial cells using the rAAV2/5 serotype. 13 –17 The safety of rAAV2/5 in human subjects was assessed and underscored in a clinical trial for acute intermittent porphyria (Clinical Trial Identifier, WCT02082860). In general, the lung is a slowly proliferative but rapidly regenerative organ, allowing prolonged gene expression using a nonintegrating vector, such as rAAV. Depending on the exact cell type, the lifespan of murine airway epithelium ranges from 6 to even more than 18 months. 18
In a previous study we assessed the efficiency and stability of rAAV2/5 reporter gene transfer into mouse airways in a perinatal setting. 19 As tissue proliferation in a growing organism is higher than in an adult animal, 20 we hypothesized that gene expression would be more stable after a single rAAV2/5 administration in an adult setting, compared with a single dose administered to fetuses or neonates. Therefore, as a follow-up, we set out to characterize rAAV2/5-based pulmonary gene transfer in adult mice to assess the applicability of this generic model for a range of respiratory diseases requiring long-term correction, such as CF, surfactant and alpha 1-antitrypsin deficiency, and primary ciliary dyskinesia.
Although stable gene transfer using a nonintegrating rAAV has been documented before in mouse airways using histology or tissue extracts, 15,16 we additionally used noninvasive bioluminescence imaging (BLI) combined with an extensive quantitative analysis of the specific cell types targeted by rAAV2/5 to fully characterize long-term stability after rAAV2/5-mediated gene transfer to mouse airways. We coadministered two rAAV2/5 vectors, each encoding a single reporter (either firefly luciferase [rAAV2/5-fLuc] or β-galactosidase [rAAV2/5-β-gal]) intratracheally in adult FVB mice, and assessed the stability of gene expression up to 15 months postinjection (pi), which is approaching the lifespan limit of a laboratory mouse (1.5–2 years). Preexisting or vector-induced neutralizing antibodies (nAb) against the AAV capsid can pose an obstacle to successful readministration in an adult organism with a fully mature immune system. 21,22 Other studies showed the feasibility to readminister rAAV, despite high-level circulating nAb. 16,17 We sought to determine the kinetics of nAb after a single rAAV2/5 administration and to evaluate if readministration is feasible as soon as serum nAb decline.
Our results demonstrate that rAAV2/5 transduces both upper and lower mouse airways (nasal mucosa and lung) and that transduced cells remain present until 15 months pi. Using cell-specific markers, transduced cell types were identified as being ciliated, club (previously known as Clara), or alveolar type II (ATII) cells, proffering rAAV2/5 for different lung-targeted gene therapy applications. In addition, this vector was successfully readministered 14 months after primary vector administration, resulting in increased gene expression in the lung. Taken together, this study demonstrates the therapeutic potential of rAAV2/5 for pulmonary gene transfer in a preclinical setting, suggesting further development of an rAAV2/5-based gene therapy for human airway diseases.
Materials and Methods
Viral vector production
rAAVs were produced by triple transient transfection using transfer (AAV-TF-enhCB-LacZnls 16 and AAV-TF-enhCB-eGFP-P2A-fLuc 13 ), packaging (AAV rep/cap), and helper (pAdbDeltaF6) plasmids in a ratio of 1:1:1 in HEK293T producer cells. The transgenes are under the transcriptional control of the cytomegalovirus-enhanced chicken β-actin (enhCB) promoter. 23 Vectors were purified by iodixanol gradient centrifugation as previously described. 19,24 Vector titers were determined by real-time PCR using the primer probe set for the polyA sequence and presented as DNase resistant GC/ml. 10,23
Animal studies and vector administration
For all experiments, FVB mice were used (Janvier). Intratracheal vector instillation was performed in anesthetized adult mice 8–12 weeks of age. After exposure of the trachea through a midline incision, 160 μl of vector diluted in PBS (rAAV2/5-fLuc [2 × 1011 GC/animal]; rAAV2/5-β-gal [4 × 1011 GC/animal]) was instilled into the trachea with a 26 G PTFE catheter (BD Vasculon Plus; VWR) using a stereoscopic zoom microscope (3× magnification). Animals injected with the same volume of physiological saline were taken along as negative controls. Of note, two animals died during the experiment at 8 months after vector administration. All animal procedures were approved by the KU Leuven ethics committee (laboratory accreditation number LA1210579; ethics committee approval number P047-2013).
Bioluminescence imaging
BLI was performed as previously described. 13 Measurements are reported as the total photon flux from a 4.3 cm2 rectangular region of interest (ROI) for the lung and a 2.0 cm2 circular ROI for the nose. Of note, 5 animals were excluded from BLI measurements at 1 month pi because of dehydration.
Power regression analysis
BLI data for lung and the nose measurements of animals monitored during the whole span of the experiment were fit according to a nonlinear power regression model (y = Axp ). The data were fit to a linear regression model after a log–log transformation on the data set (x = time, y = BLI signal), so ln(y) = ln(Axp ) → ln(y) = ln(A) + [p × ln(x)]. From this linear regression, the constant p in the original formula y = Axp was calculated, being the slope of the linear regression curve ln(y) = [p × ln(x)] + b. From the constant b = ln(A), which is the intercept in the linear regression formula, the constant A in the original formula was derived by applying the natural exponential function to b; that is, A = eb . The R 2 represents how close the log–log-transformed data are to the fitted linear regression line.
