
Abstract
Insufficient local levels of neurotrophic factor after spinal cord injury (SCI) are the leading cause of secondary injury and limited axonal regeneration. Neuritin belongs to a family of neurotrophic factors that promote neurite outgrowth, maintain neuronal survival, and provide a favorable microenvironment for the regeneration and repair of nerve cells after injury. However, it is not known whether the exogenously applied neuritin protein has a positive effect on nerve repair after SCI. This was investigated in the present study using purified human recombinant neuritin expressed in and purified from Pichia pastoris, which was tested in a rat SCI model. A recombinant neuritin concentration of 60 μg/ml induced the recovery of hind limb motor function and stimulated nerve regeneration in rats with SCI. Continuous administration of neuritin at this dose at an early stage after SCI inhibited poly ADP ribose polymerase (PARP) protein degradation and decreased neuronal apoptosis. In addition, during the critical postinjury period of axonal regeneration, exogenous neuritin treatment increased the expression of neurofilament 200 and growth-associated protein 43 in the damaged tissue, which was associated with the restoration of hind limb movement. These results suggest that neuritin creates an environment that promotes nerve cell survival and neurite regeneration after SCI, which contribute to nerve regeneration and the recovery of motor function.
Introduction
A
Neuritin, also known as candidate plasticity-related gene 15, is a neurotrophic factor that promotes neurite growth, synapse formation and maturation, 9 and neuronal migration while inhibiting neuronal apoptosis. 10 –14 As such, neuritin is a positive regulatory factor in neural development and nerve regeneration after injury. Neuritin gene expression was found to be downregulated 7 days after SCI in rats and returned to normal levels at 14 days. 15 Neuritin is coexpressed with microtubule-associated protein 1A, attractin, and myelin oligodendrocyte protein in postmitotic neurons, and these proteins collectively promote neurite growth. Given that the time course of changes in neuritin gene expression mirrors that of functional recovery after SCI, it is hypothesized that neuritin provides a microenvironment that is conducive to this process. We previously demonstrated that recombinant neuritin promotes neurite outgrowth in dorsal root ganglia as well as in PC12 cells in a dose-dependent manner. 16 Silencing neuritin gene expression inhibits axonal growth induced by NGF without affecting neuropathic pain caused by NGF signaling. 17 However, whether exogenously applied neuritin can promote nerve regeneration and the effective dose that is required for this effect are still not known.
In the present study, we purified biologically active recombinant human neuritin and tested its effectiveness in restoring nerve function in a rat model of SCI. We found that treatment with exogenous recombinant neuritin inhibited caspase-3 activation and poly ADP ribose polymerase (PARP) degradation during early stages of SCI while suppressing apoptosis of damaged nerve cells. Moreover, neuritin induced axonal regeneration in remaining neurons, which was associated with a restoration of motor function in injured animals.
Materials and Methods
Animals and groups
Female and male Wistar rats (n = 102 each; 8–9 weeks old; ∼250 g) were supplied by the Experimental Animal Center of Xinjiang Medical University. Animals were divided into sham-operated control (n = 6) and acute SCI (n = 96) groups. To establish optimal neuritin dosage, SCI rats were divided into a His tag (n = 6) and 30, 60, and 90 μg/ml neuritin (n = 6 each) groups. To test the effects of neuritin, SCI rats were divided into His tag (n = 36) and neuritin (n = 36) groups, each with 6 subgroups (analyzed at 6 hr and 3, 7, 14, 28, and 56 days after SCI). Experiments were carried out in accordance with the Ethics Guidelines for Animal Experiments of the First Affiliated Hospital of Shihezi University Medical College, Xinjiang Province, China.
