
Abstract
Osteoporosis is a major healthcare burden affecting mostly postmenopausal women characterized by compromised bone strength and increased risk of fragility fracture. Although pathogenesis of this disease is complex, elevated proinflammatory cytokine production is clearly involved in bone loss at menopause. Therefore, anti-inflammatory strategies hold a great potential for the prevention of postmenopausal osteoporosis. In this study, we investigated the effect of gene therapy of recombinant adeno-associated virus (rAAV)–mediated human alpha-1 antitrypsin (hAAT), a multifunctional protein that has anti-inflammatory property, on bone loss in an ovariectomy-induced osteoporosis mouse model. Adult ovariectomized (OVX) mice were intraperitoneally (i.p.) injected with hAAT (protein therapy), rAAV8-CB-hAAT (gene therapy), or phosphate buffer saline (PBS). Age-matched and sham-operated animals were used as controls. Eight weeks after the treatment, animals were sacrificed and bone-related biomarkers and vertebral bone structure were evaluated. Results showed that hAAT gene therapy significantly decreased serum IL-6 level and receptor activator of NF-κB (RANK) gene expression in bone. Importantly, hAAT gene therapy increased bone volume/total volume and decreased structure model index (SMI) compared to PBS injection in OVX mice. These results demonstrate that hAAT gene therapy by rAAV vector efficiently mitigates bone loss possibly through inhibition of proinflammatory cytokine IL-6 and RANK gene expression. Considering the safety profile of hAAT and rAAV vector in humans, our results provide a new alternative for the treatment of osteoporosis.
Introduction
O
Human alpha-1 antitrypsin (hAAT), mainly produced in the liver, is a 52 kDa serum glycoprotein, and its normal level in serum is 150–350 mg/dl. 5 It is a potent inhibitor of neutrophil elastase and protects the alveolar wall from degradation by neutrophil elastase. 6 During inflammation, the circulating level of hAAT can be increased by up to threefold, indicating its important role in response to inflammation in the human body. 7 Apart from its proteinase inhibitory properties, accumulating evidence indicates that hAAT also has cytoprotective, immunomodulatory, and anti-inflammatory activities. 8 It inhibits acute inflammatory infiltration, affects neutrophil migration, and regulates neutrophil chemotaxis. 9,10 It also inhibits lipopolysaccharide-induced release of TNF-α and IL-1β in human monocytes and enhances the production of the anti-inflammatory cytokine IL-10. 11,12 Considering the evidence regarding hAAT's anti-inflammatory and immunomodulatory properties, we have shown that hAAT therapy has a therapeutic effect on inflammation-related disease models, including type-1 diabetes and rheumatoid arthritis. 13,14 Importantly, a clinical study showed that hAAT-deficient patients have high risk of osteoporosis. 15
Recombinant adeno-associated virus vectors (rAAVs) have been used to express many genes in animal models and in human clinical studies. 16 This vector can deliver transgenes into a wide range of cell types and organs and mediate long-term transgene expression, and can be a useful gene therapy vector for the treatment of chronic diseases including osteoporosis. In this study, we tested the feasibility of rAAV vector-based hAAT gene therapy in parallel with hAAT protein therapy for the treatment of bone loss in ovariectomized (OVX) osteoporosis mouse models.
Materials and Methods
Recombinant adeno-associated virus (rAAV) vector production
The rAAV-CB-hAAT vector construct was produced and packaged as previously described. 17 Briefly, this vector carries hAAT (SERPINA1) cDNA driven by the cytomegalovirus (CMV) enhancer and chicken β-actin (ACTB) promoter. It was packaged into an AAV serotype 8 capsid by cotransfection of the vector plasmid and helper plasmid (XYZ8) into 293 cells. rAAV8-CB-hAAT vectors were purified by iodixanol gradient centrifugation followed by anion exchange chromatography. The physical particle titers of vector preparations were assessed by dot blot analysis. The final rAAV8-CB-hAAT vector were prepared and diluted in Ringer's lactate solution.
Animals
Seven-week-old bilaterally OVX and sham-operated C57BL/6 mice were purchased from Charles River Laboratories, Wilmington, MA. All mice were shipped one week after the operation. They were maintained under a 12-h light/dark cycle and in specific pathogen-free conditions. All experiments were conducted according to protocols approved by the University of Florida Institutional Animal Care and Use Committee.
