
Abstract
Corneal disease is one of the leading causes of blindness worldwide. Gene therapy is an attractive therapeutic strategy for corneal diseases, but currently underdeveloped. Recombinant adeno-associated viral (rAAV) vectors have emerged as a highly promising gene therapy platform. This study aims to identify rAAV vectors that can efficiently transduce corneal stroma for potential applications in studying pathophysiology of corneal diseases and therapeutic development. We characterized 14 rAAV serotypes expressing enhanced green fluorescent protein (EGFP), for cell specificity and transduction efficiency after either intrastromal injection or topical administration in mouse corneas in vivo. Our results show that intrastromal injections of rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 efficiently transduce mouse corneal stroma in vivo, and that topical administrations of rAAVrh.10 and rAAVrh.39 subsequent to epithelial scraping generate detectable transgene expression. In vivo imaging analysis revealed that transgene expression became detectable by 1 week postadministration, peaked at 2 weeks, and lasted for the duration of the study (i.e., 4 weeks). Both rAAVrh.10 and rAAVrh.39 transduced more than 50% of keratocytes, the major cell type in the corneal stroma, by intrastromal injection and 30% by topical administration. Histopathology indicated that rAAV transduction of cornea caused no morphological adverse effects. Overall, our findings suggest that some rAAV serotype vectors can efficiently transduce corneal stroma in vivo, constituting a potentially powerful and safe gene delivery platform for gene therapy of corneal diseases.
Introduction
C
rAAV, a single-stranded DNA virus, is an attractive vehicle for gene delivery because of its low immunogenicity, low genotoxicity, broad tropism, high transduction efficiency, and long-term expression in vivo. 3 –6 Because the predominant residing stromal cell type is the keratocyte, a specialized fibroblast that plays a role in general repair and maintenance, 7 it is no surprise that both corneal stroma and corneal fibroblasts have become major targets for rAAV-based corneal gene therapy.
AAV2 was the first tested serotype that demonstrated significant transgene delivery in the rabbit cornea in vivo. 8 Since then, additional serotypes including rAAV5, rAAV6, rAAV8, and rAAV9 vectors have been studied for gene delivery into human corneal fibroblasts in vitro, mouse corneas in vivo, and human corneas ex vivo. 9 –11 With the discovery of a large primate AAV family with unique tissue tropism, it is becoming imperative to further expand the list of AAV serotypes for efficient gene delivery to the cornea and select serotypes with higher transduction efficiency for the development of rAAV-mediated gene therapies for corneal diseases.
AAV gene transfer efficiency and duration of delivered transgene expression depend on serotype and capsid architecture. 4,5 Thus in the present study, for the first time, we comprehensively screened and tested 14 rAAV serotypes for their in vivo transduction capability specific to mouse corneal stroma. Among the numerous delivery strategies widely used for in vivo gene delivery to the cornea we chose two well-established methods that allow direct access of rAAV vectors to the stroma, namely topical administration after lamellar keratotomy and laser (or mechanical) ablation of the epithelium, 10,12 and direct intrastromal injection, which was first introduced by Carlson and colleagues in 2004. 13 We identified rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 as the leading serotypes for transducing corneal stroma via intrastromal injection, among which only rAAVrh.10 and rAAVrh.39 efficiently transduced corneal stroma by topical administration after epithelium removal. Of the major corneal stromal cells, rAAVrh.10 and rAAVrh.39 transduced more than 50% and 30% of keratocytes via intrastromal and topical administration, respectively, without noticeable adverse effects. Taken together, our study provides new rAAV serotype candidates that are safe and highly efficient in mice and carry great potential for future preclinical and clinical development of rAAV-based gene therapy for corneal diseases.
Materials and Methods
Materials and animals
Primary rabbit anti-mouse keratocan antibody was obtained from Santa Cruz Biotechnology (Dallas, TX). Primary rabbit anti-mouse GFP antibody, goat anti-rabbit IgG (H+L) secondary antibody–Alexa Fluor 488 conjugate, and goat anti-rabbit IgG (H+L) secondary antibody–Alexa Fluor 568 conjugate were purchased from Life Technologies (Grand Island, NY). VECTASHIELD anti-fade mounting medium with 4′,6-diamidino-2-phenylindole (DAPI) was obtained from Vector Laboratories (Burlingame, CA).