Construction of the pPIC9K-neuritin transfer vector
The strategy for cloning recombinant human neuritin is outlined in Fig. 1A (“pPIC9K transfer vector”). The fragment encoding full-length human neuritin was amplified from a previously constructed pBS-neuritin vector (generated by cloning the full-length cDNA encoding human neuritin from a 293T cell mRNA library and inserting the fragment into the pBS vector) using primers containing EcoRI and NotI restriction sites and six additional histidine residues (6×His tag). Forward and reverse primers were as follows: P1, 5′-GAC
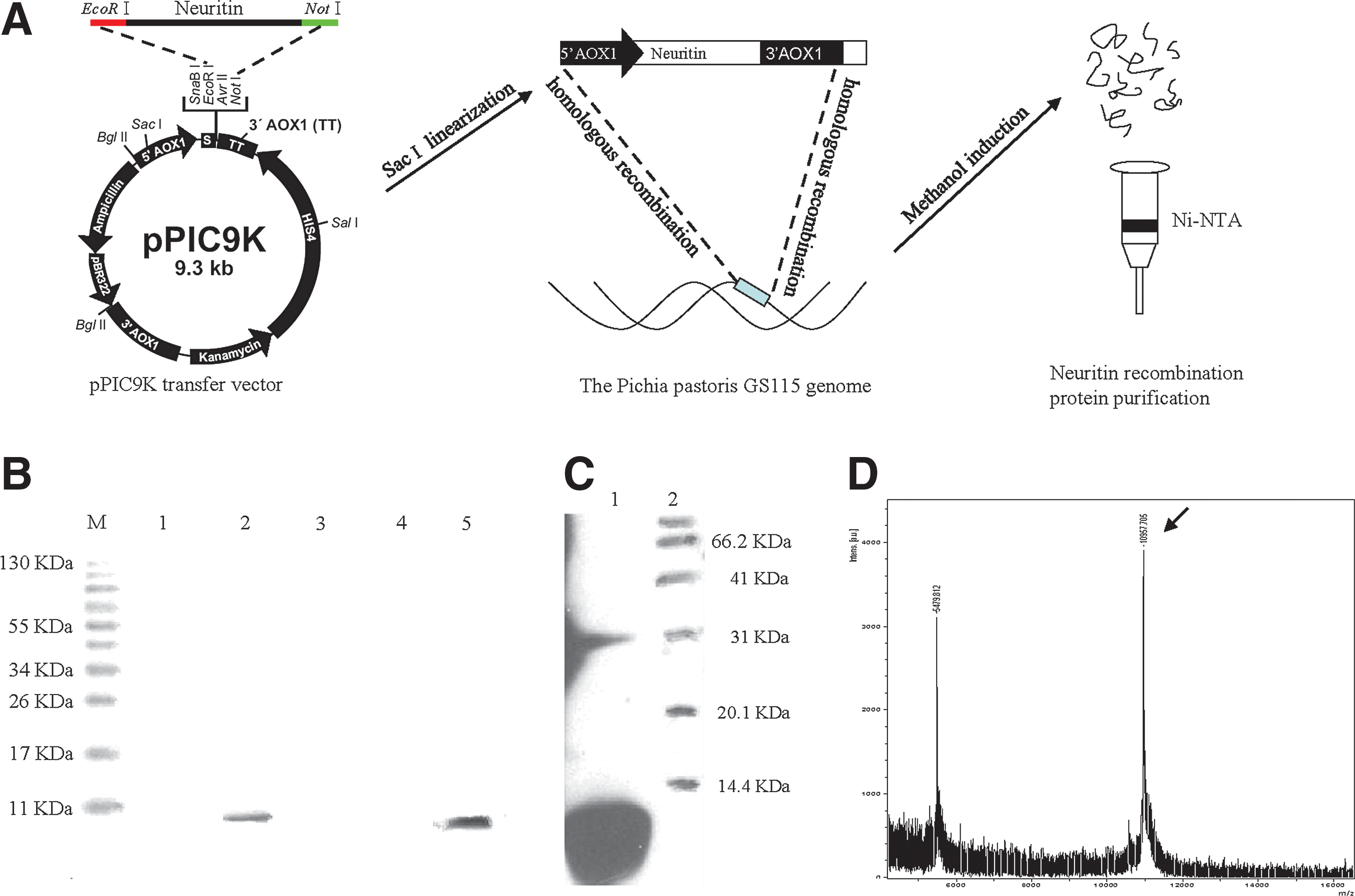
Expression of recombinant human neuritin in Pichia pastoris.
