
Abstract
Immune responses to HIV in the vaginal tract effectively trigger both systemic and mucosal protection, providing a double layer of defense. However, recombinant adenoviral (rAd) vectors delivered intravaginally do not effectively penetrate the mucus layer and vaginal epithelium, and instead are rapidly cleared. To overcome these barriers, we previously synthesized a novel cationic polyethylene glycol derivative that can self-assemble into nanocomplexes with rAd. These nanocomplexes can help rAd bypass the mucus layer and enhance mucosal immune response to the encoded antigen. However, the resulting cellular and humoral responses were still lower than those elicited by single intramuscular injection of rAd. Therefore, in the present study we investigated a new vaccination strategy involving intravaginal priming with our nanocomplexes, followed by an intramuscular boost with rAd-gag. Mice immunized in this way showed more potent humoral and cellular responses, as well as higher IgA levels, than animals primed and boosted intravaginally with nanocomplexes. These results show the promise of a prime–boost strategy for developing vaccine candidates against HIV.
Introduction
H
Increasing evidence shows that initiating immune reactions at mucosal sites provides both systemic and mucosal immune responses, leading to a double layer of protection against pathogens. 5,8 However, intravaginal administration is problematic, because the vaginal epithelium and a layer of secreted mucus on top of the vaginal mucosa pose substantial barriers to pathogen entry. 9 The secreted mucus acts as a sieve, preventing passage of larger species (with diameters >1 μm) as well as trapping charged species. These particles are then rapidly eliminated during mucus turnover, 10 which in mice occurs at 1 ml/day at diestrus. 11 This has led researchers to focus on developing effective trans-mucosal vectors.
Recently, Ensign et al. have shown that nanoparticles coated with a high density of low-molecular-weight (2–5k) polyethylene glycol (PEG) can rapidly penetrate human cervicovaginal mucus, presumably because the PEG prevents the particle core from interacting with mucus. 12,13 Similarly, our group synthesized a novel cationic PEG derivative with a wheat-like structure, amino-(EO)n/(AGE)m-Cyss (APCs), which comprises a neutral, linear molecular PEG (molecular-weight is 5k) bearing approximately 10.5 cationic amine pendants (5k-APC). 14 This derivative self-assembles into nanocomplexes with recombinant adenovirus (rAd) (Fig. 1A). The 5k-APC-rAd nanocomplexes transducted VK2/E6E7 and DC 2.4 cells more efficiently than rAd alone and enhanced gene expression in reproductive tissues after intravaginal administration. 14 We also found that 5k-APC-rAd in which the rAd was carrying the HIV gag gene triggered more powerful cellular, humoral, and mucosal immune responses to the gag protein than rAd after intravaginal administration. 14 Nevertheless, cellular and humoral immune responses elicited by intravaginal administration of 5k-APC-rAd were substantially lower than those elicited by a single intramuscular injection of rAd. 14 This led us to question whether we could devise a vaccination protocol for 5k-APC-rAd that would (1) improve its intravaginal delivery, thereby triggering both mucosal and systemic immune responses at the same time, and (2) take advantage of intramuscular injection, which can generate higher-avidity IgG than mucosal administration, as well as larger numbers of polyfunctional CD8+ T lymphocytes, which can migrate to systemic and mucosal compartments. 15 In addition, we wanted a protocol that would (3) avoid the disadvantages of intramuscular immunization, which might lead to production of antiadenovirus antibodies that would prevent 5k-APC-rAd from inducing a robust mucosal response. 16,17