Six- to 8-week-old female C57BL/6J mice (Charles River Laboratories, Wilmington, MA) were maintained and used according to the guidelines of the Institutional Animal Care and Use Committee (IACUC) of the University of Massachusetts Medical School (Worcester, MA). All animal experiments conformed to the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research. Before experimental operation, all animals were anesthetized by an intraperitoneal injection of a ketamine–xylazine (100 and 10 mg/kg, respectively) mixture. The right eyes of mice were treated.
rAAV vector production
rAAV vectors were generated by triple-plasmid transfection of HEK293 cells. The self-complementary pAAV-CB-PI-EGFP plasmid was used for packaging with capsids from 14 different serotypes to produce rAAV1, rAAV2, rAAV3b, rAAV4, rAAV5, rAAV6, rAAV6.2, rAAV7, rAAV8, rAAV9, rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43. 14 Viruses were purified by CsCl gradient ultracentrifugation and titered by both quantitative polymerase chain reaction (qPCR) and silver staining of sodium dodecyl sulfate (SDS)–polyacrylamide gels. 14
rAAV transduction of mouse cornea by intrastromal injection
Intrastromal injection was performed according to a previously published procedure 11 (Fig. 1A). Briefly, a 1.0-mm-long incision through the corneal epithelium, equidistant between the corneal–scleral junction and the corneal center, was first created with the tip of a 26-gauge needle. The tip of a 33-gauge needle attached to a 5-μl Hamilton microliter syringe (Hamilton, Reno, NV) was then introduced through the incision into the corneal stroma and 2.4 × 1010 genome copies (GC) of vector in 4 μl of phosphate-buffered saline (PBS) was injected. Antibiotic ointment was applied after injection.

Overview of intrastromal injection and topical administration of rAAV vector to cornea.
rAAV transduction of mouse cornea by topical administration
rAAV transduction via topical administration was conducted with or without the corneal epithelium removed (Fig. 1B). Corneal epithelium scraping was performed accordingly. 10 Alcohol-soaked filter membrane was applied to each cornea for 20 sec and the whole layer of epithelium covering about 80% of the corneal area was removed by gentle scraping with a #64 Beaver blade (Beaver-Visitec, Waltham, MA) under an operating microscope. Viral vector (2.4 × 1010 GC) in 4 μl of PBS was directly applied to the intact cornea or to the corneal stroma after epithelium removal and allowed to sit for 2 min. After drying the cornea with an antiseptic cotton swab, antibiotic ointment was applied.
In vivo microscopy studies
Animals of each group were observed in vivo 1, 2, 3, and 4 weeks after rAAV administration. Imagery of EGFP expression in the mouse eye was captured with a Micron III camera (Phoenix Research Labs, Pleasanton, CA).
Histological and immunofluorescence–histochemical analysis
After sacrifice, mouse eyes were enucleated. Eight eyes from each group were fixed in 4% paraformaldehyde. Among them, four corneas with limbus were harvested for corneal flat mounts, which were blocked in 5% goat serum in PBS and stained with rabbit anti-mouse GFP primary antibody (diluted 1:1000), followed by goat anti-rabbit IgG–Alexa Fluor 488 secondary antibody (diluted 1:1500). The corneal whole mounts were then mounted for observation and imaging analysis.
The remaining four eyeballs harvested from each group were embedded in O.C.T. compound (Fisher Scientific, Pittsburgh, PA) for cryosectioning at a thickness of 10 μm, and then blocked in 5% goat serum and stained with rabbit anti-mouse keratocan primary antibody (diluted 1:50) and goat anti-rabbit IgG–Alexa Fluor 568 secondary antibody (diluted 1:1500). All immunofluorescence stained sections were mounted with VECTASHIELD medium containing DAPI, and fluorescence detection of native EGFP expression and stained keratocan in eyeball samples was generated with a Leica DM5500 microscope. The embedded samples were stored at −80°C.
In the meantime, two eyes from each group were fixed in 10% formalin and embedded in paraffin to be sectioned later at 4-μm thickness and stained with hematoxylin and eosin (H&E) for histological analysis. Images were obtained with a Leica DMC2900 microscope.
Quantification analysis of EGFP expression in corneal whole mounts
Digital images of the corneal whole mounts were taken with the Leica DM5500. EGFP-positive area and fluorescence signal intensity were measured on these flat mounts, using ImageJ software (National Institutes of Health, Bethesda, MD;
Quantification analysis of EGFP expression in cryosections
Four eyeballs from each group were fixed and cryosectioned for keratocan immunostaining. For each sample, images of five corneal slides were captured. EGFP+ cells and keratocan+ cells on each image were counted separately, and the number of colocalized cells was then obtained using Imaris 8 software (Bitplane, Concord, MA) to determine percentages of EGFP+keratocan+ cells among keratocan+ cells.
