
Abstract
Recombinant adeno-associated viral (rAAV) vectors have recently achieved clinical successes in human gene therapy. However, the commonly observed, heavier particles found in rAAV preparations have traditionally been ignored due to their reported low in vitro transduction efficiency. In this study, the biological properties of regular and high-density rAAV serotype 8 vectors, rAAVRD and rAAVHD, were systemically compared. Results demonstrated that both rAAVRD and rAAVHD exhibited similar DNA packaging profiles, while rAAVHD capsids contained fewer VP1 and VP2 proteins, indicating that the rAAVHD particles contained a higher DNA/protein ratio than that of rAAVRD particles. Dynamic light scattering and transmission electron microscopy data revealed that the diameter of rAAVHD was smaller than that of rAAVRD. In vitro, rAAVHD was two- to fourfold less efficient in transduction compared with rAAVRD. However, the transduction performance of rAAVHD and rAAVRD was similar in vivo. No significant difference in neutralizing antibody formation against rAAVRD and rAAVHD was observed, suggesting that the surface epitopes of rAAVRD and rAAVHD are congruent. In summary, the results of this study demonstrate that rAAVRD and rAAVHD are equally competent for in vivo transduction, despite their difference in vitro. Therefore, the use of rAAVHD vectors in human gene therapy should be further evaluated.
Introduction
A
Recently, there have been a number of advancements in rAAV gene therapy for genetic diseases such as congenital blindness, 1,2 choroideremia, 3 α(1)-antitrypsin deficiency, 4 lipoprotein lipase deficiency, 5,6 and hemophilia. 7 Encouraging preliminary results are moving rAAV gene therapy closer to a routine clinical application. Although there have been continuous and significant progressions toward flexible and scalable rAAV production in recent years, 8 –13 a high demand still exists for more efficient vector production. This is partially due to the overwhelming amount of quality vectors needed to support preclinical research using large animal models and clinical trials. 14,15 Furthermore, low transgene expression in clinical trials has increased the demand for quality rAAV vectors in order to administer high-dose injections. 16
Naturally occurring AAV produced using adenovirus (Ad) as a helper can be fractionated by isopycnic centrifugation in cesium chloride (CsCl) into three predominant classes: defective interfering (DI)-AAV, regular AAV, and heavy AAV. 17 The most prominent visible band in the CsCl gradient contains DI-AAV, with a density of 1.32–1.35 g/cm3. These particles are referred to as defective interfering particles because they can interfere with the replication of regular and heavy AAV. They appear “empty” when examined using an electron microscope, and have been reported to contain either no DNA 18 or DNA of a truncated/incomplete genome. 19 Truncated/incomplete genomes will have deleted internal regions but retain the genome termini. 20 –22 The band containing regular AAV2 has a density of 1.41 g/cm3, whereas the band containing heavy AAV2 is 1.45 g/cm3. 17 It has been previously reported that heavy AAV particles were unstable in CsCl and present in a greater proportion early after infection. 17 In these studies, the ratio of regular and heavy AAV ranged from 1:1 to 1:3 at different time post infection. 17 The heavy AAV particles have also been identified in preparations of AAV1, 23 AAV3, 24 AAV4, 19 avian AAV, 25 and H-1 parvovirus. 26
The physical and biological properties of regular and heavy AAV particles have been previously investigated using different methods. Electron microscopy revealed that the heavy AAV2 was 10–20% smaller in diameter than the regular AAV2. 27 De la Maza and Carter found that both regular and heavy bands had the same DNA/protein ratio and the same density in metrizamide gradients. 17 Spectrophotometric analysis and velocity sedimentation of regular and heavy AAV2 particles failed to demonstrate any significant difference, and the DNA extracted from both regular and heavy AAV2 appeared to be similar. 17 However, contradictory reports have been published that reveal that heavy AAV2 has a higher DNA/protein ratio than regular AAV2 has. 24,27 It was also determined that the ratio of infectious units to physical particles of regular AAV2 is 16–300-fold higher than that of heavy AAV2 particles. 17,28 Other reports have found no significant differences in the infectivity between the regular and heavy particles of AAV4, the minute virus of mice (MVM), or the H1 parvovirus. 19,26,29 However, the AAV particles used in the aforementioned studies were all wild-type AAV, and did not address particles from rAAV preparations.
For the first time, the physical and biological properties of regular and high-density rAAV (rAAVRD and rAAVHD) vectors were characterized. This was accomplished through a comparison of genome content, capsid size and composition, in vitro and in vivo transduction efficacy, and potential antigenicity. Moreover, studies analyzing regular and high-density rAAV have not included the novel isolated serotypes, for example AAV8, 30 which has been widely used for liver and heart gene delivery. 31 –33 Defining the properties of rAAVRD and rAAVHD particles using AAV serotype 8, both in vitro and in vivo, will not only provide a unique insight into gene transduction, but also demonstrate their potential for use in gene therapy.
Materials and Methods
Construction of rAAV vector plasmids
The pAAV-hHCHL plasmid consisting of a 3.7 kb AAV genome carrying the heavy chain (hHC) of the human factor VIII (FVIII) gene was described previously. 34 The pdsAAV-Gluc (1.8 kb) and pdsAAV-EGFP (2.1 kb) plasmids were designed to carry either the Gaussia Luciferase gene (Gluc) or enhanced green fluorescent protein (EGFP) gene, driven by a human β-actin promoter with a CMV enhancer (CB). These two plasmids contained one full ITR and one mutant ITR, and were used to generate self-complementary AAV vectors (scAAV-EGFP and scAAV-Gluc).