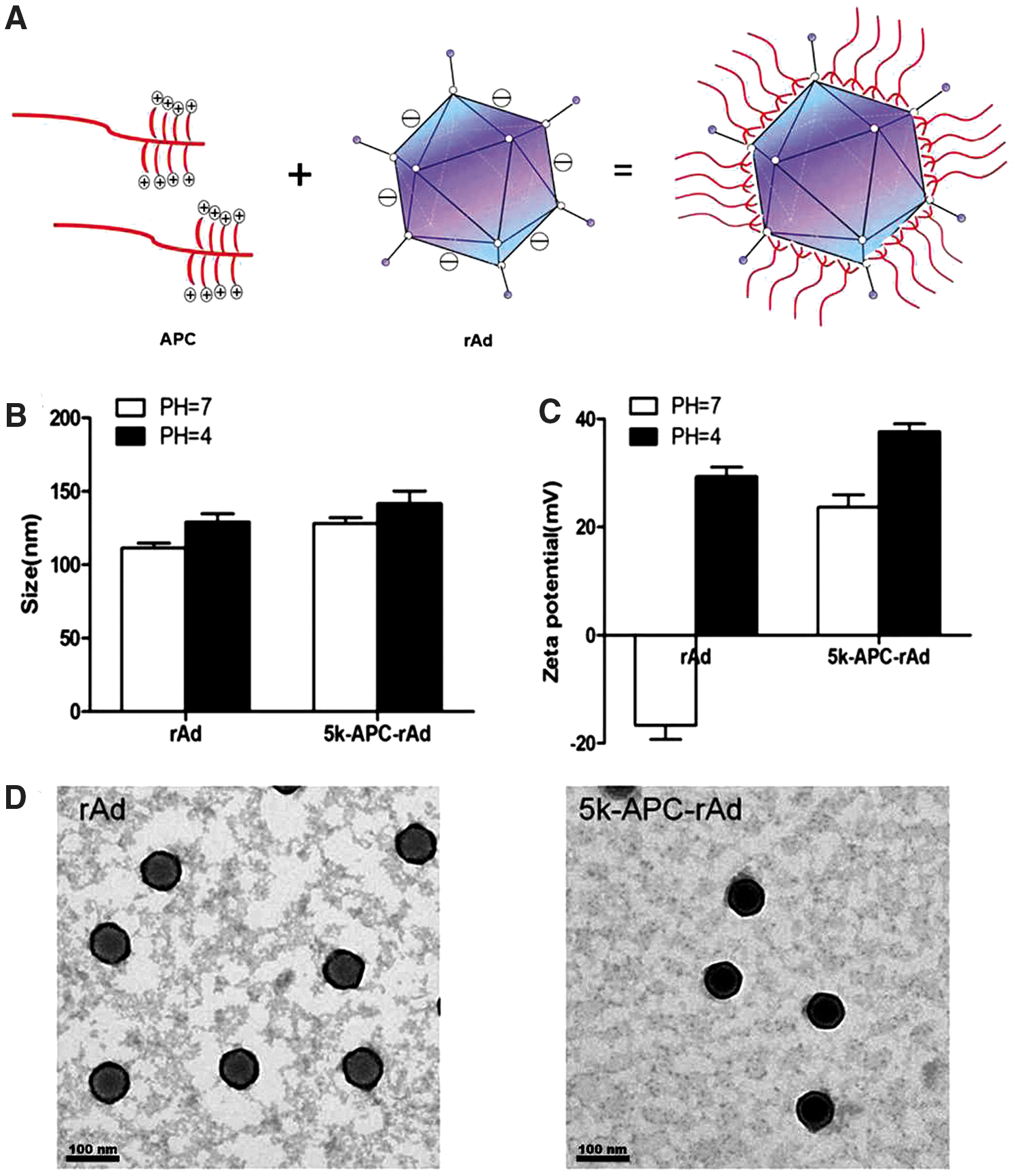
Morphology and biophysical characteristics of 5k-APC-rAd nanocomplexes.
To accomplish these goals, we developed a prime–boost strategy in which 5k-APC-rAd nanocomplexes were first administered intravaginally, followed by intramuscular administration of rAd. We evaluated the prime–boost strategy in vivo using rAd carrying the HIV gag gene, thereby providing a model for HIV vaccination.
Materials and Methods
Materials
Carboxyfluorescein diacetate succinimidyl ester (CFSE) was purchased from Invitrogen. Brefeldin A and the following antimouse antibodies were purchased from eBioscience: FITC-labeled antibodies against CD8a and CD4; PE-labeled antibodies against IFN-γ, IL-4, and CD44; and PE-Cy7-labeled antibodies against CD62L. Goat antimouse horseradish peroxidase (HRP)-conjugated antibodies against IgA, IgG1, and IgG2a were obtained from Santa Cruz, and goat antimouse HRP-conjugated antibody against IgG was obtained from ZSGB-BIO. HIV gag peptide corresponding to the H-2 Kd-restricted immunodominant CTL epitope (AMQMLKETI) was obtained from GenScript. Recombinant human HIV-1 gag p24 was obtained from USBiological (Swampscott). Amino-PEG-poly (AGE-Cys) (APC) was prepared as described. 14
Animals
Specific pathogen-free female BALB/c mice aged 6–8 weeks were purchased from the Laboratory Center of Sichuan University. Animals were housed at constant temperature and humidity under a 12 hr light–dark cycle. All procedures with animals were conducted in accordance with institutional animal care and use guidelines.
rAd vectors
An E1/E3-deleted adenovirus (serotype 5)-derived vector encoding HIV-1 gag p24 was kindly provided by Prof. Hildegund C.J. Ertl (Wistar Institute of Anatomy and Biology, Philadelphia, PA). Adenovirus was amplified in the 293T cell line and purified though CsCl gradient centrifugation following standard methods. Virus particle number was determined using ultraviolet absorption at 260 nm, and infectious titer was determined by titration on 293T cells. All virus stocks were stored at −80°C before use.
Preparation of rAd and cationic polymer nanocomplexes
The PEG derivative 5k-APC was prepared as described. 14 The derivative solution and rAd solution were adjusted to the desired concentrations using 5% glucose, and then the rAd solution was added to the polymer solution. The mixture was vortexed for 10 sec and allowed to stand at room temperature for 30 min.
Characterization of rAd and cationic polymer nanocomplexes
Samples (0.2 ml) of rAd or 5k-APC-rAd nanocomplexes were diluted with ultrapure water (pH 7) or lactic acid solution (pH 4) to a final volume of 1 ml. Mean particle size and zeta potential were determined at 25°C using photon correlation spectroscopy (Malvern Zetasizer Nano ZS90; Malvern Instruments) with a 50 mV laser. Measurements were performed in triplicate over at least 20 runs in each experiment.
The morphology of rAd and nanocomplexes was observed by transmission electron microscopy (Tecnai G 2 F-20; FEI). Each sample was processed by the negative stain method. Freshly prepared samples were allowed to absorb into copper grids for 5 min. Then bulk solution was removed and the samples were stained with 1% (w/v) aqueous uranyl acetate for 2 min. Excess dye was removed with filter paper, and samples were air-dried and then analyzed.