Quantification analysis of rAAV genome copy number and RNA expression
Corneas of three eyes in each group were harvested. Genomic DNA was isolated with a QIAamp DNA kit (Qiagen, Hilden, Germany) in accordance with the manufacturer's instructions, and then digested with SalI (New England BioLabs, Ipswich, MA) at >10 U/μg of DNA at 37°C for 1 hr. There are two SalI sites in the rAAV genome, and SalI digestion ensures single copies of EGFP transgene for droplet digital PCR (ddPCR) quantification. Multiplexed ddPCR was performed on a QX200 ddPCR system (Bio-Rad Laboratories, Hercules, CA) using TaqMan reagents targeting EGFP (Cat. No. 4400293; Life Technologies, Carlsbad, CA) and the reference gene transferrin receptor (Tfrc) (Cat. No. 4458367; Invitrogen, Waltham, MA). For each sample, about 10 ng of template was used in 40 μl of ddPCR mix with 1 μl of TaqMan reagents targeting EGFP and 1 μl of targeting Tfrc. Cycling conditions were used according to the manufacturer's manual (
Statistical analysis
Results were expressed as means ± SD. Analysis was performed by one-way analysis of variance (ANOVA) for multiple variables, and Bonferroni's post hoc multiple-comparison test was used for between-group differences, using GraphPad Prism 6.0 (GraphPad Software, La Jolla, CA); p < 0.05 was considered significant in all cases.
Results
Intrastromal injections of rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 transduce cornea efficiently
To evaluate the transduction efficiency of 14 different rAAV serotypes, we intrastromally injected rAAV1, rAAV2, rAAV3b, rAAV4, rAAV5, rAAV6, rAAV6.2, rAAV7, rAAV8, rAAV9, rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 expressing EGFP at a dose of 2.4 × 1010 GC per eye, and PBS as a negative control, into normal mouse corneas (Fig. 1A). Compared with the PBS control, we observed robust EGFP expression in corneas at week 4 after injection with rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 (Fig. 2A); the percentage of EGFP-positive area and EGFP intensity in the corneal whole mounts of those four groups are presented in Fig. 2B and C. Nearly 80% of the whole corneal area was efficiently transduced with rAAVrh.8, rAAVrh.10, and rAAVrh.39 vectors (Fig. 2B), and the EGFP intensities associated with rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 were approximately 4-fold stronger than those of the other rAAV serotypes tested (Fig. 2C). These results demonstrated that rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 could transduce mouse cornea in a highly efficient manner.

Intrastromal injection of rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 efficiently transduces mouse cornea.
Topical administrations of rAAVrh.10 and rAAVrh.39 transduce the cornea with corneal epithelium removed