Quantification of vector genomes in lung tissue
Animals were sacrificed at regular time points and lung tissue was frozen according to the protocol described previously. 19 Genomic DNA was isolated from lung tissue using a mammalian genomic DNA miniprep kit (Sigma). Vector genomes were quantified by real-time PCR using a primer–probe set for the polyA sequence as described in the section Viral vector production. Genomic DNA input was normalized to endogenous GAPDH using the following primer set with SYBR green (Roche) detection: forward 5′-ACGGCAAATTCAACGGCAC and reverse 5′-TAGTGGGGTCTCGCTCCTGG. 25 Results are presented relative to the average of the group harvested at one month pi. A difference of 5 Ct was observed between the animal with the lowest rAAV genome copies and the negative control animal.
X-gal staining for β-gal expression in tissue sections
Processing of mouse lung and nose samples and X-gal staining on tissue sections were performed as described previously. 13,19 X-gal staining was followed by Mayer's paracarmine counterstaining. β-Gal-positive cells were visualized using a Leica Biopoint 2 light microscope. Brightness and contrast were optimized using Adobe Photoshop CS6.
Immunohistochemistry and quantification of transduced cells
Immunohistochemical analysis on nose and lung sections was performed in parallel with X-gal staining, as a subsequent step in the protocol. Sections were pretreated with 3% hydrogen peroxide and incubated with the following antibodies: mouse anti-β-tubulin (Sigma T4026; 1/1000), rabbit anti-CCSP (Millipore; 07-623, 1/10000), and rabbit anti-SP-C (Santa Cruz; FL-197; sc-13979, 1/500), which recognize β-tubulin, club (Clara) cell secretory protein (CCSP), and surfactant protein C (SP-C), respectively, in 10% goat serum overnight at 4°C. After three washing steps (1× PBS-0.1% triton-X, 2× PBS), sections were incubated for 1 hr at room temperature with biotin-conjugated secondary antibody anti-mouse or anti-rabbit IgG (1/300; DakoCytomation), followed by incubation with streptavidin–horseradish peroxidase complex (DakoCytomation). Protein immunoreactivity was visualized using 3,3-diaminobenzidine (DAB, 0.4 mg/ml; Sigma-Aldrich) as a chromogen. Sections were counterstained with paracarmine and mounted with mowiol, a water-soluble hydrocolloid mucoadhesive based on polyvinyl alcohol (8% mowiol 4-88 [cat. no. 81381; Sigma], 20% glycerol in 0.1 M Tris HCl buffer pH 8.5). Double-positive cells were visualized using a Leica Biopoint 2 light microscope. Brightness and contrast were optimized using Adobe Photoshop CS6.
Quantification of positive cells was performed using the Cell-Counter plugin from ImageJ. The number of β-gal-positive cells within a high-magnification image (40×) was quantified using a first marker. Within the transduced (i.e., β-gal positive) population, a second marker was used to quantify specific cell populations based on marker staining (i.e., β-tubulin for ciliated cells; CCSP for club cells; SP-C for alveolar type II cells). Next, the ratio of double-positive cells (i.e., β-tub+ β-gal+, CCSP+ β-gal+, or SP-C+ β-gal+) in the total β-gal+ population in the conducting airways for ciliated and club cells or in the alveolar region for ATII cells was calculated. Ten high-magnification images (40×) were analyzed per animal, per region (conducting airways for β-tubulin and CCSP; alveoli for SP-C) and averaged per time point (1 and 15 months, respectively). Data are presented as mean ± SEM.
rAAV transduction inhibition assay
Individual blood samples were collected at different time points from animals by retro-orbital bleeding. Serum samples were heat-inactivated for 45 min at 56°C. The rAAV transduction inhibition assay was performed as described previously with minor modifications. 19 Briefly, HEK293T cells stably expressing renilla luciferase (rLuc) were incubated with a mixture of rAAV2/5-fLuc (∼1 × 109 GC/well) and 4-fold dilutions of serum samples prepared in DMEM without FCS. Wells with the same volume of DMEM served as no-serum controls. Twenty-four hours later, the transduction efficiency was analyzed by quantifying fLuc activity normalized to rLuc activity. The percentage of inhibition was calculated relative to no-serum controls.
Luciferase assay
HEK293T cells stably expressing rLuc and transduced with rAAV2/5-fLuc were lysed in 70 μl of lysis buffer (50 mM Tris pH 7.5, 200 mM NaCl, 0.2% Nonidet P-40 [NP-40], 10% glycerol) and the lysate was assayed according to the manufacturer's protocol for both rLuc and fLuc activity (ONE-Glo and Renilla-Glo luciferase assay system; Promega).
Statistical analysis
Linear regression analysis was performed to obtain the Pearson's correlation coefficient on BLI and real-time PCR data. Statistical analysis for the effect of readministration analyzed on BLI data was performed using a paired t-test for comparison of the BLI signal before and after readministration and an unpaired t-test for comparison between the readministered group and naive controls. Statistical tests were performed after logarithmic transformation on the original data. Statistical analysis for rAAV transduction inhibition assay data analyzed at dilution 1/2560 was performed using repeated measures ANOVA with Tukey's multiple comparison post hoc test. All statistical analyses were performed using GraphPad Prism 5 software. A p < 0.05 was considered statistically significant.