Mouse immunization
We compared the following formulations and routes of administration: (1) a single dose of rAd delivered intramuscularly; (2) rAd delivered intravaginally as a prime, with rAd delivered intravaginally as a boost; (3) rAd delivered intravaginally as a prime, with rAd delivered intramuscularly as a boost; (4) 5k-APC-rAd delivered intravaginally as a prime, with 5k-APC-rAd delivered intravaginally as a boost; and (5) 5k-APC-rAd delivered intravaginally as a prime, with rAd delivered intramuscularly as a boost. In all cases, female BALB/c mice were given medroxyprogesterone acetate (day 0), and then anesthetized 5 days later.
For intravaginal administration (i.vag.), rAd or 5k-APC-rAd nanocomplexes suspended in 5% glucose were pipetted into the lumen (20 μl/animal, 2 × 1010 viral particles). Mice were kept positioned with the vagina facing upward for more than 40 min to reduce leakage. For intramuscular administration (i.m.), rAd (2 × 109 viral particles) was injected into the lower leg muscle. Primes were administered on day 5, and boosts on day 19.
Mice were sacrificed 2 weeks after boosts and analyzed in an in vivo cytotoxic T lymphocyte (CTL) assay, intracellular cytokine staining (ICS) assay, enzyme-linked immunosorbent assay (ELISA), and enzyme-linked immunospot assay (ELISPOT). Mice were sacrificed 40 days after boosts for analysis in assays of memory T-cells.
CTL assay
Spleens were extracted by sterile dissection. Single-cell lymphocyte suspensions were prepared by grinding. ACK lysis solution (0.15 M NH4Cl, 10.0 mM KHCO3, 0.1 mM EDTA, pH 7.4) was added to lyse red blood cells. Half of the lymphocytes were pulsed with 2 μm HIV gag peptide for 1 hr at 37°C and then labeled with 4 μm CFSEhigh for 10 min. The other half were incubated with the medium for 1 hr at 37°C and then stained with 0.4 μm CFSElow for 10 min. CFSE labeling was quenched by addition of fetal bovine serum (FBS) to a final concentration of 20% (v/v). Equal numbers of cells from the two aliquots were pooled and injected into immunized mice or naive mice via the tail vein. At 18 hr after injection, splenocytes from recipient mice were harvested and analyzed by flow cytometry (Beckman Coulter).
Percentage of specific lysis was determined as follows: specific lysis (%) = 100 × [1 − (ratio of CFSElow/CFSEhigh cells recovered from naive mice/ratio of CFSElow/CFSEhigh cells recovered from immunized mice)].
IFN-γ ELISPOT assay
The mouse IFN-precoated ELISPOT kit (Dakeve) was used. The plate was blocked for 10 min at room temperature with RPMI 1640 medium. Spleens harvested from immunized mice were grinded in mouse lymphocyte separation medium (Dakeve). Splenocyte suspensions were transferred to centrifuge tube, and 200 μl RPMI 1640 medium was added. After gradient centrifugation, lymphocytes were harvested. Then lymphocytes were pipetted into strips. Cells were stimulated with HIV gag peptide (10 μg/ml) or medium alone. PHA was added as a positive control as directed by the manufacturer. After 18 hr incubation at 37°C, we decanted solution from wells, and lysed cells at 4°C by adding cold demineralized water. After 10 min, solution was discarded from wells and the wells washed 5 times with washing buffer. Then 100 μl of the appropriate dilution of biotinylated detection antibodies was added to each well, and the plate was incubated for 1 hr at 37°C. Strips were washed with washing buffer, and 200 μl of the appropriate dilution of streptavidin-HRP solution was added to each well. After another 1 hr incubation at 37°C, AEC substrate solution was added to each well, and the plate was incubated for 25 min at room temperature in the dark. Spot development was halted after 10 min by washing both sides of the PVDF membrane with demineralized water. Numbers of IFN-γ spot-forming cells (SFCs) were determined using a plate reader.
ICS assay
Spleens were extracted by sterile dissection. Single-cell lymphocyte suspensions were harvested after red blood cells were lysed using ACK lysis solution. Lymphocytes were seeded into 96-well plates and pulsed (or not) with 2 μg/ml HIV gag peptide for 1 hr at 37°C. Brefeldin A was added to all wells, and plates were incubated another 4 hr. Lymphocytes were resuspended in 100 μl staining buffer and incubated in the dark for 30 min at 4°C with FITC-conjugated antimouse CD8a or CD4 antibodies (1:100 dilution). Cells were fixed with 100 μl IC fixation buffer (eBioscience), followed by 1× permeabilization buffer (eBioscence); then the suspensions previously stained with anti-CD8a were stained for 20 min with PE-conjugated antimouse IFN-γ antibody (1:100), whereas the suspensions previously stained with anti-CD4 were stained for 20 min with PE-conjugated antimouse IL-4 antibody (1:100). Cells were washed and analyzed by two-color flow cytometry (Beckman Coulter).