To explore the feasibility of delivering those four leading rAAV serotypes to mouse cornea by a straightforward and noninvasive method, we evaluated EGFP transduction by rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 after topical administration to the cornea (Fig. 1B). Because the corneal epithelium is known to be a natural barrier for topical therapeutics to the corneal stroma, we performed topical application of the four leading rAAV vectors either with or without corneal epithelium scraping in comparison with intrastromal injections of the same vectors at the same dose (2.4 × 1010 GC per eye). Corneal whole-mount imaging showed that all four serotypes were unable to transduce mouse cornea when the epithelium was present; however, rAAVrh.10 and rAAVrh.39 efficiently transduced corneal stroma when the epithelium was removed (Fig. 3A). In comparison with intrastromal injections, topical administration of these two serotypes after epithelium removal produced similar percentages of EGFP-positive area (Fig. 3B), but exhibited significantly lower EGFP intensity (p < 0.001) (Fig. 3C). To determine whether there is any possible correlation between the capsid sequences and transduction biology among the four serotypes when applied topically, we analyzed the amino acid sequences of those capsids for homology. Such an analysis indicated that rAAVrh.10 and rAAVrh.39 are more closely related, whereas rAAVrh.8 and rAAVrh.43 are more distant from rAAVrh.10/rh.39 (Supplementary Fig. S1; Supplementary Data are available online at
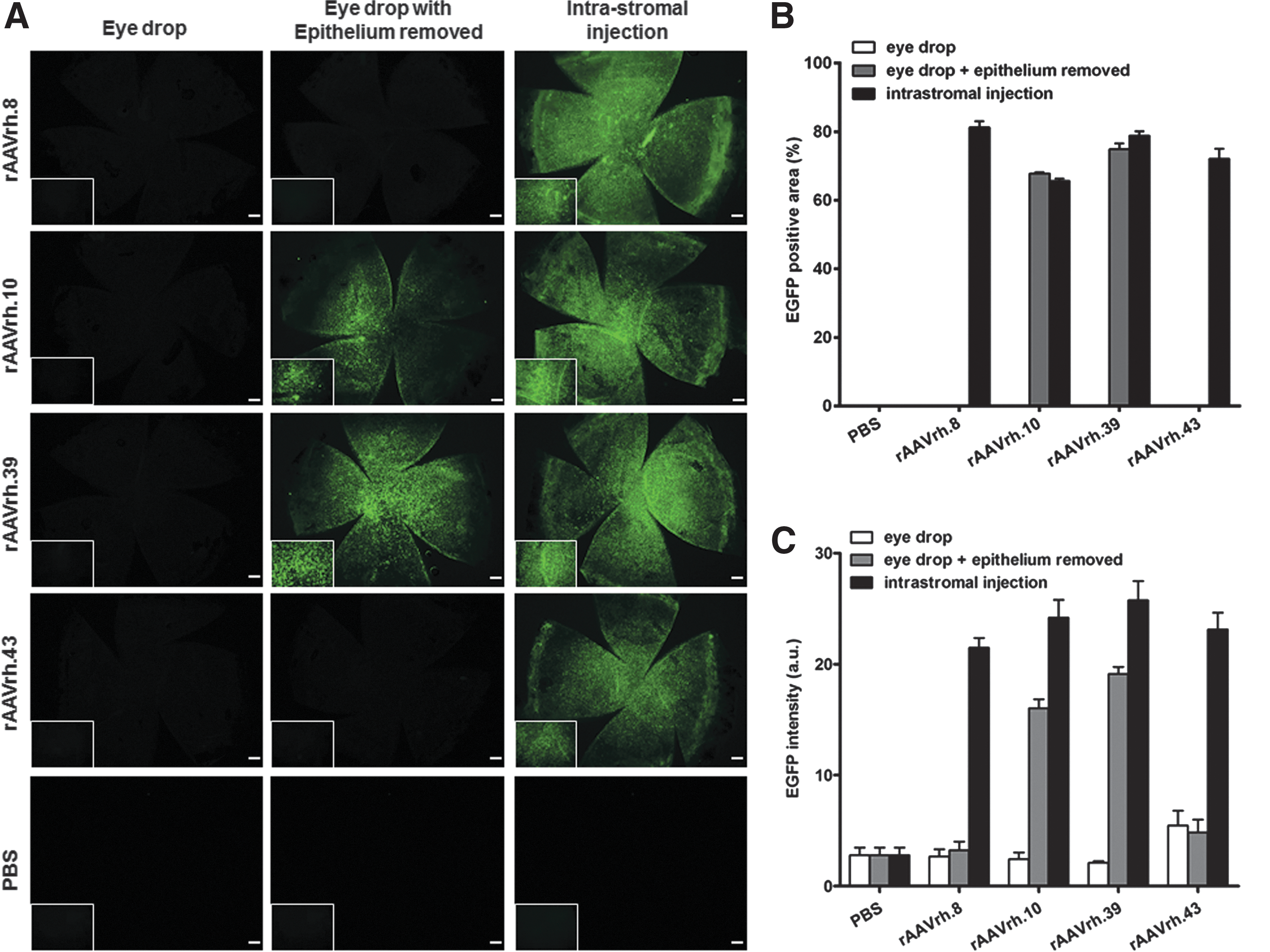
Topical administration of rAAVrh.10 and rAAVrh.39 transduced mouse cornea when the corneal epithelium was removed.
rAAVrh.10 and rAAVrh.39 achieve sustained corneal transduction
After topical administration (eye drop with corneal epithelium removed) or intrastromal injection, EGFP expression from rAAVrh.10 and rAAVrh.39 was apparent at 1 week posttreatment, reached peak expression at about 2 weeks, and remained detectable at the 4-week study endpoint (Fig. 4A) when mouse corneas were harvested for ddPCR quantification of vector genome copies and EGFP mRNA levels. Our data showed that on average, more than 5000 GC per cell persisted at 4 weeks posttreatment for intrastromally injected rAAVrh.10 and rAAVrh.39 serotypes, and approximately 200 GC per cell was detected for the two serotype vectors when topically administered (Fig. 4B). It is worth pointing out that the vector genome abundance in corneas that received rAAV intrastromally is 20- to 30-fold higher than that in mouse liver treated with an intravenous injection of rAAV9; EGFP was approximately 40-fold higher than the dose of rAAVrh.10 and rAAVrh.39 (Fig. 4B). This suggests that, on a per-genome basis, intrastromal delivery of rAAVrh.10 and rAAVrh.39 to the cornea could be more efficient (800- to 1200-fold) than systemic delivery of a more concentrated and highly liver-tropic rAAV9 vector. In addition, EGFP mRNA expression levels in the rAAV-treated corneas were well correlated with the abundance of vector genomes (Fig. 4C).