Human alpha-1 antitrypsin (hAAT) protein and rAAV8-CB-hAAT vector administration
After one week of acclimation period, OVX C57BL/6 mice were randomly divided into phosphate buffer saline (PBS, n = 10), hAAT protein (n = 10), or hAAT gene therapy (rAAV8-CB-hAAT, n = 10) groups. In the PBS group, OVX C57BL/6 mice were intraperitoneally (i.p.) injected with PBS (200 μl, Corning, Manassas, VA). In the hAAT protein treatment group, OVX mice were injected with hAAT (Prolastin C®, Telecris Biotherapeutics Inc., NC; 2 mg/mouse in 200μl PBS). For the hAAT gene therapy group, OVX mice were i.p. injected at week 0 with rAAV8-CB-hAAT vectors (single i.p. injection of 1 × 1011 particles/mouse in 200 μl Ringer's lactated solution). Sham-operated C57BL6 mice (n = 10) were i.p. injected with PBS (200 μl/mouse) and used as model control. The i.p. injection of either hAAT or PBS was performed every 3 days for up to 8 weeks. One mouse from the hAAT injected group died at week two and data from this mouse were excluded from analysis.
Sample collection and preparation
All animals were weighed weekly and bled biweekly by facial vein bleeding to monitor the hAAT levels. All mice were sacrificed by cervical dislocation after 8 weeks of treatment. The vertebrae were removed from each mouse, cleaned of adherent tissue, and stored in −20°C before being scanned by microcomputerized tomography (μCT) scanner. The right femur of each mouse was also removed, cleaned of adherent tissue, flashed-frozen in liquid nitrogen, and stored at −80°C. The frozen femur was pulverized with a liquid nitrogen–cooled steel mortar and pestle in order to isolate total RNA as described below.
Detection of serum cytokines (IL-1β, IL-6, and TNF-α), osteocalcin, hAAT, and antibodies against hAAT
Serum IL-1β, IL-6, and TNF-α levels were quantified by murine enzyme-linked immunosorbent assay (ELISA) using the ELISA kit from Peprotech (Rocky Hill, NJ) according to manufacturer's instructions. Serum hAAT and anti-hAAT levels were detected by ELISA as previously described. 18 –20 Serum osteocalcin was detected using mouse osteocalcin high sensitive EIA kit from Takara Bio Incorporation according to manufacturer's protocol.
Real-Time PCR
Total extracted RNA from the right femur bone was purified using TRIzol reagent (Carlsbad, CA). Complementary DNA (cDNA) was synthesized from the total RNA using reverse transcriptase (QIAGEN) and subjected to real-time PCR as previously described. 21 Results were normalized with the gene expression levels of glyceraldehyde-3-phophate dehydrogenase (Gapdh) in the same sample. The fold-change ratios between test and control samples were calculated. The primers used in this experiments as follows: for RANK (TNFSF11A), 5′-GGT CTG CAG CTC TTC CAT GAC-3′ (Forward) and 5′-GAA GAG CAG AAC GAT GAG ACT-3′ (Reverse); for Integrin β3 (ITGB3), 5′-TTC AAT GCC ACC TGC CTC AAC AAC-3′ (Forward) and 5′-ACG CAC CTT GGC CTC GAT ACT AAA-3′ (Reverse); and for Gapdh, 5′-TGC ACC ACC AAC TGC TTA G-3′ (Forward) and 5′- GGA TGC AGG GAT GAT GTT C-3′ (Reverse).
Determination of bone microstructure by μCT
The 2nd lumbar vertebra (L2) from each mouse were cleaned of adherent tissue and placed in 10.2 mm in diameter holders and scanned by a Scanco μCT scanner (μCT-40; Scanco Medical AG, Bassersdorf, Switzerland) using a 12 μm isotropic voxel size with an X-ray source power of 55 kV and 145 μA, and an integration time of 300 milliseconds as described previously. 21 For the L2 vertebra analysis the entire secondary spongiosa between the cranial and the caudal area was scanned and analyzed. Bone histomorphometry nomenclature followed recommended standards. 22 Briefly, bone volume/total volume (BV/TV, %) is a ratio of segmented bone volume to the total volume of the region evaluated. Connectivity density (Conn.Dn, 1/mm3) is a measure of the degree of connectivity of trabeculae normalized by TV. Trabecular number (Tb.N, 1/mm) is a measure of the average number of trabeculae per unit length. Bone mineral density (BMD) is the amount of bone mineral in bone tissue. Trabecular thickness (Tb.Th) is the mean thickness of trabeculae. Structure model index (SMI) is an indicator of the plate- or rodlike geometry of trabecular structures.