Results
Long-term persistence of gene expression in mouse airways for 15 months after a single dose of rAAV2/5
To assess the stability of gene expression, two rAAV2/5 vectors (rAAV2/5-fLuc and rAAV2/5-β-gal) were co-administered in adult FVB mice (n = 30) by intratracheal injection (Fig. 1A). FLuc expression was monitored monthly by noninvasive BLI up to 15 months pi (Fig. 1B). At specific time points (1, 3, 6, 12, and 15 months pi) animals were sacrificed for histological analysis of β-gal expression in the lung and the nose, respectively. The average photon flux measured in the lung region dropped only 4-fold between 1 and 15 months pi, remaining 5-fold above background (negative control animals) (Fig. 1C). The mean photon flux emerging from the nasal region declined 27-fold, but remained 1 log above background at 15 months pi (Fig. 1E). Since groups of animals were selected throughout the experiment for histological analysis, group sizes decreased over time. Therefore, the BLI kinetics are additionally shown for the group of animals (n = 6) monitored at all time points, showing a similar decline and underscoring the lack of selection bias (Supplementary Fig. S1A and B; Supplementary Data are available online at

Noninvasive BLI after rAAV2/5 reporter gene delivery reveals persistent gene expression in mouse airways for 15 months.
In general, kinetics depend on the subject age at the start of the experiment, the species and animal strain, the type of vector, vector dose, transgene(s) encoded, administration route, and target organ(s). As apparent in the fold decrease measured for both the lung and nasal region (4- and 27-fold, respectively) over a time period of 15 months, the nonlinear regression model similarly expresses the decline in gene expression as being moderate for lung (Fig. 1D), compared to a more pronounced decrease for the nasal region (Fig. 1F). In addition, we quantified relative rAAV genome copies in the lung by real-time PCR. The results demonstrated a gradual decrease in vector genome copies over time (Fig. 1G), strongly correlating with the kinetics of luciferase activity (R 2 = 0.95, p = 0.0042; Fig. 1H). Interestingly, in a repeat experiment in which only a single rAAV2/5 vector (i.e., rAAV2/5-fLuc) was administered intratracheally to adult FVB mice (n = 8), photon flux from both upper and lower airways was relatively more stable over a period of 6 months (end of experiment), showing an overall decrease in fLuc expression of only 1.8-fold in the lung and 2.5-fold in the nasal region (Supplementary Fig. S2A and C), which is similarly described when the data are fitted to a nonlinear regression model (Supplementary Fig. S2B and D for lung and nose, respectively). This suggests that the type and the number of transgenes transferred by the vector play a role in the stability of gene expression. Taken together, both experiments show long-term persistence of gene expression in mouse airways after a single dose of rAAV2/5 in an adult setting.
rAAV2/5 efficiently targets the respiratory epithelium of the upper and lower mouse airways
After demonstrating long-term expression of fLuc as measured by noninvasive BLI, we set out to identify the specific cell populations targeted by rAAV2/5 after intratracheal delivery. Histological analysis of the upper (nose) and lower (lung) mouse airways revealed β-gal-positive cells throughout examined tissue. In line with the BLI results, β-gal-positive cells were observed at all time points (1, 3, 6, 12, and 15 months pi) in both the nasal mucosa and the lung epithelium (Fig. 2), confirming sustained transgene expression in mouse airways after a single dose of rAAV2/5. In the nasal mucosa, rAAV2/5 transduction was restricted to the respiratory epithelium, with no β-gal-positive cells detected in the olfactory epithelium, indicating the specific airway tropism of this rAAV serotype (Fig. 2). β-gal gene expression levels decreased over time (representative images shown; Fig. 2), similar to the observed decline in fLuc activity between 1 and 15 months (Fig. 1). In the lung, transduced cells were observed both in the conducting airways as well as in the alveoli (Fig. 2). For both regions, a decrease in β-gal-positive cells was observed at later time points, though this was more pronounced in the conducting airways (Fig. 2). Of note, β-gal-positive smooth muscle cells were detected in blood vessels throughout the lung parenchyma (data not shown). In conclusion, these data are in line with the BLI data and additionally demonstrate at the histological level that rAAV2/5 mainly transduces airway epithelial cells after intratracheal delivery.

Persistent transgene expression for 15 months after rAAV2/5 transduction in upper and lower mouse airways. β-Galactosidase (β-gal) expression (blue) in the respiratory epithelium of upper (nose) and lower (lung) airways after intratracheal administration of rAAV2/5 in adult FVB mice. Sections were counterstained with paracarmine (pink). Representative images of lung and nasal cross sections at 1, 3, 6, 12, and 15 months pi. Scale bar = 50 μm. Color images available online at
Next, we identified and quantified the specific cell types transduced by rAAV2/5 using cell-specific markers for immunohistochemical analysis. β-Tubulin, club (Clara) cell secretory protein (CCSP), and surfactant protein C (SP-C)-specific antibodies were used to identify ciliated cells, club cells, or ATII cells, respectively, in the β-gal-positive cell population (Figs. 3 and 4). In the nasal mucosa, the majority of transduced cells were β-tubulin-positive, pinpointing ciliated cells as the cell type predominantly transduced in the respiratory epithelium (Fig. 3). In the conducting airways of the lung, 64% of β-gal-positive cells were co-stained with β-tubulin at 1 month pi (Fig. 4A) and 34% with CCSP (Fig. 4B). The relative distribution of ciliated and club cells in the conducting airways did not change at 15 months pi (Fig. 4A and B). However, the number of transduced ATII-positive cells markedly declined over time, starting with 75% SP-C-positive cells at 1 month and ending with 31% positive cells at 15 months pi (Fig. 4C), indicating that relatively more other cell types were β-gal-positive at 15 months. Taken together, since different cell populations in the airway epithelium were efficiently transduced by this vector, these results underscore rAAV2/5 as a promising viral vector to target the respiratory epithelium for a wide range of pulmonary disorders, such as CF, primary ciliary dyskinesia, and surfactant deficiencies.