Generation and purification of recombinant human neuritin from neuritin-positive recombinant Pichia pastoris
The pPIC9K-neuritin vector (5–10 μg) was linearized by digestion with SacI and mixed with 100 μl competent P. pastoris GS115 in a 0.2 cm electric rotor cup. The cells were electroporated at 1487 V and 5 μF for 4.7 ms, and after adding 1 ml freezing sorbitol the cells were placed in an ice bath for 30 min. A 300 μl volume of yeast cell suspension was plated onto a minimal dextrose plate and cultured at 30°C until colonies appeared. Each colony was cultured in 200 μg yeast extract peptone dextrose (YPD) medium in a 96-well plate at 30°C for 2 days. A 10 μl suspension mixed with 0.25, 0.5, 1.0, 1.5, 2.0, 3.0, or 4.0 mg/ml G418 was plated onto YPD agar and incubated at 30°C for 2–5 days. Positive recombinant clones were identified by PCR screening.
Expression of recombinant neuritin
Positive P. pastoris clones were grown on yeast medium, and 1% methanol was used for induction at 30°C for 96 hr. After centrifugation, the protein was purified from the supernatant using the Ni-NAT purification kit (Novagen, Madison, WI), dialyzed, an analyzed by Western blotting and mass spectrometry to confirm its identity as human neuritin.
SCI model
Rats were anesthetized with 10% chloral hydrate (300 mg/kg). After making a medial back incision, the T8–T12 thoracic vertebrae were exposed, and the T10 spinous process and vertebral plate were removed with bone forceps. Injury was inflicted using the weight drop method, in which a 15 g weight was dropped from a height of 20 cm onto the SC. 18 A sterile warm saline solution was used to rinse the SC; 30 min after the injury, a 0.1 mm homemade silicone hose was attached to the T12 SC segment along with a heparin cap to cover the catheter tip, which served as a sterile delivery channel into the subarachnoid space. A gelatin sponge was applied to the injured region and the tissue and skin were sutured. Sham-operated control rats were treated in the same manner, expect that the weight was not dropped onto the SC.
Determination of effective neuritin dose
The purified recombinant neuritin protein was diluted with saline to 30, 60, and 90 μg/ml, whereas the His tag was diluted to 60 μg/ml with saline. Rats were injected daily with 100 μl of the protein or His tag solution through the catheter tube, which was removed after 1 week. Animals in the sham group were left untreated. At 6 hr and 3, 7, 14, 28, and 56 days after SCI, rats were sacrificed for analyses.
Analysis of hind limb motor function
The Basso, Beattie, and Bresnahan (BBB) scale was used to evaluate hind limb motor function as previously described. 19 Briefly, rats were placed in an open space with dimensions of 75 cm × 125 cm, and the double-blind method was used to score motor function of the two hind legs.
Histology and immunohistochemistry
Animals were sacrificed by deep anesthetization with chloral hydrate (600 mg/kg by intraperitoneal injection) and transcardial perfusion was carried out with saline followed by 4% paraformaldehyde. A 1.5 cm section of tissue from the T-10 section of the SC—including 0.5 cm above and below the injury site—was removed from six injured rats (three each from the neuritin and His group) and embedded in paraffin. Serial sections 6 μm thick were cut and stained with hematoxylin and eosin (H&E). Sections were then washed in phosphate-buffered saline (PBS), incubated in 1% H2O2 in PBS for 10 min to quench endogenous peroxidase, and boiled for 10 min in 0.01 M citric acid for antigen retrieval. After blocking for 20 min in a solution of 5% normal goat serum, 0.5% Triton X-100, and 0.2% bovine serum albumin in PBS, sections were incubated overnight at 4°C with mouse monoclonal antineurofilament (NF)-200 (1:500) or mouse monoclonal anti-growth-associated protein (GAP) 43 (1:500) antibody (both from Abcam, Cambridge, MA). Horseradish peroxidase-conjugated goat antimouse IgG (1:500; Bio-Rad, Hercules, CA) was used as a secondary antibody. The primary antibody was omitted for control sections. Diaminobenzidine (Sigma-Aldrich, St. Louis., MO) was used for signal detection. Tissue sections from three rats in each group were used for measurements, two high-quality seections per rat were selected for analysis, and four randomly selected fields (the ventral gray matter) per slide were imaged with a 3D de-convolution microscope (Zeiss, Jena, Germany), and Image-Pro Plus software (Media Cybernetics, Rockville, MD) was used to determine the cumulative integral optical density (IOD), from which the mean density (IOD/area) was calculated.