Statistical analysis
Data were analyzed using one-way analysis of variance (ANOVA) using GraphPad Prism5 software, and Dunnett's multiple comparison test was used to compare statistical significance of treatment groups to the PBS-injected OVX group as a control. The data were expressed as mean ± standard error of the mean (SEM) and in all of the analysis, p < 0.05 was considered to be statistically significant.
Results
Recombinant AAV8 vector produced sustained levels of hAAT with negligible immune reaction
Cohorts of OVX mice were i.p. injected with the clinical grade of hAAT protein (Prolastin C®, 2 mg/mouse, every 3 days for 8 weeks), rAAV8-CB-hAAT (a single injection of 1 × 1011 viral particles/mouse) or PBS (as a control, every 3 days for 8 weeks). Serum hAAT and anti-hAAT levels were monitored by ELISAs. As expected, hAAT protein injection resulted in high serum levels of hAAT and detectable levels of anti-hAAT antibodies (Fig. 1A and B). Administration of rAAV8-CB-hAAT vector resulted in sustained levels of hAAT (Fig. 1C), while a negligible amount of anti-hAAT antibody was detected after rAAV8-CB-hAAT vector administration (Fig. 1D).
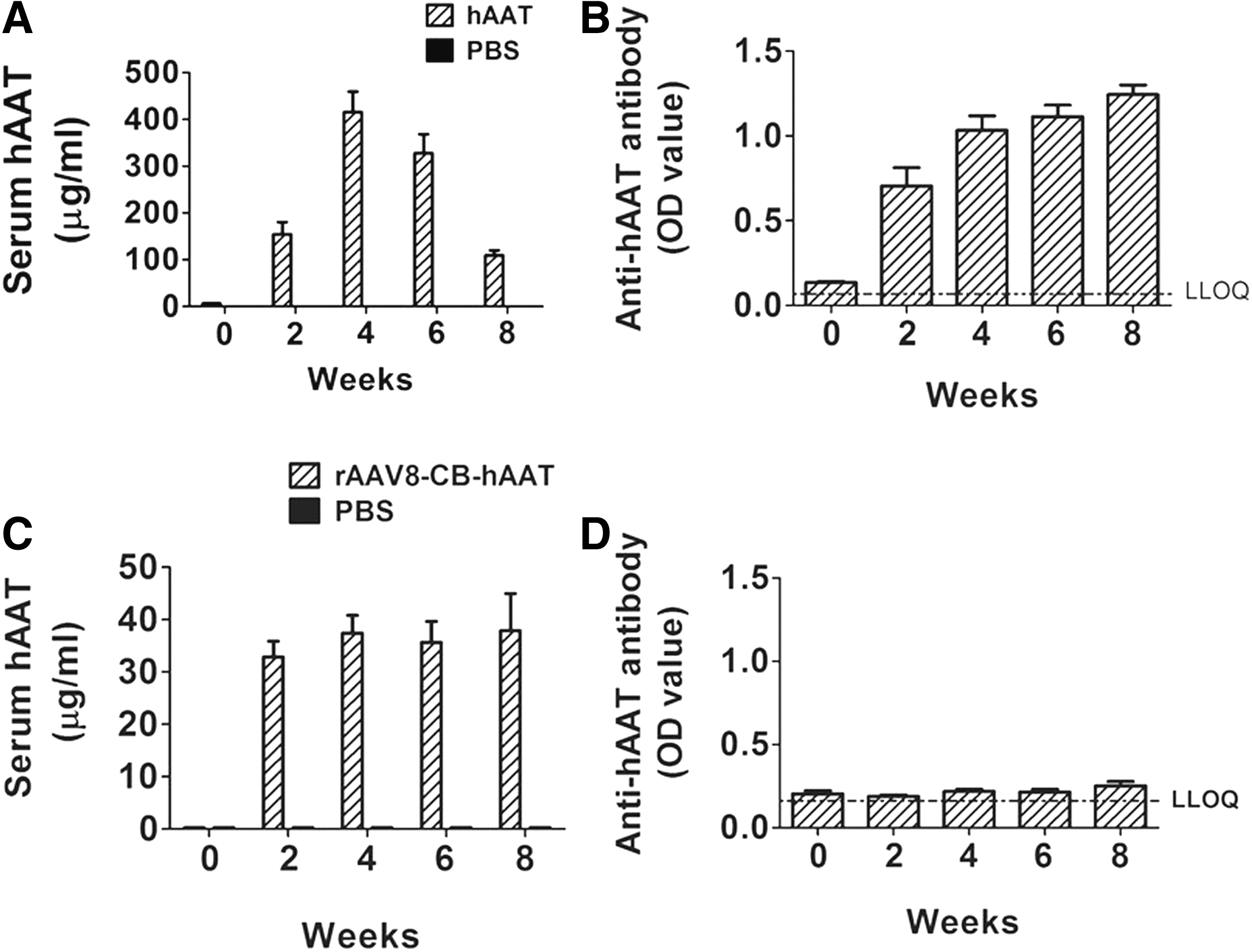
Serum levels of hAAT and anti-hAAT antibodies.