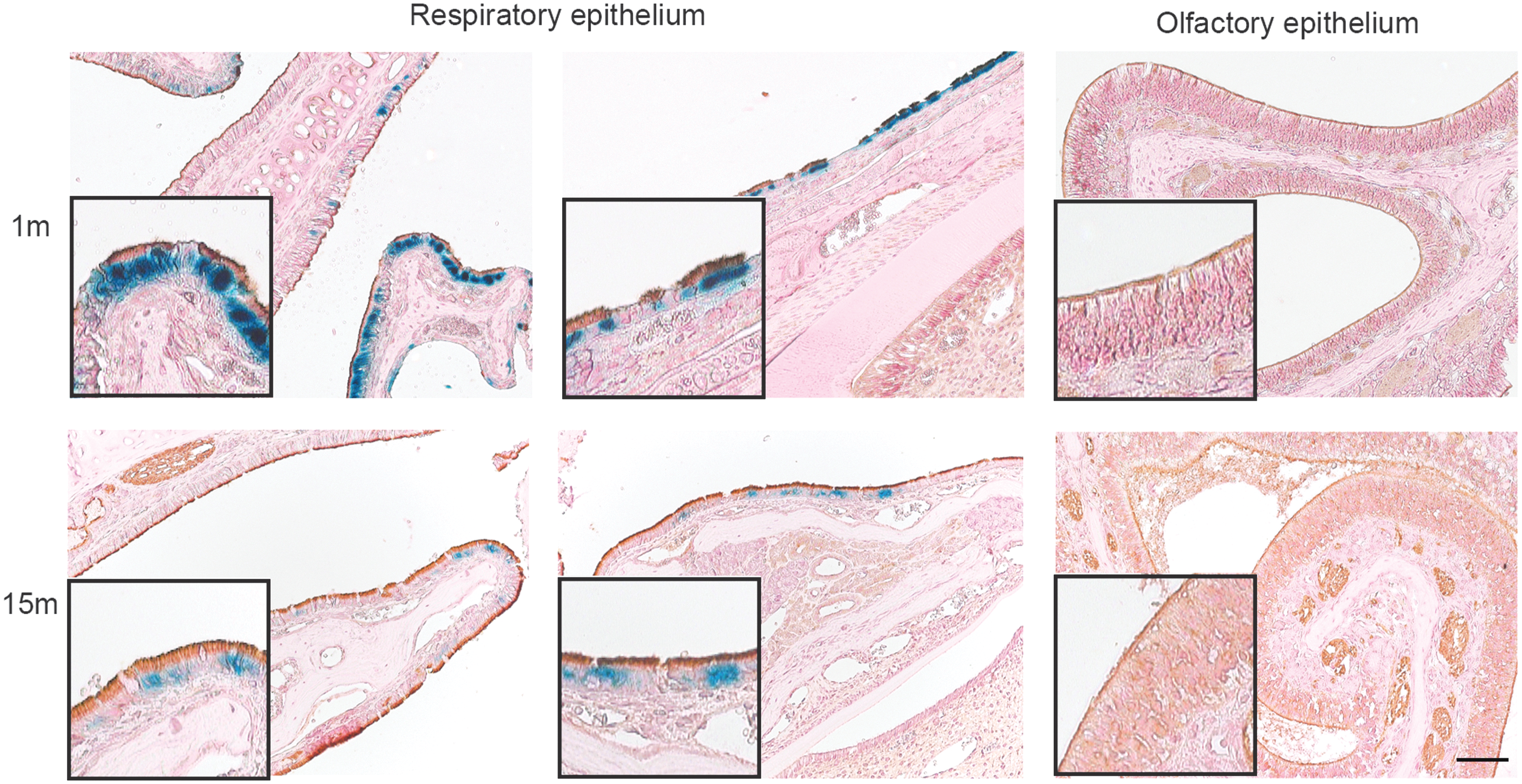
rAAV2/5-mediated reporter gene transfer efficiently targets the respiratory epithelium of the mouse nose. β-Gal-positive cells (blue) in the respiratory epithelium of the nose were identified as ciliated cells by β-tubulin marker staining (brown) at 1 and 15 months pi. Sections were counterstained with paracarmine (pink). The insets depict enlarged images of the regions with double-positive cells (β-gal+, β-tubulin+) in respiratory epithelium and absence of β-gal+ cells in olfactory epithelium. Scale bar = 50 μm.