Anti-HIV gag antibody response in serum and vaginal washes
Serum and vaginal wash samples were assayed for IgA and IgG using ELISA. The 96-well plates (Corning) were coated with 1 μg/ml HIV-1 gag/p24 protein in coating buffer and incubated at 4°C overnight. After blocking, mouse serum (1:128 dilution) or vaginal washes (1:2 dilution) were added, and plates were incubated for 2 hr. Plates were washed and incubated for 1 hr at 37°C with HRP-conjugated antimouse antibodies against IgG (1:10,000) or IgA (1:5000). Unbound conjugates were washed away with phosphate-buffered saline (PBS) containing 0.05% Tween-20, tetramethylbenzidine substrate was added, and plates were incubated at room temperature for 30 min. Color development was halted by adding 2 M H2SO4. Optical density (OD) values were measured at 450 nm using an ELISA reader (VarioSkan; Thermo Fisher Scientific).
Memory T-cell assay
Immunized mice were sacrificed 40 days after boost. Single-splenocyte suspensions were prepared as described above. After red blood cell lysis, splenocytes were washed with PBS and stained with FITC-conjugated antimouse CD8 antibody, PE-conjugated anti-CD44 antibody, and PE-Cy7-conjugated anti-CD62L antibody for 30 min at 4°C in the dark. Splenocytes were washed, diluted in PBS, and analyzed by three-color flow cytometry (Beckman Coulter).
Data analysis
All data were reported as mean ± standard deviation (SD), and experiments were conducted at least in triplicate. Measurements from two groups were compared using the Student's t-test, whereas measurements from more than two groups were compared using one-way ANOVA. The threshold of significance was defined as p < 0.05.
Results
Size, zeta potential, and morphology of complexes
Particle size and zeta potential of rAd and nanocomplexes were analyzed in both neutral and acidic media (Fig. 1B and C). This is because the mouse vaginal mucus exhibits a nearly neutral pH, whereas the human vaginal mucus exhibits a pH ∼4. Under neutral conditions, the size of 5k-APC-rAd (128 ± 7 nm) and that of rAd (111 ± 6.3 nm) were similar, and their sizes did not change substantially under acidic conditions. Whereas rAd particles were negatively charged at pH 7, 5k-APC-rAd complexes were positively charged.
Transmission electron micrographs showed the expected morphology for rAd and nanocomplexes (Fig. 1D). Nanocomplexes clearly showed shell-core structures, confirming the idea that polymers coated the rAd capsid in an outer layer that was anchored to the core through electrostatic interactions.
In vivo CTL assay
We performed in vivo CTL assays to determine whether our prime–boost strategy induces a stronger antigen-specific CD8+ CTL response. CFSEhigh and CFSElow peak heights were similar for naive mice (Fig. 2A). To quantitate immune responses, we determined the percent change in the number of HIV gag-pulsed CFSEhigh splenocytes relative to the number of nonpulsed CFSElow splenocytes (Fig. 2A and B). Antigen-specific CD8+ T-cell response in mice intravaginally primed with 5k-APC-rAd was significantly higher after intramuscular boost with rAd (93.88%) than after intravaginal boost with 5k-APC-rAd (36.42%, p < 0.001; Fig. 2A and B). Animals primed intravaginally with rAd and boosted intramuscularly with rAd showed 78.17% specific lysis, whereas animals primed and boosted intravaginally with rAd showed only 11.91% specific lysis (p < 0.001; Fig. 2A and B).