Human AAT-reduced serum levels of IL-1β, IL-6, and osteocalcin
Since proinflammatory cytokines including IL-1, IL-6, and TNF-α are primary mediators of the accelerated bone loss in both animals and humans, 1,23 we next investigated the effect of hAAT on proinflammatory cytokine levels induced by OVX or estrogen deficiency. As expected, our results showed that OVX caused significant increases of serum levels of IL-1β and IL-6 compared to the sham-operated group (Fig. 2A and B). Importantly, both hAAT protein and gene therapy decreased OVX-induced IL-1β and IL-6 levels, although there was no significant change of TNF-α (Fig. 2C). We also observed that hAAT treatment decreased serum osteocalcin levels, which was induced by OVX (Fig. 2D).
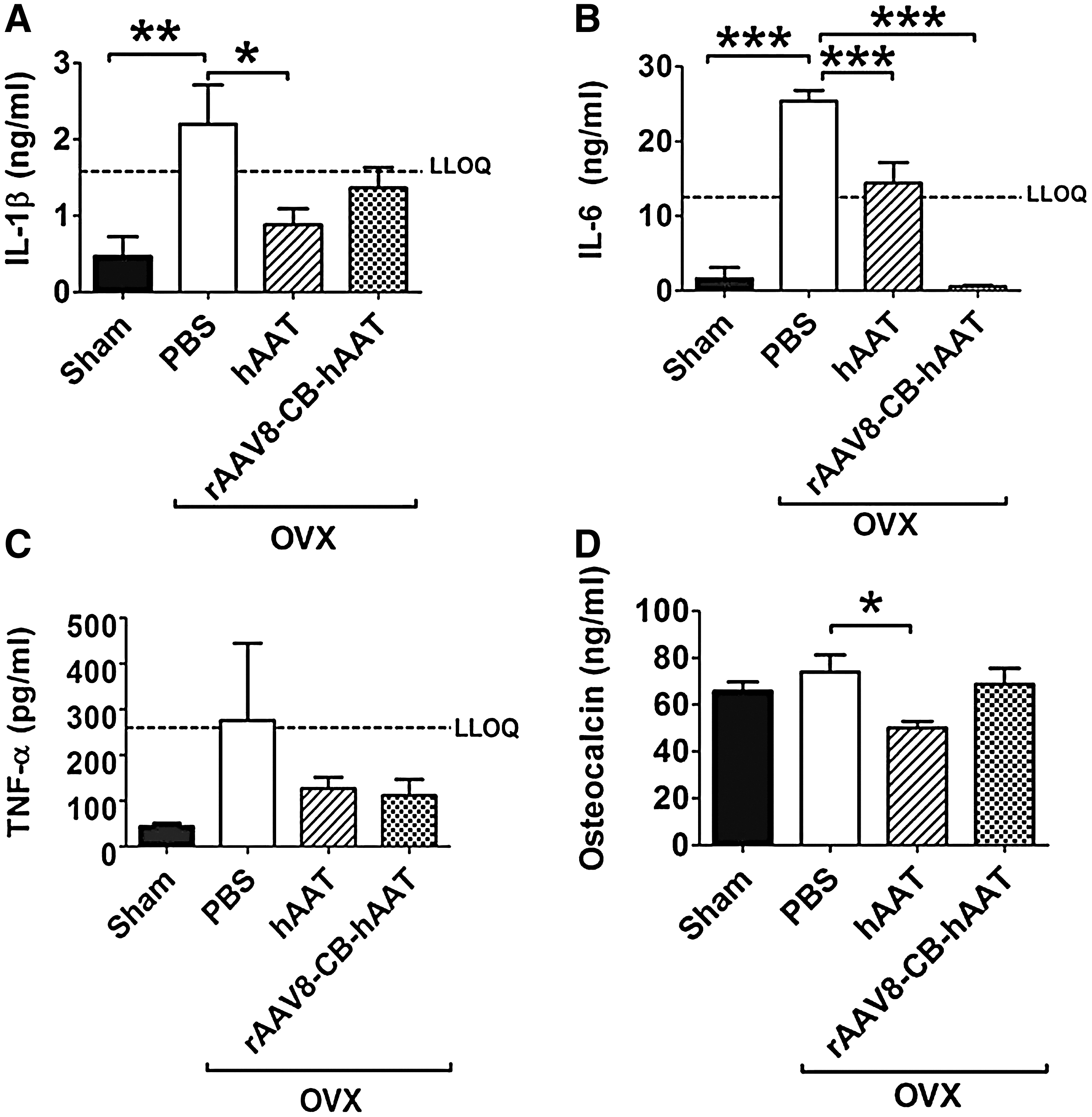
Serum levels of proinflammatory cytokines and osteocalcin detected by ELISA. Serum levels of
Human AAT down-regulated RANK and integrin β3 gene expressions
After 8 weeks of hAAT treatment or vector injection, all animals were sacrificed and examined the osteoclast-related gene expressions in bone tissue. Real-time PCR analysis revealed that hAAT protein and gene therapy completely abolished OVX-induced increase in RANK gene expression (Fig. 3A). We also observed that hAAT protein therapy significantly decreased OVX-induced integrin β3 gene expression (Fig. 3C). Although a statistically significant difference was not detected, hAAT protein and gene therapy groups showed a decreasing trend in the expression of RANKL and integrin β3 genes, respectively (Fig. 3B and C).