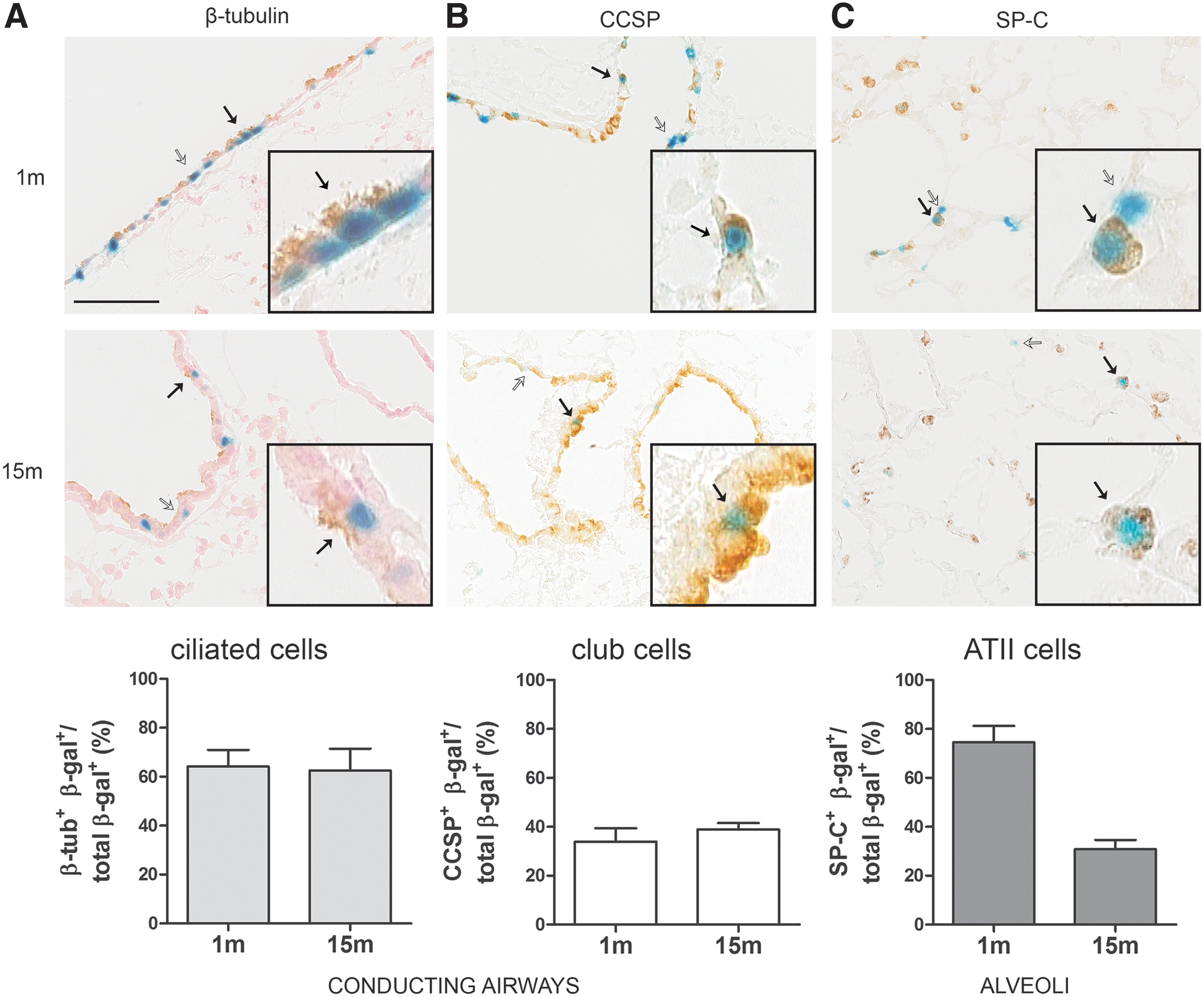
rAAV2/5-mediated reporter gene transfer targets distinct subpopulations of cells in the mouse lung. β-Gal-positive cells (blue) in the lung were identified by a cell-specific marker (brown) as
rAAV2/5 serum neutralizing antibodies decline over time
The humoral immune response against the rAAV2/5 capsid was assessed by collecting sera at 1, 6, and 14 months pi (Fig. 1A). The respective sera were tested for the presence of nAb in an rAAV transduction inhibition assay (Fig. 5). In this assay, rAAV2/5-fLuc was incubated with serum samples of treated animals before HEK293T cell transduction: nAb hamper rAAV2/5 transduction and result in reduced transduction efficiencies. The relative amount of nAb is obtained by measuring the degree of transduction inhibition expressed as relative light units relative to no-serum control. Based on this functional assay, nAb were observed as soon as 1 month pi with an almost complete inhibition even at dilution 1/2560 (95% relative transduction inhibition; Fig. 5A). rAAV2/5 serum nAb decreased approximately four-fold between 1 and 6 months; at 6 months pi 54% transduction inhibition was detected at dilution 1/2560, whereas the same level of inhibition was measured 1 month pi at a four-fold higher dilution (Fig. 5A). Neutralizing Ab titers decreased further by 14 months after rAAV2/5 vector administration, though to a less extent than the decline measured in the first 6 months (Fig. 5B).
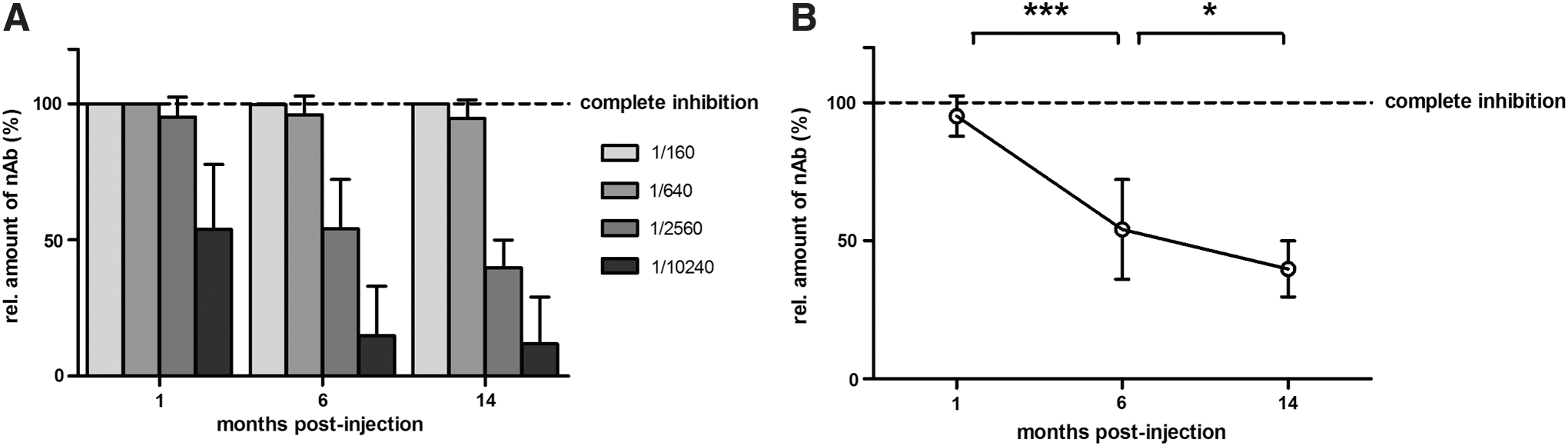
Serum neutralizing antibodies (nAb) against rAAV2/5 decrease over time in treated mice. Analysis of nAb against the rAAV2/5 capsid after intratracheal vector administration in adult FVB mice (n = 5) using an rAAV transduction inhibition assay.