Western blotting
A 4 cm segment of injured SC tissue was excised from three rats and flash-frozen in liquid nitrogen. The frozen tissue was ground, mixed with radioimmunoprecipitation assay (RIPA) lysis buffer containing proteinase inhibitor [50 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 1 mM EDTA, 0.1 mM Na3VO4, 25 mM NaF, 10 mg/ml aprotinin, 10 mg/ml leupeptin, 1 mM phenylmethylsulfonyl fluoride] (Beyotime, Jaingsu, China) for 30 min on an ice bath, and centrifuged at 4°C and 12,000× g for 10 min. The protein concentration in the supernatant was measured with the Bradford assay, and 100 μg total protein was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes, which were probed with the following primary antibodies: mouse monoclonal anti-neuritin 1 (1:200; R&D Systems, Minneapolis, MN), rabbit monoclonal anti-PARP (1:1000; Cell Signaling Technology, Danvers, MA), mouse monoclonal anti-NF-200 (1:1000) and anti-GAP-43 (1:1200) (both from Abcam), and mouse monoclonal anti-β-actin (1:4000, A1978; Sigma-Aldrich).
Statistical analysis
Data are presented as mean ± SD. Experiments were repeated three times and assays were performed in triplicate. Differences between two groups and among multiple groups were evaluated with the Student's t-test and analysis of variance, respectively; p < 0.05 was considered statistically significant.
Results
Purification of recombinant human neuritin
After induction of P. pastoris for 96 hr with 1% methanol, a recombinant protein of about 11 kDa was detected in the culture supernatant, demonstrating that neuritin was efficiently expressed (Fig. 1B). A Western blot analysis showed that the protein was specifically recognized by the antineuritin antibody, confirming that the protein was stable (Fig. 1C). The molecular weight of the recombinant protein was 10,957.705 Da, as determined by mass spectrometry (Fig. 1D).
Rat model of acute SCI
In order to create a uniform injury, an extra metal pad was placed on the rat's T10 vertebra (Fig. 2A) such that the 15 g weight dropped from a height of 20 cm inflicted injury to the SC with 300 g·cm of potential energy. Edema and hemorrhage occurred after SCI; the rat tail-pinch and lower limb retraction reflexes were abolished, and both hind limbs exhibited flaccid paralysis, demonstrating that SCI was modeled successfully. This was confirmed by assessing motor function in rats with the BBB scale; compared with the sham group, scores were lower for rats in the SCI than in the sham group at each time point (Fig. 2B). H&E staining revealed extensive hemorrhage, necrosis, and inflammatory cell infiltration in the injured SC; moreover, the nuclei of neurons became light and pyknotic, and were fewer in number, whereas there were also fewer Nissl bodies in the damage zone (Fig. 2D). These results demonstrate that severe injury was inflicted to the SC in the SCI model.

Evaluation of the rat SCI model.
Exogenous neuritin restores motor function after SCI in a dose-dependent manner
A preliminary concentration gradient test with purified recombinant neuritin was carried out in order to determine the effective concentration required for nerve repair. Since a concentration of 5 μg was found to promote sciatic nerve regeneration in rats, we tested concentrations of 30, 60, and 90 μg/ml for the SC. Four weeks after SCI, the regenerative capacity of axons was increased in rats treated with neuritin at each dose compared with those treated with the His tag (Fig. 3A, B). The greatest functional recovery was observed at a concentration of 60 μg/ml (p < 0.05 vs. His), as evaluated by BBB score (Fig. 3C–E). These results indicate that the purified neuritin protein has biological activity at 60 μg/ml, and this concentration was therefore used in subsequent experiments.

Effect of recombinant neuritin treatment at different doses and treatment times.