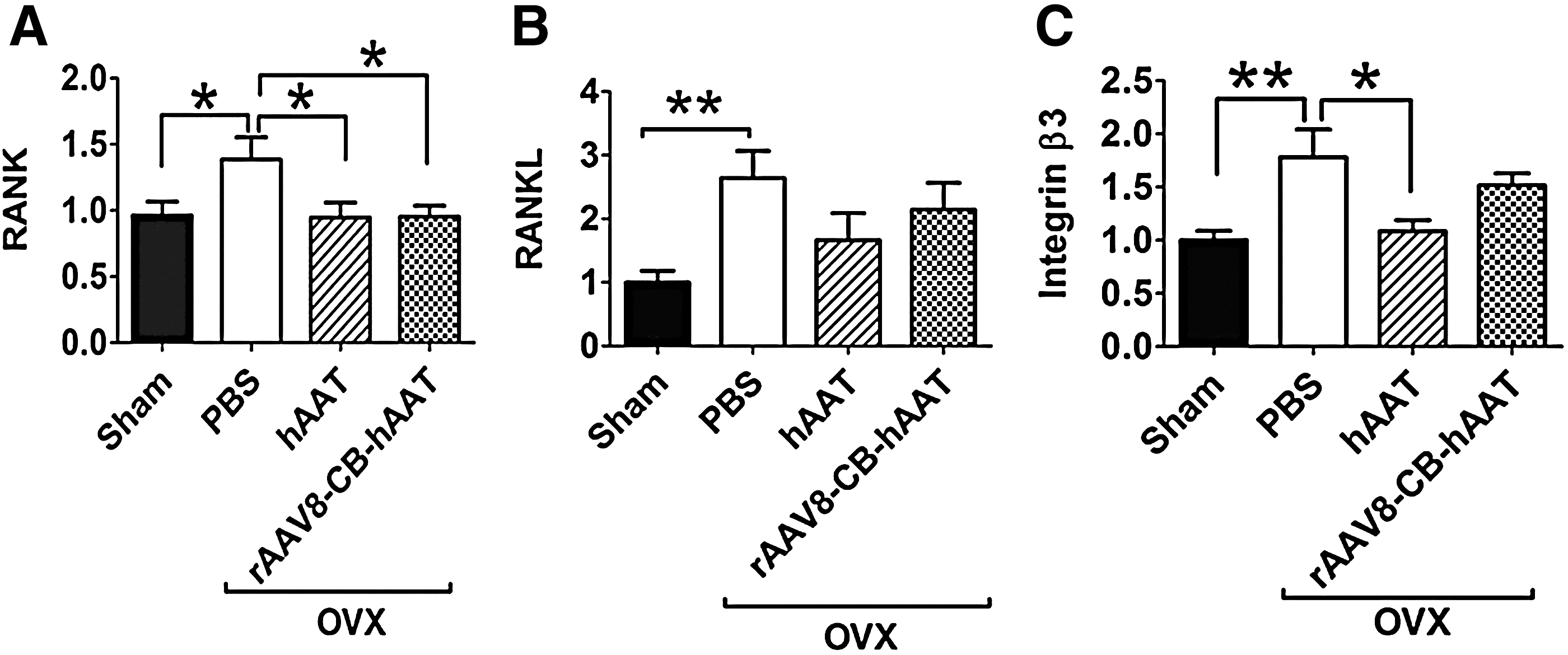
RANK, RANKL, and integrin β3 gene expression in bone tissue. Levels of mRNA were detected by quantitative real-time PCR. Fold induction of mRNA levels relative to sham-operated control groups were plotted.
Human AAT reduced OVX-induced bone loss
The second lumbar vertebra of each animal was subjected to μCT scanning for the evaluation of bone microstructure. As expected, OVX induced significant bone loss indicated by lowered bone volume/total volume (BV/TV), Conn.Dn (connectivity density), Tb.N (trabecular number), bone mineral density (BMD), Tb.Th (trabecular thickness), and higher SMI (structure model index) compared to sham-operated mice (Fig. 4A–F). These results are consistent with the previous observation that estrogen deficiency is detrimental to bone metabolism. 24 Human AAT gene therapy significantly increased BV/TV levels compared to that in PBS-injected OVX group, while hAAT protein therapy showed significant increase in BMD and an increasing trend in BV/TV. We showed that both protein and gene therapies significantly decreased OVX-induced SMI compared to PBS-injected OVX group. Gene therapy group appeared more protective (p < 0.001) than the hAAT protein therapy group (P < 0.05) relative to the PBS-injected OVX group.
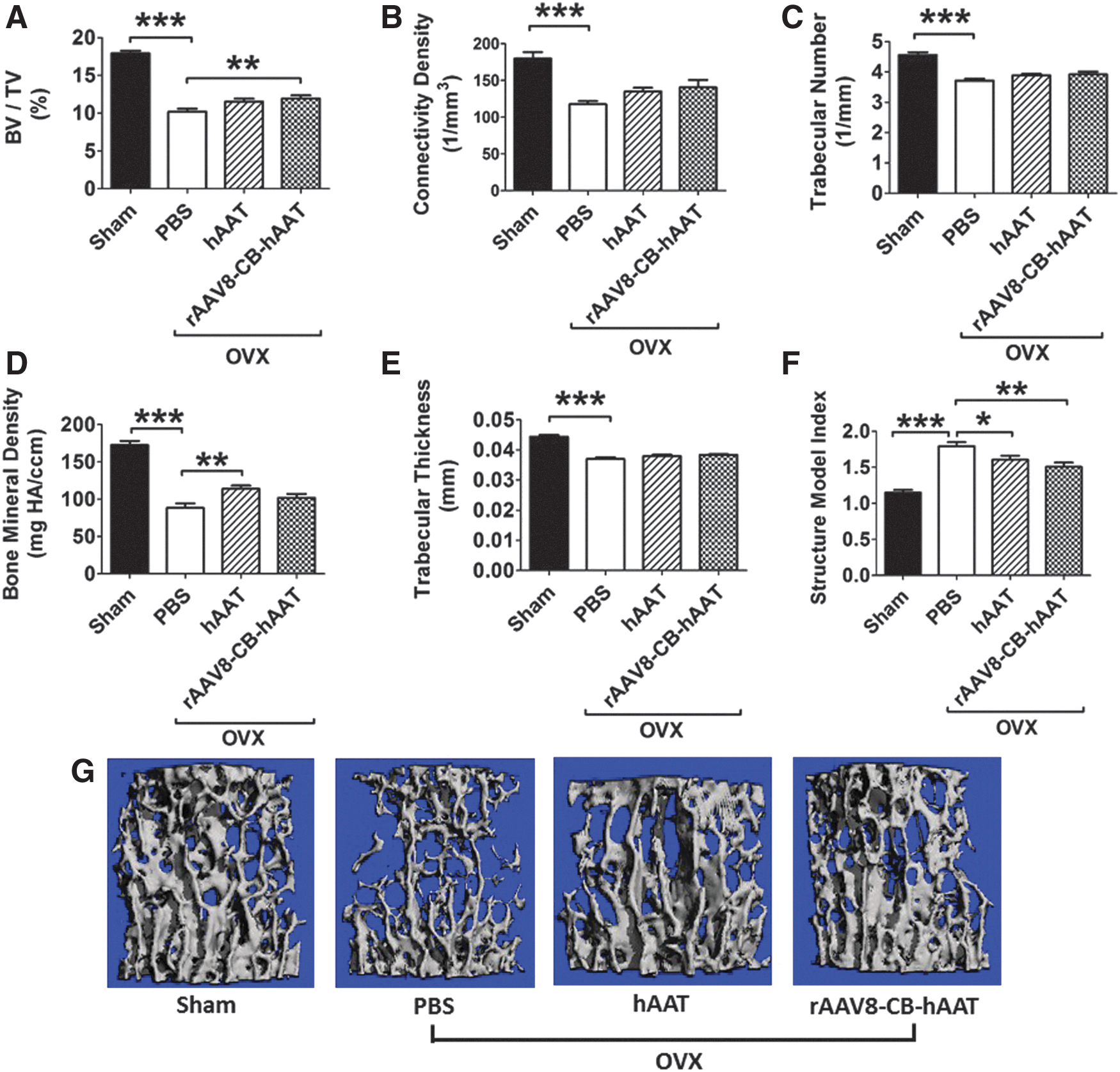