Readministration of rAAV2/5 at 14 months after the first vector dose is feasible
To achieve lifelong gene correction by rAAV-mediated gene transfer in a proliferative tissue such as the lung, repeated vector administrations will most likely be necessary. Considering the strong reduction in nAb 14 months pi, we assessed the feasibility of repeated intratracheal administration of rAAV2/5 into mouse airways (Fig. 6A). Fourteen months after the initial vector dose, animals were divided into two groups based on their BLI signal emerging from the lung region. The second vector dose was administered to the group showing the lowest BLI signal emerging from the lung (n = 5) and fLuc activity was measured 1 month later (Fig. 6B and C). As a control group, age-matched naive animals (n = 5) without prior exposure to the vector were taken along. FLuc activity in the lung and the nasal region was compared before and after readministration for each animal, that is, at 14 months after the first vector dose (Fig. 6B and C “before readmin.”) and at 1 month after readministration (Fig. 6B and C “1m post readmin.”). A significant 2.6-fold increase (p = 0.0032) in photon flux was observed in the lungs of readministered animals (Fig. 6B). However, fLuc expression after the second vector dose was significantly lower than that observed in the immunologically naive control group (Supplementary Fig. S3). For the nasal region, in 3 out of 5 mice increased gene expression levels were observed, resulting in an average 2-fold increase in BLI signal after readministration. However, this increase was not statistically significant (p = 0.17) (Fig. 6C). Taken together, rAAV2/5 readministration 14 months after primary vector administration is feasible and results in increased reporter gene expression in the mouse airways.
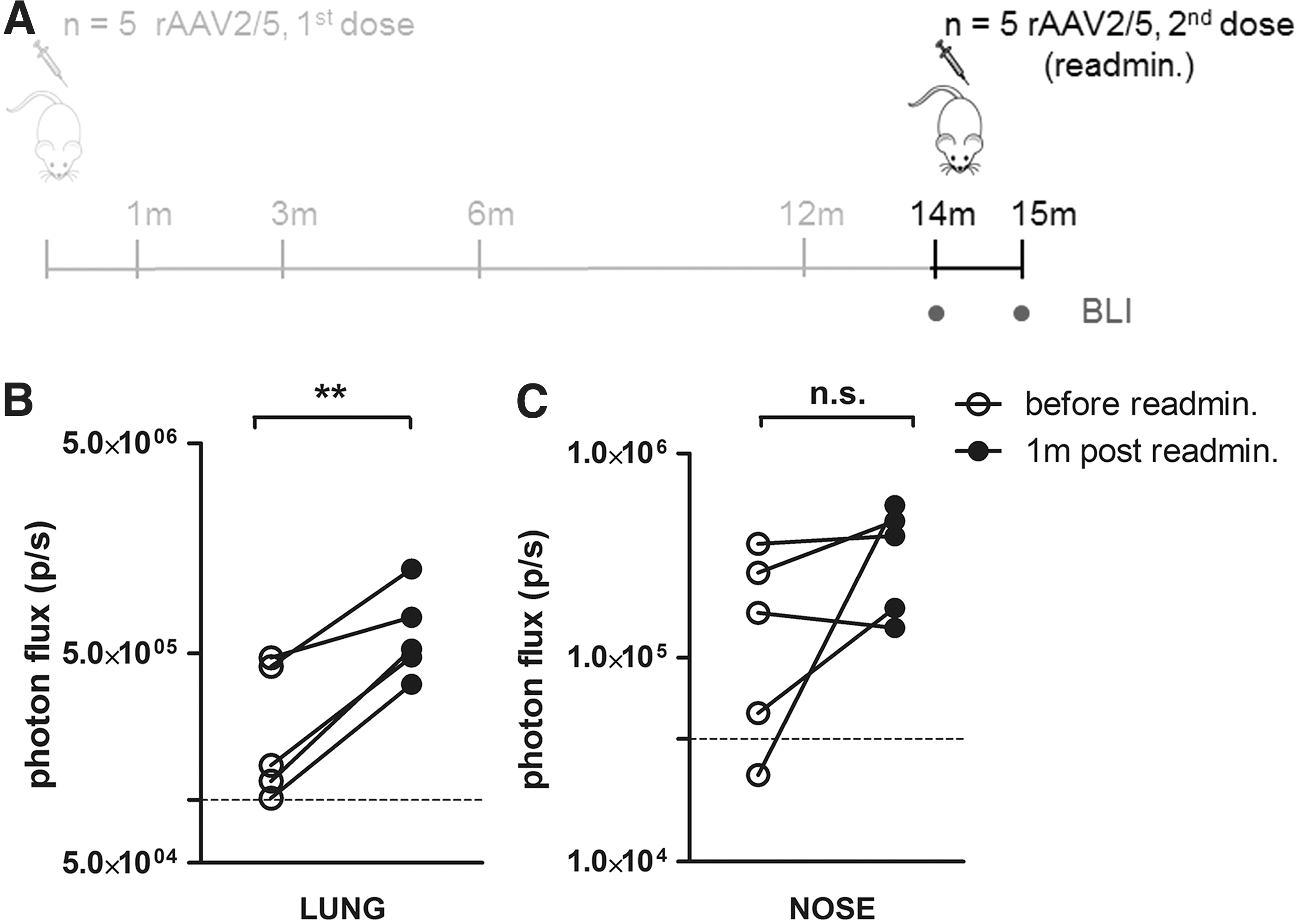
Readministration of rAAV2/5 is feasible in mouse airways.