Recombinant neuritin alleviates nerve damage by suppressing neuronal apoptosis
At various time points after SCI (6 hr and 3, 7, 14, 28, and 56 days), rats injected with 60 μg/ml neuritin or His protein were examined for the expression of PARP, an early molecular marker of apoptosis. 20 Starting at 7 days, the number of Nissl bodies and synapses increased, whereas the number of vacuolated cells decreased in neuritin-treated rats compared with control rats (Fig. 4A). PARP protein degradation increased after SCI, reaching a peak at 3 days before disappearing by day 28, as determined by Western blotting. From 6 hr to 28 days postinjury, PARP degradation was reduced in neuritin-treated rats compared with His-treated rats (Fig. 4B, C), indicating that neuritin treatment inhibits neuronal apoptosis induced by SCI.

Determination of SCI index.
Recombinant neuritin promotes axonal regeneration after SCI
NF200 is the main component of the axonal cytoskeleton; under normal conditions, there is little or no NF200 in the cell body. 21 A close correlation has been observed between the number of NF200-positive neurons and recovery of hind limb function; therefore, NF200 reflects not only the functional status of neurons but also the degree of axonal regeneration. 22 In the present study, NF200 expression was higher in the dorsal horns of the neuritin group than in the His group at each time point from 7 to 56 days after SCI, as detected by immunohistochemistry and Western blot analysis (Fig. 5A–D) (p < 0.05).
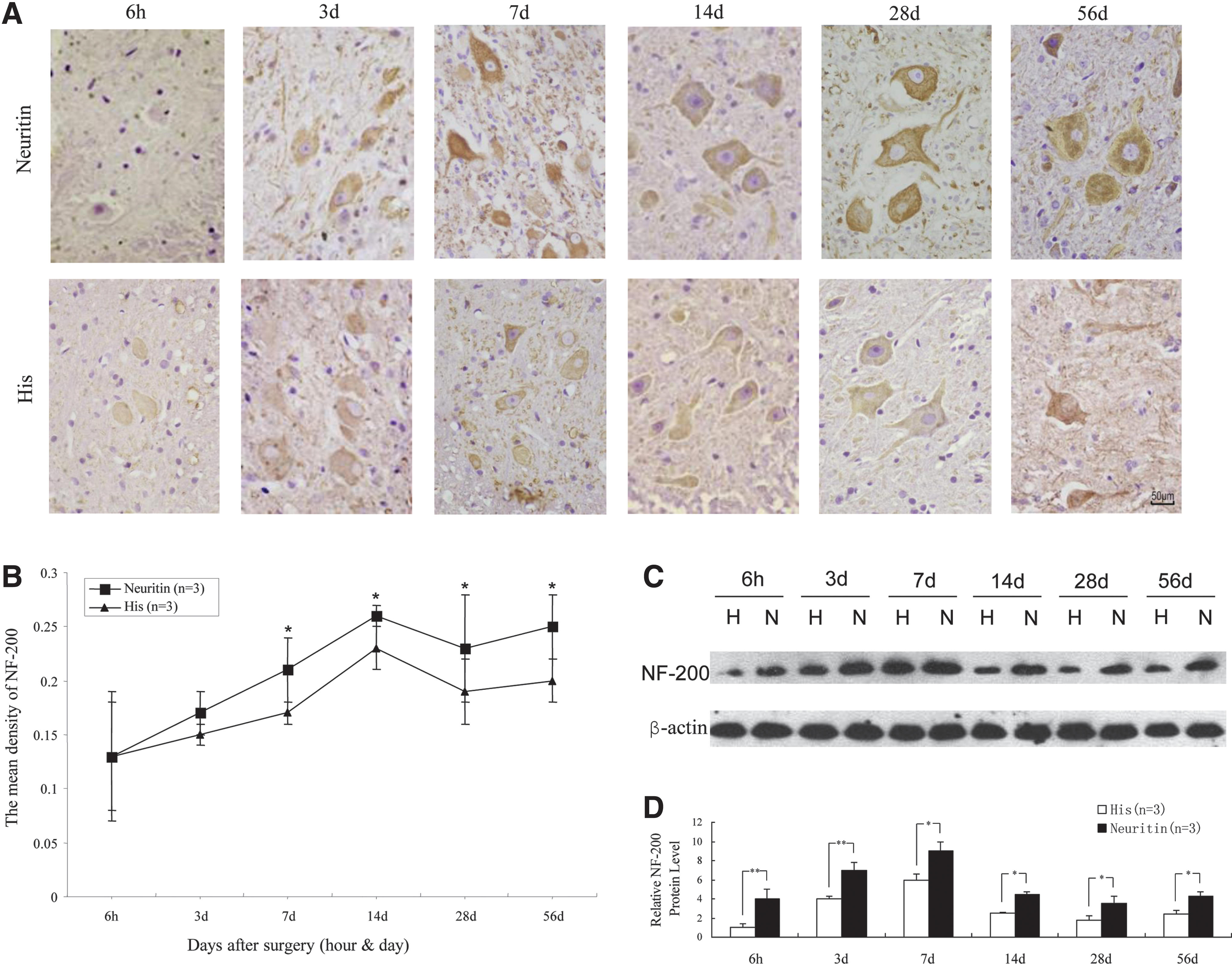
Axonal repair index in rats subjected to SCI as evaluated by NF-200 expression.