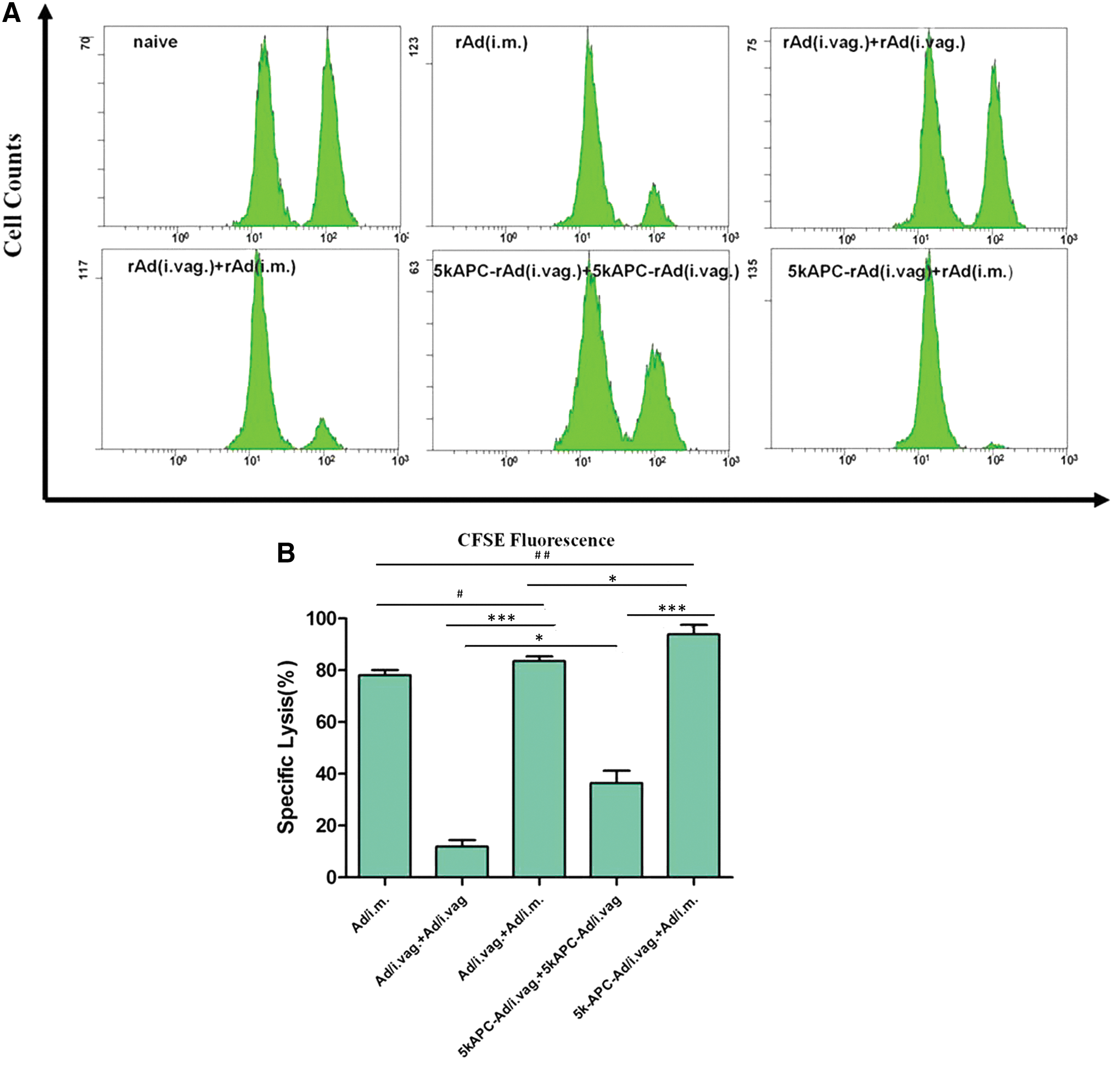
HIV gag-specific cytolytic T-cell response elicited by rAd carrying the HIV gag gene. BALB/c mice were primed intravaginally with 2 × 1010 viral particles (vp) of 5k-APC-rAd and boosted either intravaginally with 2 × 1010 vp of 5k-APC-rAd or intramuscularly with 2 × 109 vp of rAd. BALB/c mice were primed intravaginally with 2 × 1010 viral particles (vp) of rAd and boosted either intravaginally with 2 × 1010 vp of rAd or intramuscularly with 2 × 109 vp of rAd. Another group of animals received a single intramuscular injection of 2 × 109 vp of rAd. At 2 weeks postimmunization, equal numbers of HIV gag-pulsed CFSEhigh and nonpulsed CFSElow splenocytes from naive mice were injected into the immunized animals via the tail vein. At 18 hr later, splenocytes from recipient mice were harvested and analyzed by flow cytometry.
HIV gag peptide-specific CD8+ and CD4+ T-cell responses
The ICS assay was used to quantitate IFN-γ production by CD8+ T-cells and IL-4 secretion by CD4+ T-cells (Fig. 3A and B). Immune responses were analyzed by flow cytometry. In animals primed intravaginally with 5k-APC-rAd, the percentage of CD8+ T-cells producing IFN-γ in response to HIV gag was significantly higher after an intramuscular boost with rAd (1.18%) than after an intravaginal boost with 5k-APC-rAd (0.88%, p < 0.01; Fig. 3A and C). The corresponding result in mice that received a single intramuscular injection of rAd was 0.83%.

Antigen-induced IFN-γ production by CD8+ T-cells and IL-4 secretion by CD4+ T-cells.
In animals primed intravaginally with 5k-APC-rAd, the percentage of CD4+ T-cells secreting IL-4 in response to HIV gag was similar after an intramuscular boost with rAd (0.84%) or an intravaginal boost with 5k-APC-rAd (0.85%; Fig. 3B and C). These results are consistent with other experiments in which single intramuscular injection of rAd elicited a similar CD4+ T-cell response as a single intravaginal administration of 5k-APC-rAd (data not shown).
IFN-γ ELISPOT assay
Numbers of IFN-γ-secreting cells in the IFN-γ ELISPOT assay were higher after intravaginal prime with 5k-APC-rAd and intramuscular boost with rAd (1197 SFCs/106 cells) than after intravaginal prime and boost with 5k-APC-rAd (227 SFCs/106 cells; Fig. 4A and B). This is consistent with results of the in vivo CTL assay. Among animals that received an intravaginal prime with rAd, numbers were higher after an intramuscular boost with rAd (850 SFCs/106 cells) than after intravaginal boost with rAd (142 SFCs/106 cells, p < 0.5; Fig. 4A and B).