Effect of hAAT protein and gene therapies on vertebral bone structures:
Discussion
Our results show for the first time that hAAT gene therapy ameliorates bone loss in OVX mouse models. The protective effects of hAAT gene therapy were linked to reduction of serum levels of proinflammatory cytokine IL-6 and downregulation of the expression of RANK gene in bone. Since rAAV vector has been proven to be safe in humans, hAAT gene delivery by rAAV8-CB-hAAT vector could be a new strategy for the treatment of osteoporosis. This study also confirmed and expanded on our previous results indicating that injection of hAAT protects against OVX-induced bone loss. 25
Osteoporosis is a serious public health concern. Currently, no treatment can completely reverse established osteoporosis, and currently available drugs are associated with side effects. For example, bisphosphonates, a first line treatment option for osteoporosis, can cause osteonecrosis of the jaw, atypical fractures, and atrial fibrillation. 26 Therefore, a continued search for more efficient and safer treatment is warranted.
The exact mechanisms underlying the inhibitory effects of hAAT on OVX-induced bone loss is complex since hAAT is a multifunctional protein. First, the anti-proteinase activities (e.g., neutrophil elastase) may contribute to protect against bone tissue damage and inflammation. Second, inhibition of proinflammatory cytokines including IL-1β and IL-6 is clearly important in the protection