GAP-43 is a phospholipoprotein that is expressed in neurons, Schwann cells, and glia; it is considered as a marker of neuronal growth, axonal regeneration, and plasticity. 23 GAP-43 expression in axons is upregulated by 20- to 100-fold after nerve injury and during regeneration. 24 As such, GAP-43 level is a molecular index for axonal regeneration. In the present study, GAP-43 levels were higher in the dorsal horns of the neuritin group than in the His group from 7 to 14 days postinjury, as determined by immunohistochemistry and quantitative analysis (Fig. 6A, B) (p < 0.05). A similar trend was observed by Western blotting (Fig. 6D, E). These results indicate that neuritin treatment promotes the repairing of damaged axons.
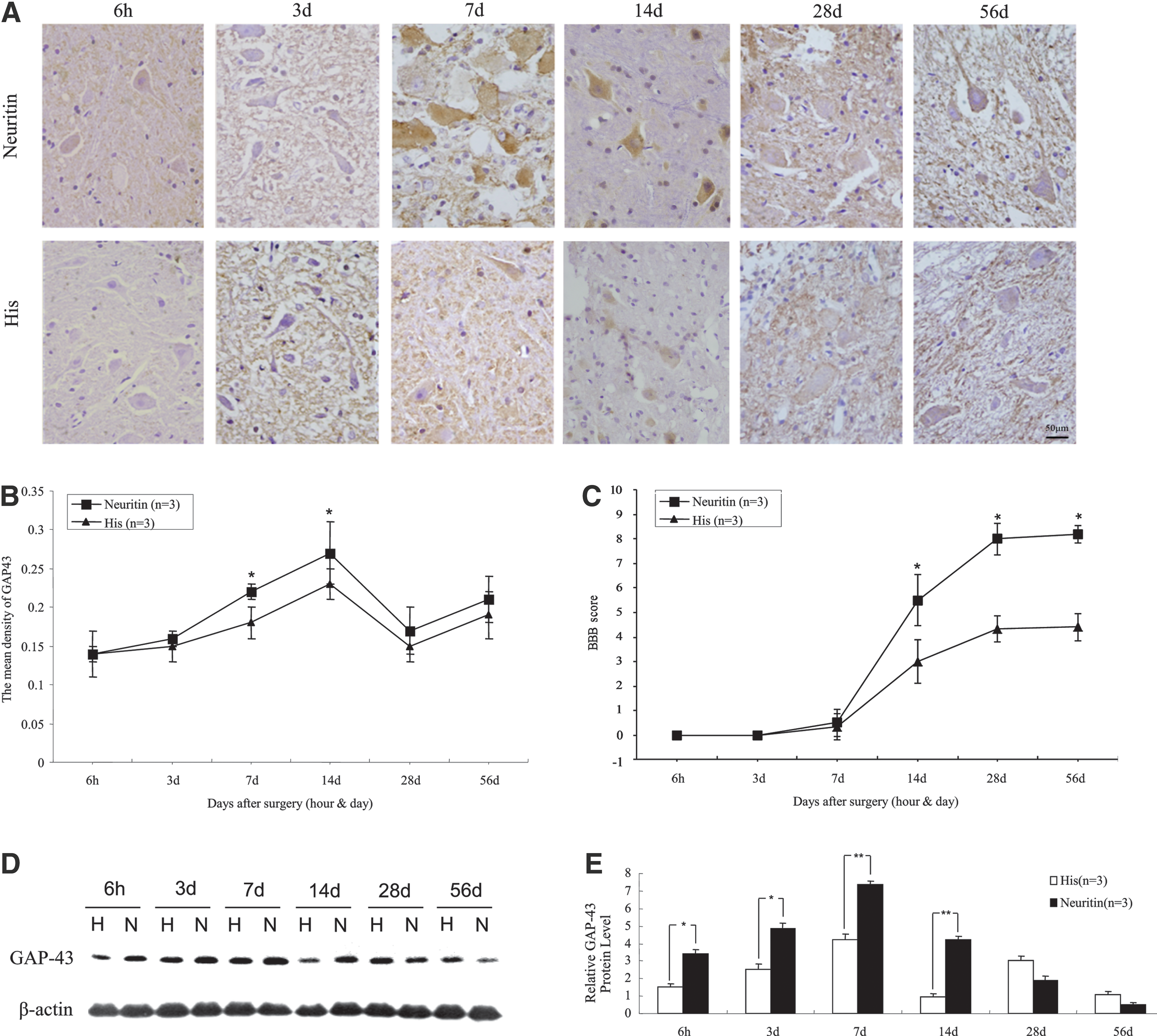
Axonal repair index as evaluated by GAP-43 expression and recovery of locomotor function in rats subjected to SCI.
The BBB scores for each group increased with time after SCI. In the early stages after injury, there was no difference in BBB scores between the neuritin and His groups (p > 0.05); however, starting from 14 days postinjury, the score for neuritin-treated rats was consistently higher than that of His-treated animals (Fig. 6C) (p < 0.05). These results demonstrate that the axonal regeneration in the SC induced by administration of exogenous neuritin leads to an improvement in motor function.
Discussion
The amino acid sequences of rat and human neuritin are identical 9 ; we therefore expressed human neuritin cDNA with an N-terminal 6× His tag in P. pastoris to obtain the recombinant neuritin protein, which was purified by Ni-NTA affinity chromatography. The binding of Ni2+ and 6× His was carried out under nondenaturing conditions. This preserved the biological activity of the protein, which was used directly for functional studies without the need for renaturation steps.