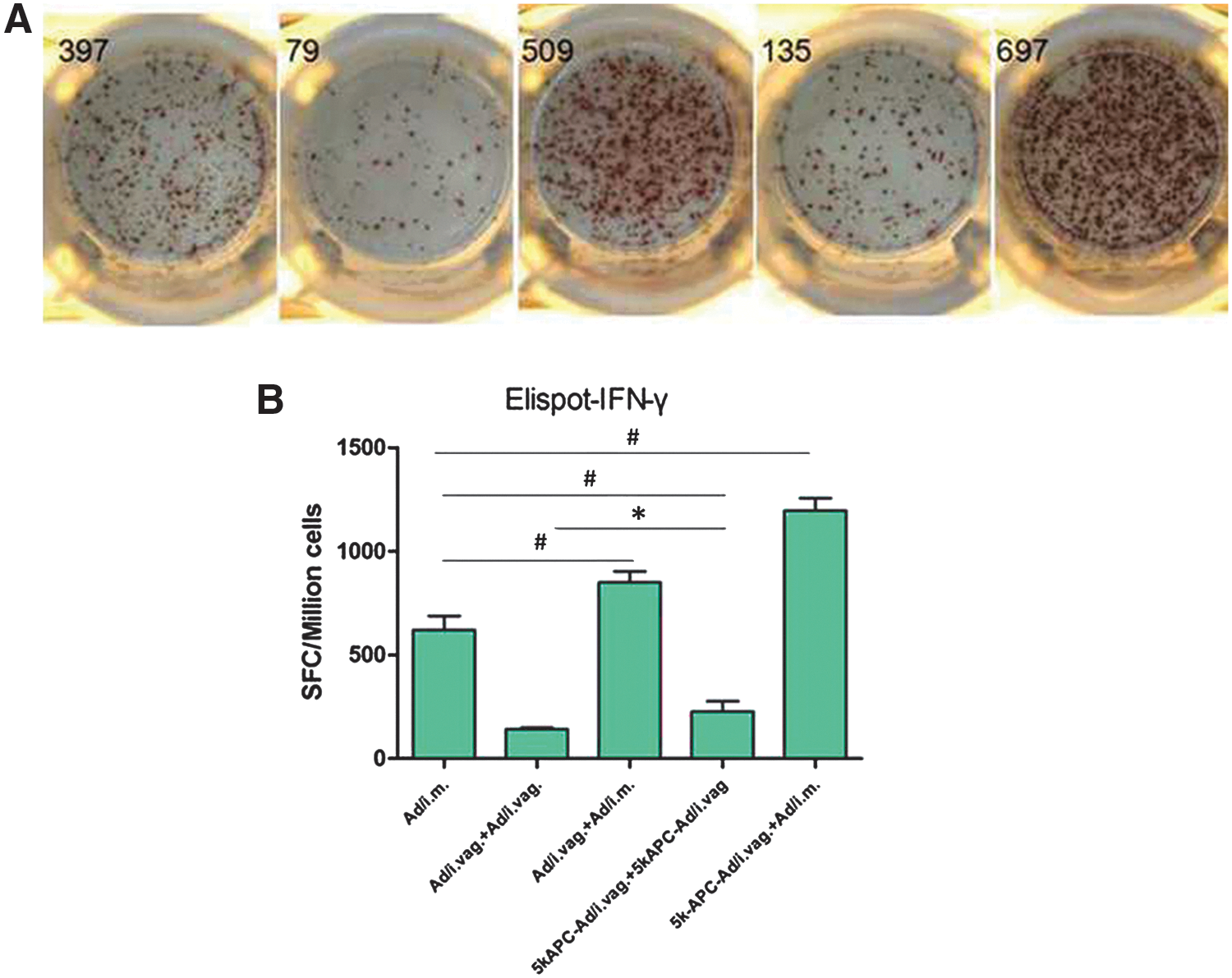
Detection of IFN-γ-secreting cells in the spleens of mice immunized using rAd carrying the HIV gag gene.
HIV gag peptide-specific antibody response in serum and vaginal washes
Efficacy of most mucosal vaccines correlates well with levels of secretory immunoglobulin A (sIgA). 18 Therefore, we harvested vaginal lavage fluid and assayed local secretion of IgA using ELISA. Concentrations of IgA in vaginal washes were similar between mice primed intravaginally with 5k-APC-rAd and boosted intramuscularly with rAd (OD = 0.44) and mice primed and boosted intravaginally with 5k-APC-rAd (OD = 0.46). Levels of IgA were higher in mice primed intravaginally with rAd and boosted intramuscularly with rAd (OD = 0.30) than in mice that received a single intramuscular injection of rAd (OD = 0.17). In fact, IgA concentration among all treatment groups was lowest in animals that received a single intramuscular injection of rAd.
To assess whether our prime–boost strategy could evoke robust HIV gag-specific antibody in serum, IgG levels were assayed using ELISA. Examination of serum sampled 14 days after boosting showed that mice primed intravaginally with 5k-APC-rAd and boosted intramuscularly with rAd showed higher levels of antigen-specific IgG (OD = 1.15) than mice primed and boosted intravaginally with 5k-APC-rAd (OD = 0.84, p < 0.001; Fig. 5B). Levels of IgG were also higher in animals primed intravaginally with rAd and boosted intramuscularly with rAd (OD = 0.97). Levels in mice primed and boosted intravaginally with 5k-APC-rAd (OD = 0.84) were similar to those in mice that received a single intramuscular injection of rAd (OD = 0.83; Fig. 5B).

IgA levels in vaginal mucosa and IgG levels in serum after immunization with rAd carrying the HIV gag gene.
Memory T-cell assay
Eliminating viral infection may require inducing a persistent memory CD8+ T-cell response. Two subsets of memory T-cells mediate this response. 19 Effector memory T (TEM) cells act as the first line of defense, providing immediate protection at environmental barriers, whereas central memory T (TCM) cells act as the second line of defense, “recalling” past experience with the antigen and triggering T-cell proliferation to mount a response. 20 TCM cells respond more vigorously to secondary challenge than TEM cells do, 19 and so we assessed the ability of our prime–boost immunization protocol to increase numbers of CD8+ TCM cells (CD8+ CD44+ CD62L+). The percentage of CD8+ TCM cells was highest after intravaginal prime with 5k-APC-rAd and intramuscular boost with rAd (2.75%), and lower after intravaginal prime and boost with 5k-APC-rAd (1.93%) or after single intramuscular injection with rAd (1.92%; Fig. 6).