rAAVrh.10 and rAAVrh.39 transduction in mouse cornea continued for at least 4 weeks in vivo.
rAAVrh.10 and rAAVrh.39 primarily target keratocytes in corneal stroma
As keratocytes with characteristic interconnecting dendritic processes comprise 96% of the cornea in mice and humans, 7 we next attempted to characterize the keratocytic tropism of rAAVrh.10 and rAAVrh.39, using keratocan as a cell marker for keratocytes (Fig. 5A). Our quantitative analysis of EGFP+keratocan+ cells in the corneal stroma revealed that rAAVrh.10 and rAAVrh.39 transduced 51.1 ± 3.0% and 55.97 ± 3.5% of keratocytes, respectively, by intrastromal injection, and 35.9 ± 8.1% and 33.64 ± 7.7% of keratocytes, respectively, by eye-drop application to the epithelium-removed corneas (Fig. 5B). In other words, intrastromal injection of rAAVrh.10 and rAAVrh.39 transduced 1.5- to 2-fold more keratocytes when compared with topical administration (Fig. 5B), which was aligned with results from the ddPCR quantification of EGFP mRNAs (Fig. 4C).
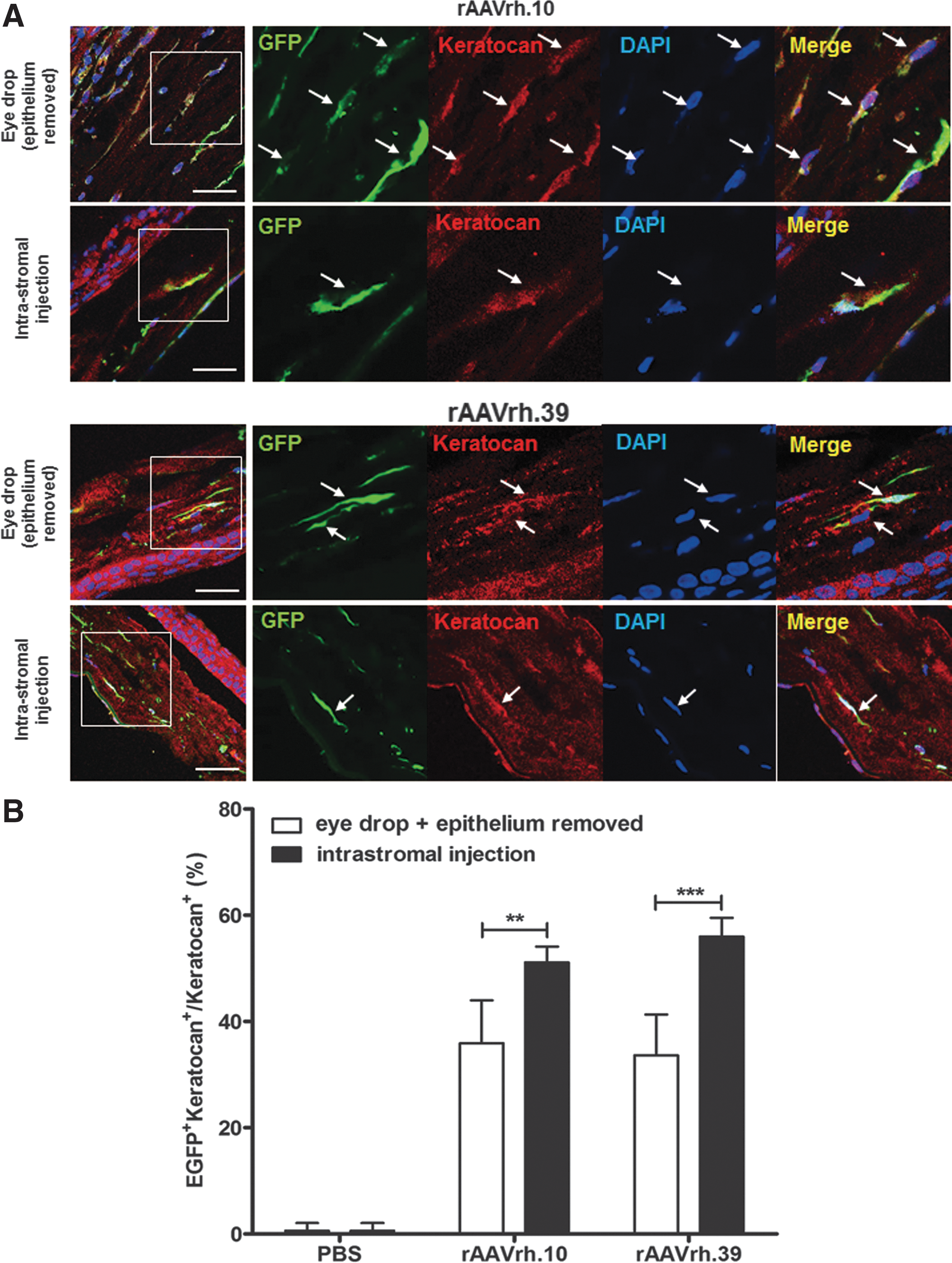
rAAVrh.10 and rAAVrh.39 transduce mouse keratocytes in corneal stroma.