Discussion
The aim of this study was to fully characterize the stability of gene expression using a nonintegrating rAAV vector and identify the cell types targeted, thereby providing a fully characterized preclinical animal model that allows investigation of rAAV-based gene therapy for a variety of pulmonary disorders. As there is a substantial body of evidence in the literature supporting efficient gene delivery by rAAV2/5 to mouse airways, 14,15,17 including our previous work, 19 we specifically chose this rAAV serotype for our study. A first interesting finding was the sustained gene expression obtained up to 15 months after a single rAAV2/5 dose in mouse airways, which is approximately the maximum length that adult-inoculation experiments can be tested for in the mouse, as it approaches the lifespan limit. Second, characterization of transduced cell types identified different subpopulations being targeted by rAAV2/5 (ciliated, club, ATII cells), underscoring the therapeutic potential of this vector to develop gene therapy for a variety of pulmonary disorders. Third, despite the induction of nAb against the rAAV2/5 capsid after the first vector dose, they significantly diminished over time, allowing successful readministration to mouse airways 14 months after primary vector dose.
The stability of gene expression is a crucial aspect to treat and ultimately cure genetic disorders that require lifelong correction. In addition to previous studies that have addressed long-term stability after rAAV-mediated airway gene transfer using histology or tissue extracts, 15,16,26 we use BLI to follow up fLuc expression in mouse airways. BLI is a noninvasive method for the detection and quantification of fLuc gene expression in vivo allowing gene expression to be followed up over time in the same animal, and so detailed kinetics can be obtained. Therefore, the number of animals exposed to experimentation can be reduced. We show in this study persistent gene expression over a period of 15 months for both the lung and the nasal mucosa, which has not been reported before. However, we report a gradual decline in gene expression, which likely presents a dilution of episomal rAAV genomes because of proliferation of transduced tissue or immune response-mediated destruction of transduced cells. This was corroborated by quantification of vector genomes over time in lung tissue. The gradual loss of rAAV copy number over time showed a significant correlation with the decrease measured in luciferase activity for the lung, suggesting that the decrease in gene expression was not because of epigenetic silencing of vector-mediated transgene expression.
In contrast to the lung, fLuc expression in the nasal mucosa declined faster (27- vs. 4-fold). Since the respiratory epithelium of the nasal mucosa is directly exposed to the outside environment, we speculate that the turnover rate of nasal epithelium is higher than that of the conducting airways and lung parenchyma. An independent experiment in which only a single rAAV2/5-fLuc vector was administered to adult mouse airways demonstrated stable gene expression up to 6 months (Supplementary Fig. S2). Strikingly, the decrease in reporter gene expression was far less pronounced over this 6-month time period, in particular in the upper airways (1.8- and 2.5-fold for the lung and the nasal region, respectively, compared with a decrease of 3- and 11.2-fold when two rAAV2/5 vectors encoding fLuc and β-gal were co-administered). One explanation for these results is that administration of an additional antigen, in this case an immunogenic reporter like β-gal, could reduce gene expression by triggering a destructive immune response. Since the stability of gene expression is transgene dependent (reviewed in ref. 27 ), we speculate that this may reflect a difference in cellular immune response mounted against the transgene(s), which in turn affects the stability of gene expression by destruction of transduced cells, although a direct comparison would be necessary to confirm this observation. In that light, gene transfer of a therapeutic gene encoding a self-protein such as CFTR would most likely induce a far less pronounced immune response compared with non-self-immunogenic reporters.