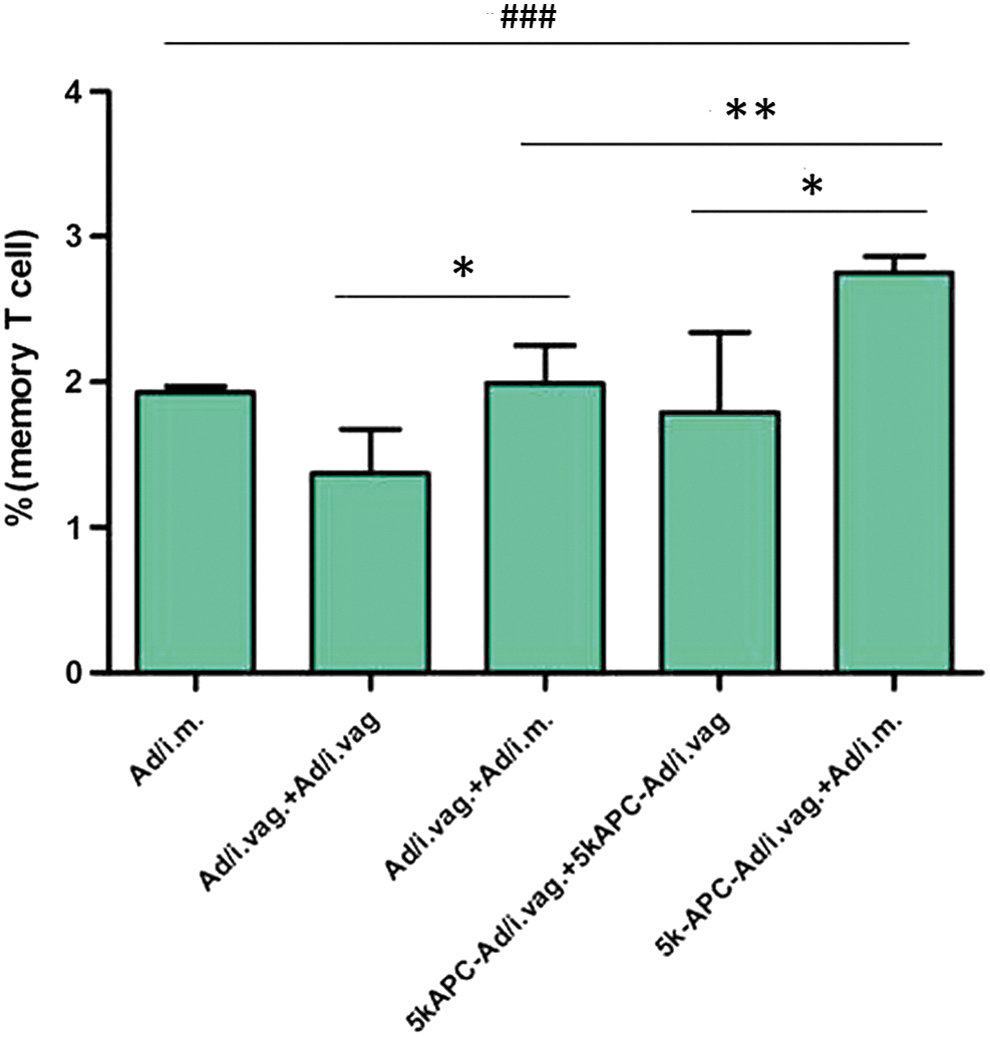
Memory T-cell response after immunization with rAd carrying the HIV gag gene. Mice were sacrificed at 40 days after boost. Single-splenocyte suspensions were prepared and stained with FITC-conjugated antimouse CD8 antibody, PE-conjugated anti-CD44 antibody, and PE-Cy7-conjugated anti-CD62L antibody (eBioscience) at 4°C for 30 min in the dark. Memory T-cell response was measured using flow cytometry. Data are mean ± SD (n = 4). **p < 0.01, *p < 0.05 (vs. i.vag+i.m. and i.vag.+i.vag.) and ###
p < 0.001 (vs. i.m.). Color images available online at
Discussion
A broader understanding of mucosal immunology and different routes of vaccine administration and immune responses is urgently needed. Mucosal administration is critical for effective vaccination against HIV-1 because it can simultaneously elicit systemic and mucosal immune responses. In a previous study, we examined specially modified rAd particles capable of entering the vaginal mucosa efficiently. We wanted to improve immune response to mucosal vaccination, given that immunogenic antigen doses are usually higher for mucosal administration than via other parenteral routes. 21 In the present study, we sought to strengthen anti-HIV-1 humoral and cellular and mucosal responses at the same time, and so we tested various prime–boost combinations involving different rAd-based formulations and delivery routes. Our present results suggest that intravaginal prime with modified rAd particles followed by intramuscular boost with rAd can elicit strong systemic and mucosal responses, including persistent stimulation of memory T-cells. These effects are stronger than those observed with single intravaginal or single intramuscular administration with rAd.