Corneal transduction with rAAVrh.10 and rAAVrh.39 vectors causes no histopathology
To evaluate possible vector-related toxicity caused by rAAV transduction in the corneal stroma, we analyzed histopathology of corneal tissues treated with rAAVrh.10 and rAAVrh.39 vectors at 4 weeks posttreatment. Histological images of H&E-stained tissue sections of corneas treated with rAAVrh.10 and rAAVrh.39 via either topical eye drops or intrastromal injection presented structures and morphologies similar to those that received PBS or no treatment; all corneal stromas remained organized into interweaving collagen lamellae with an even distribution of keratocytes (Fig. 6). These findings suggest that rAAV transduction of the corneas induced no adverse effects on corneal stromal morphology throughout the course of 4 weeks, implying that rAAV did not deter overall health of the cornea tissue.
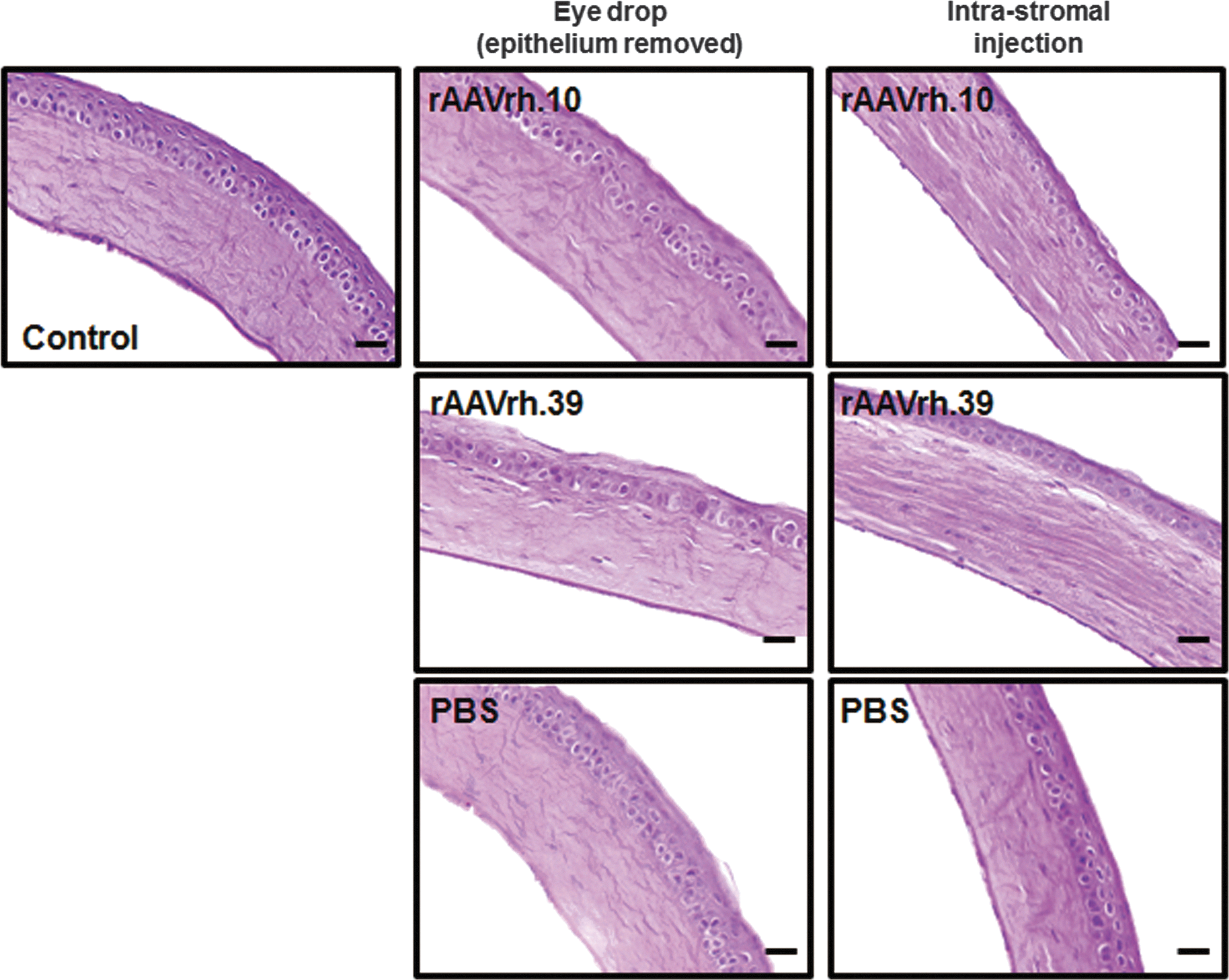
Transduction of rAAVrh.10 and rAAVrh.39 vectors had no adverse effect on cornea histology. Paraffin-embedded sections of the corneas stained with hematoxylin and eosin (H&E) displayed normal structures: all layers of the cornea were clear without obvious morphological changes compared with the control group or any other signs of inflammatory and immune reactions. Original magnification: ×200; scale bar, 50 μm.