Recombinant neuritin has a molecular weight of 11 kDa; its molecular properties are such that it does not readily pass through the blood–SC barrier. 25 We therefore used intrathecal administration rather than intravenous or intramuscular injection in order to ensure that the protein was delivered to the area of injury at a high concentration. However, neurotrophic factors typically have a short half-life 26 ; for this reason, we administered neuritin daily at 6 μg/day for 1 week into the subarachnoid space through a catheter to avoid making repeated puncture wounds and to stably maintain the effective concentration of neuritin in the SC.
After SCI, the area of damage gradually expands for up to 7 days. Apoptosis rate and changes in the spatial distribution of nerve cells and glia during this period are correlated with the expansion of the damage zone, 27 suggesting that the 1-week period after SCI is critical for mitigating secondary injury, which can be accomplished by providing sufficient levels of neurotrophic factor that can protect the remaining nerves, promote regeneration, and ultimately preserve neurological function. 28 –32 Neuritin has demonstrated effects in nerve repair not only in SCI, 15 but also in cerebral ischemia-reperfusion injury, 33 Alzheimer's disease, 34 hypoxia, 35 traumatic brain injury, 36 electroconvulsive therapy, 37,38 and after antidepressant treatment. 39 Given that the expression level of neuritin is positively correlated with the recovery of neural functioning, overexpressing the protein would be predicted to accelerate the repair of damaged nerves. Recent studies have demonstrated that neuritin overexpression fully restored dendritic complexity and normalized dendritic spine density, mitigating cognitive function deficits and other neural defects of Alzheimer's disease. 34,40 Additionally, application of recombinant neuritin improved retinal ganglion cell survival and stimulated axonal regeneration of a crushed optic nerve in vitro and in vivo. 41 These results suggest that administering neuritin is a potential therapeutic approach for neurodegenerative diseases and central nervous system injury.