of OVX-induced bone loss. For example, in premenopausal women, IL-1β levels increased after ovariectomy with a simultaneous increase in bone loss, and estrogen replacement therapy decreased IL-1β to preoperative levels. 27 Similar to the findings in humans, bone loss was reduced after treatment with IL-1 receptor agonist in mouse and rat OVX models. 28,29 Previous work done by Jilka et al. showed that ovariectomy caused a 10-fold increase in osteoclastogenesis, and administration of antibodies to IL-6 abolished this effect. 30 Another group showed that IL6-deficient mice maintained their bone mass following ovariectomy and did not experience a change in bone turnover rate, unlike the IL-6-producing wild-type controls. 31 In our study, hAAT treatments decreased IL-1β and IL-6 levels in OVX mice, which were consistent with previous observations in other disease models. 32,33 These results clearly showed that an anti-inflammatory effect of hAAT in the OVX mouse model could play an important role in reducing bone loss. Consistent with our previous study, we also observed an inhibitory effect of hAAT on serum osteocalcin levels, likely due to decreased bone turnover with hAAT treatment. Furthermore, we have shown that hAAT treatment inhibited OVX-induced RANK and integrin β3 gene expression, which play important roles in bone resorption. 34,35 Future studies dissecting each protective signaling pathway will uncover novel mechanisms.