Discussion
Since the discovery of a large family of novel AAVs, many of those new AAVs have been extensively evaluated for their abilities to transduce a variety of tissues. 15,16 Our study is the first to comprehensively characterize a large panel of rAAV serotypes for their ability to transduce mouse cornea in vivo. Two well-established delivery methods, intrastromal injection and topical administration, were evaluated to investigate the transduction efficiency of 14 different rAAV serotypes. We identified rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 as the leading serotypes for transducing corneal stroma by intrastromal injection and rAAVrh.10 and rAAVrh.39 for corneal stroma targeting by topical administration after epithelium removal.
Considering that rAAV vectors are safe and efficient to transduce cells in vivo, there is growing interest in using rAAV to treat corneal pathologies such as corneal neovascularization. 17,18 Although many diseases can lead to corneal neovascularization, the direct cause is not always known, and targeting neovascularization through rAAV-mediated gene transfer might be one effective option to prevent long-term complications. In addition, rAAV displays serotype-dependent tropism and transduction efficiency, which can be exploited to achieve cell type-restricted transduction. Thus, it is important to characterize AAV serotype(s) for efficient gene transfer into cornea, especially the corneal stroma, which plays a vital role in corneal transparency.
In the present study, we demonstrate for the first time that rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 are able to transduce mouse cornea at efficiencies significantly higher than previously reported serotypes (rAAV5, rAAV6, rAAV8, and rAAV9). 9 –11 EGFP expression levels of those previously reported rAAV serotypes were consistent with our findings, with rAAV9 taking the lead and rAAV5 showing the weakest performance (rAAV9 > rAAV8 > rAAV6 > rAAV5) 10,11 ; the reproducibility of previous findings by our study also demonstrates the consistency of corneal transduction by rAAV. For this comparative analysis we attempted to analyze EGFP transduction, using two strategies. First, using the area of EGFP-positive signal can indicate the spread of the vector. On the other hand, EGFP intensity could be more indicative of expression levels of a functional transgene product. We found that the vector spread and EGFP intensity did not always correlate. One possibility could be that limited vector spread leads to high EGFP intensity in a relatively small area. However, this will still depend on the number of cells transduced by functional viral particles. For example, rAAV1 shows an EGFP-positive area of about 40%, whereas the intensity is only about 10%. One possible reason for this finding could be that rAAV1 in cornea can distribute relatively well, but only low levels of functional transgene expression were achieved in the transduced cells. In contrast, rAAVrh.43 seems to result in an about 50% EGFP-positive area (but lower than that of rAAVrh.39) whereas the EGFP intensity is about the same between rAAVrh.43 and rAAVrh.39. This could be interpreted as meaning rAAVrh.43 cannot spread intracorneally as well as rAAVrh.39 but results in a similar amount of EGFP expression.