By assessing the specific cell types transduced, we further characterized the translational potential of the rAAV2/5-based gene therapy in mouse airways as a preclinical model for a variety of pulmonary disorders, as the cell types transduced relate to the type of disease to be cured. Identification of transduced cell types in the airways mostly is based on morphology, although some groups have additionally assessed this using cell-specific markers. 15,28 In this study, we performed a detailed quantitative analysis of different cell populations in the airways at the beginning and the end of the study (1 and 15 months, respectively), which has not been reported before. Detailed histological analysis of the lung revealed preferential transduction of ciliated (64%), club (34%), and ATII cells (75%) at 1 month pi, and predominantly ciliated cells in the respiratory epithelium of the nasal mucosa, proffering rAAV2/5 as a suitable candidate for gene therapy for CF, primary ciliary dyskinesia, or surfactant deficiencies in particular. In contrast, a study by Sumner-Jones et al. in which rAAV5/5 was instilled intranasally in BALB/c mice showed preferential targeting of sustentacular cells of nasal olfactory epithelium and ATII cells in the alveolar epithelium, with only few reporter expressing club cells and no ciliated cells in the conducting airways. 15 The discrepancy in airway transduction with our study could be because of the different mouse strains used, the administration route, and/or the promoter. An additional difference is the viral vector itself, rAAV5/5 compared with our vector, rAAV2/5, pseudotyped with the AAV5 capsid, but containing the AAV2 genome. Yet, this should not affect vector tropism, since both vectors are encapsidated with capsid proteins from AAV serotype 5, which ultimately determines vector tropism. However, it cannot be excluded that postentry steps might differ between both types of rAAV vectors, such as second-strand synthesis or a ubiquitin/proteasome-dependent block in nuclear uptake of the vector in a cell type-specific manner. 29,30
The mouse nasal mucosa reflects the human lung closely based on cell composition and bioelectric properties. 31 In contrast, the mouse lung differs from the human lung, in particular with regard to the relative abundance of club cells in mice, the restriction of basal cells to proximal regions of the conducting airways, and finally the paucity of submucosal glands. 32 These factors could influence transduction efficiency, highlighting the importance of performing comparative analyses in other animal models that phenocopy the human situation more closely. Therefore, the appropriate rAAV serotype should be selected for each species studied. In that regard, rAAV2/1 has been shown to efficiently transduce primary airway cultures or brushings of ferrets, pigs, and humans. 33 –35
Apart from rAAV-based gene transfer, also other viral vector systems are currently being investigated to target the airways, such as lentiviral (LV) vectors (reviewed in ref. 36 ). Several envelopes have been used for airway transduction, such as glycoprotein-G from the vesicular stomatitis virus, which requires pretreatment with tight junction openers, 37 as well as other envelopes with natural airway tropism such as baculo- and Sendai virus-derived envelopes (gp64 and F/HN, respectively). 38,39 Similar to what we observe after rAAV-mediated gene transfer, efficient and long-term reporter gene expression in murine airways has been achieved, making LV an alternative candidate for CF gene therapy. 4,19,40 It is yet to be elucidated if sustained LV-mediated gene expression results from transduced long-living terminally differentiated epithelial cells or from vector integration into stem cells. 39 Although an integrating vector is in theory capable of transducing airway stem cells, stem and progenitor cells in an adult organ reside in well-protected niches, making them challenging targets for gene transfer. Second, integration into the host genome comes with the extra safety risks of insertional mutagenesis (reviewed in ref. 41 ), which should be carefully monitored in treated patients.
rAAV administration inevitably induces a systemic humoral immune response against the rAAV vector after a single dose with the induction of capsid-specific nAb (reviewed in ref. 42 ). To achieve lifelong gene correction, even in a low proliferative tissue such as the lung, repeated vector administration will be necessary. However, capsid-specific nAb pose a challenge to successfully readminister an autologous vector. Different strategies are currently being investigated to obtain successful rAAV readministration. One method is to first transduce with empty rAAV particles to deplete the serum or broncheo-alveolar lavage fluid from rAAV-specific nAb. 43 Other strategies focus on serotype switching with a serologically distinct rAAV serotype, 44 immunosuppression, 45 vector administration to an immature immune system, 19,46 or prolonging the time period between vector administrations. 16,17 The latter approach hypothesizes that a decrease in nAb over time will facilitate repeated vector dosing. In our setup, serum nAb levels declined over time, but remained fairly high even at 14 months, when the second dose was administered (Fig. 5). This could explain the less efficient readministration when using the same vector serotype. We, however, show that it is feasible to obtain successful readministration with an autologous vector, resulting in a significant increase in pulmonary gene expression (Fig. 6). Although the average increase in the nasal region was not significant, in three out of five mice, increased gene expression levels were observed, supporting the possibility to readminister rAAV2/5 despite high-level circulating nAb.
Sumner-Jones and colleagues reported that rAAV5/5 readministration 8.5 months after primary vector dose was not successful. 21 In contrast, two studies reported that rAAV2/5 could already be readministered 5–6.5 months after primary airway delivery, despite the presence of serum nAb. 16,17 These conflicting data show that serum nAb might not be the best predictor to determine whether rAAV transduction will be hampered, as this will also depend on the animal species or strain used, the delivery route, target organ, and vector dose.
Taken together, we have demonstrated that rAAV2/5-based gene transfer into mouse airways provides a generic model to study gene therapy for a variety of respiratory diseases in a preclinical context, allowing first proof-of-concept studies to be performed on gene transfer efficacy. However, a comparative analysis in larger animals as well as in relevant human airway cell lines is a prerequisite to confirm these findings, before becoming clinically applicable. In conclusion, the results of this study underscore the therapeutic potential of rAAV2/5 in a preclinical setting and open perspectives to develop a successful gene therapy approach for pulmonary diseases based on a nonintegrating rAAV vector.
Footnotes
Acknowledgments
D.V. is a doctoral fellow supported by a KU Leuven grant, DBOF/10/062. M.S.C. is a postdoctoral fellow supported by the “Fund Alphonse and Jean Forton,” King Baudouin Foundation, Belgium. A.Q.J. is a predoctoral fellow supported by an ERASMUS Student Mobility for Placements grant. Research was funded by IWT-Vlaanderen (SBO Myriad) and by the IMIR program from KU Leuven. We would like to acknowledge Dr. Johan Nuyts for his help with power regression analysis of gene expression kinetics. Finally, we would like to thank Liesbeth De Keersmaecker for excellent technical assistance and acknowledge the Leuven Viral Vector Core (LVVC) for rAAV vector productions performed by Jooke Van der Veken and Sylvie De Swaef.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.