We found here that application of exogenous neuritin promoted neurite regeneration in surviving neural cells and inhibited PARP protein degradation in the early stages of SCI. PARP protects damaged DNA from nuclease hydrolysis and guides the binding of DNA repair enzymes 42 ; this process consumes ATP and nicotinamide adenine dinucleotide (NAD+). Excessive DNA injury leads to the consumption of large amounts of NAD+ and ATP because of PARP hyperactivation, resulting in the cell death from energy depletion. 43 A certain amount of intracellular ATP and NAD+ is also a precondition for apoptosis. 44 When PARP was cleaved into P89 and P21 by caspases 3 and 7 upon severe DNA damage, DNA repair was inhibited, and intracellular ATP was not depleted by PARP, which ensured an adequate supply of energy for apoptosis. 45,46 Previous studies have reported that neuritin blocks neuronal apoptosis by inhibiting caspase 3 activation. 10,47 We propose that inhibiting caspase 3 activation will reduce PARP protein degradation, decrease neuronal apoptosis, and ultimately alleviate secondary damage after SCI.
The preservation of remaining axons and stimulation of their repair and regeneration are the most important elements for improving patient outcome after SCI. 48 GAP-43 is a membrane phosphoprotein that is implicated in neural development, axonal regeneration, and synaptic reconstruction. 49 It is primarily expressed in axon terminals and at the presynaptic membrane, and GAP-43 expression level is correlated with axonal regeneration and is therefore an important indicator of this process. 50 GAP-43 participates in the release of calmodulin and may induce the expansion of the presynaptic membrane by promoting vesicle fusion or endocytosis at the presynaptic terminal. 51 GAP-43 also can induce the aggregation of F-actin to counter the retraction of growth cones. 52 Meanwhile, NF200 is the main cytoskeletal component of axons, and plays an important role in the maintenance of neuronal function and axonal transport; it is therefore closely linked to the repair of damage resulting from SCI. 22 In our study, we found that exogenous neuritin increased NF200 and GAP-43 expression in the dorsal horns during the critical 1-week time window after SCI during which axonal regeneration is initiated. The upregulation of these two factors likely promoted the sprouting of axon collaterals and dendrites, which increased the surface area of nerve endings so that new contacts could be established, thereby leading to the recovery of motor function. GAP-43 and NF200 expression gradually diminished after synapse formation. On day 28 after injury, the expression of NF200 but not of GAP-43 differed significantly from control levels. Neurite sprouting and regeneration are orderly processes that require GAP-43 for growth cone extension and axon guidance as well as a variety of intra- and extracellular signaling proteins that remodel the cytoskeleton. 53 Our results demonstrated that early SCI is a critical period for intervention, and that applying the recombinant neuritin protein during this time window promotes axonal regeneration and the recovery of motor function by accelerating cytoskeletal remodeling and synaptogenesis.
In addition to its roles in neurite extension and axonal regeneration, neuritin has been shown to be involved in neuronal proliferation and apoptosis in the context of tumorigenesis. For example, neuritin is overexpressed in the U251 human astrocytoma cell line and in glioma cells 54,55 ; Kaposi's sarcoma and invasive breast carcinoma also show high levels of neuritin expression. 56,57 These findings imply that neuritin expression level is positively correlated with tumor malignancy. Thus, neuritin has dual roles in nerve regeneration and tumorigenesis, and future research should focus on how its positive regulatory role in the nervous system can be potentiated while suppressing its detrimental functions.
In conclusion, our study demonstrated that the application of exogenous recombinant neuritin can inhibit nerve cells apoptosis, induce the upregulation of NF200 and GAP-43 expression, and accelerate the recovery of hind limb motor function after SCI. These findings indicate that exogenously applied neuritin creates a favorable microenvironment for neuronal survival and axon regeneration, and suggest that neuritin can be used as a potential new therapeutic strategy for improving the outcome of SCI patients.
Footnotes
Acknowledgments
This study was supported by grants from the Supporting Xinjiang Project (No. 2014AB048), the Major Technology Innovation Project of Hangzhou (No. 20152013A01), the Natural Science Foundation of China (No. 31560269), and the Foundation for High-level Talents of Shihezi University (No. RCZX201448).
Author Disclosure
No competing financial interests exist.