To highlight the versatility of rAAV to transduce mouse cornea, we compared gene transfer efficiencies of these four rhesus serotypes on delivery by two major techniques, allowing vector access to corneal stroma via topical administration or intrastromal injection. Intrastromal injection is a commonly used clinical procedure to deliver therapeutics to the cornea, whereas topical eye dropping is a noninvasive, straightforward, and routinely used method to apply ophthalmological drugs to the eyes. In general, the gene transfer efficiency of rAAV vectors was higher when delivered by intrastromal injection as compared with topical application. For topical administration, none of the four serotypes (rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43) mediated transgene expression with the corneal epithelium intact, highlighting the function of the corneal epithelium as a natural barrier. On the other hand, a lesion made by epithelial scraping after alcohol burn, which potentially mimics a variety of corneal injuries, 19 allowed direct access of rAAVrh.10 and rAAVrh.39 to keratocytes by eye-drop delivery compared with eyes bearing an intact epithelial barrier. Moreover, the differences in capsid amino acid sequences among rAAVrh.8/rAAVrh.43 and rAAVrh.10/rAAVrh.39 might explain the dramatic differences in corneal transduction via eye drops with the epithelium removed, highlighting critical role(s) of the viral capsid in dictating tissue tropism and transduction biology of an AAV vector. It is worth noting that except for the differences in one region that spans seven amino acids, SNGILIFGK for rAAV8 but VTGSCFWQ for rAAVrh.43, the VP3 capsid amino acid sequences of both serotypes are otherwise identical. This may imply that the amino acid differences in this region might have contributed to the dramatic difference in corneal transduction efficiency between rAAV8 and rAAVrh.43. A future study is warranted to further understand correlations between capsid structures and corneal transduction biology for those two AAV vectors. We also observed that intrastromal injection favored higher EGFP intensity compared with topical application. This increased intensity after intrastromal injections may be due to rAAV remaining confined within the corneal stroma, leading to higher transduction, whereas eye-drop delivery disperses rAAV across the ocular bulb, thus losing rAAV to ocular surfaces distal to the cornea. Taken together, these factors might contribute to the differences between the two delivery methods in gene transfer efficiencies. Furthermore, intrastromal delivery potentially could be more efficient than systemic delivery to the liver (Fig. 4B and C). However, this result might be biased by the fact that many rAAV capsids could be trapped in the extracellular space. This is further supported by the ratio of EGFP mRNA to GAPDH mRNA. In acquired corneal diseases, corneal stromal wounds occur from traumas, physical and chemical injuries, or adverse effects of refractive surgery. 20 During wound healing, keratocytes exit quiescence and differentiate into activated myofibroblasts at or adjacent to the site of injury. 11,20 The activated myofibroblasts then migrate toward the damaged area, whereby repopulation of keratocytes may clear the episomal vector genomes of transduced cells, 11 making rAAV vectors a possible transient therapy for treating acquired corneal injuries. Such corneal surface injuries were simulated in the epithelium-removed corneas of our study, and we found that rAAVrh.10 and rAAVrh.39 highly transduced the corneal stroma up to 2 weeks after topical treatment, followed by waning EGFP expression beyond 2 weeks posttreatment, likely due to the repopulation of keratocytes caused by corneal surface damage. The turnover of keratocytes and the incapacity of rAAVs to integrate into the host cell genome suggest that rAAV-mediated gene delivery holds promise as a transient gene therapy for acute cornea-related disease, which is a less studied application of AAV. Furthermore, future studies to explore the contribution of immune response to vector genome loss in the cornea are needed.
Vector-related toxicity is a critical component in the safety profile of gene therapy in any target tissue, emphasizing the risk–benefit relationship of any gene therapeutic. A wealth of published preclinical and clinical studies has documented the safety of rAAV vector-mediated gene therapy for a wide spectrum of target tissues and diseases. 21,22 Our study contributes to the evaluation of the safety profiles of those new rAAV vectors for potential use in the gene therapy of corneal diseases. Nonetheless, further studies are warranted to confirm the safety of rAAV for corneal gene therapy in clinical settings.
In summary, we have carried out an extensive characterization of the major rAAV serotype vectors available, and identified several rAAVs that can efficiently and safely transduce normal cornea in vivo when delivered by two different methods. The performance of rAAVrh.8, rAAVrh.10, rAAVrh.39, and rAAVrh.43 appears to be promising for cornea-related diseases, but additional studies involving dosage, visual acuity changes, and toxicity will need to be evaluated in subsequent studies. Our findings will help to establish a critical foundation to study pathomechanisms and develop treatment for a wide spectrum of corneal diseases.
Footnotes
Acknowledgments
This work was funded by a research grant from the National Nature Science Foundation of China (No. 81273424) to X.X. and L.Y., a corporate sponsored research grant from Chengdu Kanghong Pharmaceutical Group Co., Ltd to G.G., X.X., and Z.Q., and an internal grant from University of Massachusetts Medical School to G.G.
Author Disclosure
G.G. is a co-founder of Voyager Therapeutics, specialized in rAAV-based CNS gene therapy; and holds equity in the company. G.G. is an inventor on patents with potential royalties licensed to Voyager Therapeutics and other biopharmaceutical companies. Z.Q. is a salaried employee of Kanghong Pharmaceutical Group Co., Ltd., Chengdu, China.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.