We chose to work with rAd because it is relatively easy to produce, safe, and able to elicit broad immune responses. 22 Indeed, rAd shows excellent potential as a vaccine delivery vector against influenza virus H5N1, 23 Ebola virus, 24,25 and hepatitis B virus. 26 Whereas the negatively charged rAd in the neutral medium turned into positively charged in the acidity medium, which could be trapped by the human vaginal mucus (negatively charged, pH 4) through electrostatic adhesion, 14 5k-APC-rAd nanocomplexes are positively charged in both the neutral and acidity medium, but the PEG coating could provide the particles with hydrophilic surface and thus avoid mucus' hydrophobic entrapment, 10 and increase the gene expression and the immune response.
Prime–boost approaches via different administration routes can elicit cellular and humoral immune responses that are comparable or superior to single intramuscular injection with rAd in terms of magnitude, quality, and localization. Intramuscular Ad injection can trigger high production of neutralizing antibodies, 16,17 and such preexisting immunity can pose a boost problem for Ad-based vaccines, because the host produces anti-Ad antibodies that can inactivate the rAd vaccine vehicle. To reduce this problem in our case, we primed first with the modified rAd particles, which were optimized for mucosal entry, and then later administered rAd intramuscularly.
Our approach of intravaginal prime with 5k-APC-rAd followed by intramuscular rAd boost generated the strongest T-cell responses among all the vaccination regimes that we tested. T-cells play a major role in controlling chronic viral infection, 27,28 and viral infection triggers vigorous expansion of antigen-specific CD8+ T-cells, which eliminate pathogens by directly killing infected cells or by secreting cyto- or chemokines to trigger further responses. 20 Using the in vivo CTL assay, we showed that our prime–boost regimen elicited the most potent cytolytic activity in this assay. This likely reflects the ability of intramuscular rAd injection to trigger a strong CD8+ T-cell response.
One of the cytokines secreted by CD8+ T-cells to recruit further responses is IFN-γ, which serves as a Th1 marker. 29 Therefore, we examined our prime–boost approach in ICS and IFN-γ ELISPOT assays to examine proportions of IFN-γ-producing T-cells. Both assays showed that our prime–boost approach elicited the strongest IFN-γ response. CD4+ T-cells can help induce and maintain CD8+ T-cell responses and B cell responses, and IL-4 produced by CD4+ T-cells serves as a Th2 marker of humoral immune response. 30 Therefore, we used the ICS assay to determine proportions of IL-4-secreting CD4+ T-cells for quantifying the specific CD4+ T-cell response. Our prime–boost approach again proved to be the best at inducing CD4+ T-cell secretion of IL-4. In contrast, single intramuscular rAd administration showed the lowest percent of CD4+ T cells positive for IL-4.
Vaccines against viruses such as HIV that enter so efficiently through mucosa should strongly induce production of sIgA. This antibody prevents mucosal infection by trapping the virus and blocking interactions between molecules on the viral surface and the target epithelium. Our prime–boost approach of intravaginal prime with 5k-APC-rAd followed by intramuscular rAd boost induced higher production of anti-HIV gag sIgA in the vaginal cavity than did intravaginal prime and boost with 5k-APC-rAd. Ideally, an anti-HIV vaccine should also trigger higher levels of IgG in serum. We observed significant IgG levels in serum after priming with either rAd or 5k-APC-rAd, as long as intramuscular boost with rAd followed. Even after boosting with 5k-APC-rAd by the vaginal route, 5k-APC-rAd prime still evoked higher IgG than a single intramuscular immunization with rAd.
Taken together, these experiments suggest that our prime–boost approach can trigger a strong humoral and cell-mediated immune response in the short-term. As a first step toward examining whether our approach can induce long-lasting protection, we measured levels of TCM cells. Immune memory involves both TCM cells and TEM cells, which differ in their surface expression of CCR7 and CD62L and in their migration patterns: TCM cells tend to circulate though lymphoid organs; TEM cells, though multiple peripheral tissues. 31,32 We focused on TCM cells because of their importance for inducing pathogen-specific CTL responses in the long-term. Our prime–boost strategy generated the highest proportion of TCM cells of all vaccination regimes we tested.
Conclusions
Using 5k-APC as a vaccine carrier and a prime–boost strategy involving 5k-APC-rAd nanocomplexes and rAd, we effectively induced cellular, humoral, and mucosal immune responses to the HIV gag antigen. Our approach led to higher CD8+ and CD4+ T-cell responses and cytokine production (IFN-γ, IL-4) than several other vaccination regimes that we tested. Our approach also induced higher levels of HIV gag-specific IgG and IgA, and higher percentage of CD8+ TCM cells. These results justify future studies to develop a homologous prime–boost strategy to prevent HIV-1. They also suggest the potential of our approach for delivering other genes useful for vaccination or gene therapy.
Footnotes
Acknowledgments
We are grateful to the National Natural Science Foundation of China (No. 81173011 & 81422044) and the National Science & Technology Major Project of China (No. 2012ZX09304004) for the financial support. We thank Prof. Hildegund C.J. Ertl from the Wistar Institute of Anatomy and Biology for kindly providing the replication-defective adenovirus 5 HIV-gag vector.
Author Disclosure
No competing financial interests exist.