In this study, we choose to use rAAV serotype 8 vector for its high efficiency and low immunogenicity. Numerous studies have shown that rAAV8 vectors can efficiently transduce many organs and tissues including heart, muscle, and liver. 36 Importantly, this vector can induce immune tolerance to the transgene product. 17 This unique feature is particularly useful in gene therapy for inflammation-associated diseases in which immune response to the transgene product (therapeutic protein) could complicate the results (therapeutic effect) in two ways. First, a strong immune response, which is a characteristic feature of these diseases, may partially reduce the therapeutic effect of gene therapy. Second, the immune response to the transgene product as a foreign antigen may shift the Th1 and Th2 balance with foreign antigen–induced nonspecific effect. Therefore, the immune response may complicate the specific therapeutic effect. We have previously shown that rAAV8-CB-hAAT vector mediated no or low immune response to hAAT in mouse models of type-1 diabetes and arthritis. 14,17 Consistent with these observations, results from the present study showed that rAAV8-CB-hAAT vector mediated nondetectable levels of anti-hAAT antibodies. Although other serotypes of rAAV vectors may also be used, the rAAV8 vector clearly holds greater advantage in gene therapy for chronic and inflammation-associated disease.
In our previous study, we evaluated bone structural changes of femurs at 4 weeks after hAAT treatment and showed that hAAT protein therapy improved BV/TV and Tb.Th. 25 Since femurs are weight-bearing bones that support the majority of the body weight in mice, the increase of body weight loading due to OVX may partially offset bone deterioration induced by estrogen deficiency. Therefore, the evaluation of femur bone may not be suitable for the assessment of long-term therapeutic effect of hAAT on bone. In the present study, we evaluated bone microstructure of vertebrae, which are not affected by body weight, and performed a relatively long-term experiment (8 weeks). Similar to previous findings, our results in this study demonstrated that hAAT treatment significantly improved bone structure including BV/TV, BMD, and SMI. Importantly this long-term experiment model allowed us to evaluate rAAV8-mediated hAAT gene therapy since the transgene expression needs at least 2 weeks to reach the maximal expression level in mouse model. 37
It is intriguing that a single i.p. injection of rAAV8-CB-hAAT resulted in the similar protection against bone loss compared with hAAT protein therapy. The protein therapy of hAAT resulted relatively high and variable levels of serum hAAT in the mice (100–400 μg/ml) and induced strong immune response to hAAT. Gene therapy with rAAV8-CB-hAAT, however, resulted in relatively lower but sustained serum levels of hAAT (30–40 μg/ml). Therefore, it appears that stable lower serum levels of hAAT could have the same bone protective effect as high but variable serum levels of hAAT. Future studies focusing on pharmacokinetics and pharmacodynamics of hAAT gene and protein therapies will optimize the protective effect.
Considering osteoporosis is an age-associated chronic disorder and requires long-term treatment, rAAV-mediated hAAT gene therapy may have several advantages compared to protein therapy: (1) avoiding frequent protein injection; (2) overcoming the potential shortage of human plasma purified hAAT; and (3) achieving more therapeutic effects. Therefore, rAAV vector–based hAAT gene therapy can be a unique treatment strategy for the treatment and/or prevention of osteoporosis.
Footnotes
Acknowledgments
This work was supported by grants from the University of Florida and the USDA Agricultural Research Service program “Food Factors to Prevent Obesity and Related Diseases,” Current Research Information System no. 3062-51000-053-00D. The U.S. Department of Agriculture, Agricultural Research Service, Plains Area is an equal opportunity/affirmative action employer, and all agency services are available without discrimination. Mention of trade names or commercial products in this article is solely for providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. Some of the data in this article were presented at ASGCT 19th annual meeting on May 6 in Washington DC.
Author Disclosure
No competing financial interests exist.