Production and purification of rAAV vectors
All rAAV vectors were generated utilizing the triple plasmid, co-transfection method as previously described. 35 Briefly, pAAV-Rep&Cap (serotype 8), pAd helper, and the transgene plasmids were co-transfected into HEK 293 cells cultured in roller bottles at a ratio of 1:1:1. Vectors from both the transfected cells and medium were harvested 72 h post transfection, and purified by two rounds of CsCl gradient ultracentrifuge. The rAAVRD and rAAVHD particles were collected and exchanged extensively against phosphate-buffered saline (PBS) with 5% D-sorbitol. The density of the rAAVRD and rAAVHD particles was determined by collecting the vectors dropwise following the second round of CsCl gradient ultracentrifuge. The density of the fractions (refractive index: 1.380–1.362) was measured, and the capsid of the vectors were characterized by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and quantitative polymerase chain reaction (qPCR) analysis.
Characterization of rAAV capsid composition
The capsid composition of the rAAV vectors was determined by both silver staining and Western blot analysis. Briefly, rAAV vectors (∼1 × 1010 particles) were electrophoresed using a 10% SDS-PAGE gel. Following separation, the gel was stained using a commercially available silver staining kit (Pierce® Silver Stain Kit; Thermo Scientific, Rockford, IL). Western blot analysis was also performed as previously reported. 36 In short, an anti-AAV capsid antibody (B1; American Research Products, Belmont, MA) was diluted to a 1:500 ratio, and applied to a membrane containing the rAAV vectors. Next, a fluorescently labeled, secondary antibody (IRDye® 800CW Goat Anti-Mouse IgG [H + L]; LI-COR, Lincoln, NE) was diluted at a ratio of 1:1,000, and the membrane was applied. The resulting membrane was scanned using an Odyssey imaging system.
qPCR and Southern blot analysis of rAAV genomes
For titration of the rAAV vectors, a 10 μL aliquot of each vector was treated with Benzonase and proteinase K, and diluted to different ratios. To elucidate the genome copy number in the liver of injected mice, the total DNA was extracted from the liver cells using a GeneJET genomic DNA purification Kit (K0721; Thermo Scientific). All qPCR analysis was performed using the Fast SYBR Green Master Mix, which has been previously described, 35,37 using the following primers: hHC forward primer, 5′-CTGAAATGGATGTGGTCAGG-3′; hHC reverse primer, 5′-AGTCCCAGTCCTCCTCTTCA-3′; Gluc forward primer, 5′-CCACCGAGAACAACGAAGA-3′; Gluc reverse primer, 5′-CATCTCTTTGAGCACCTCCA-3′; EGFP forward primer, 5′-TGACCCTGAAGTTCATCTGC-3′; EGFP reverse primer, 5′-GAAGTCGTGCTGCTTCATGT-3′. The genome copy number of vector found in each sample was determined by comparison with a standard curve made from serial dilutions of plasmid containing the corresponding AAV genomes. A Southern blot analysis of genomes extracted from the vectors was performed using a 32P-labeled hHC probe made from a fragment of the plasmid pAAV-hHCHL digested using the enzymes AccI and KpnI.
Particle size measurement by dynamic light scattering and transmission electron microscopy
The diameter of the rAAV particles was measured using a particle analyzer (Zetasizer Nano ZS; Malvern Instruments, Westborough, MA). For measurement, a 735 μL (∼1 × 1012 vg) aliquot of vector in PBS solution was loaded into the disposable, low-volume cuvette and mounted in the dynamic light scattering (DLS) chamber. Each sample was measured three times, with 12 runs in each measurement. The average size of the particles was calculated from these multiple measurements in order to estimate the predominant viral particle size in the population. For transmission electron microscopy (TEM) analysis, formvar was coated on a microscope slide and floated onto a water bath. Cleaned 200 mesh grids were placed on this film and collected using parafilm. Five microliters of purified AAV (∼1 × 1013 vg/mL) was placed on the grid and allowed to dry, then 8 μL of 1% phospho tungstic acid (Electron Microscopy Science, Hatfield, PA) was applied and drawn off. After drying, the grids were observed using a Philips Transmission Electron Microscope CM 12 (Philips, Eindhovern, Netherlands) with an accelerating voltage of 100 KV and imaged with a DVC detector controlled by AMT software (Danvers, MA). The diameter of the rAAV particles was measured using Image J software.
rAAV transduction in vitro
The cell lines used in this study—Hela, CV1, BHK, HEK 293, GM16095, and Cos-1—were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS; HyClone, Logan, UT), penicillin (100 IU/mL), and streptomycin (100 μg/mL) at 37°C in a moisturized environment supplied with 5% CO2. For each transduction experiment, 50,000 viable cells were seeded in a 24-well plate 24 h prior to transduction. Regular and heavy AAV particles of scAAV8-EGFP or scAAV8-Gluc, with or without Ad5, were added directly to each well. EGFP expression was observed 24 h post infection using a fluorescent microscope. Gaussia luciferase(Gluc) expression from the media was determined by adding 5 μL of coelenterazine substrate (CLZ, 200 μg/mL) in 0.1 M of Tris buffer (pH 7.5) containing 0.5 M of NaCl. Bioluminescence was measured with a POLARstar Omega plate reader (BMG Labtech, Ortenberg, Germany). All experiments were performed in triplicate, and the results are presented as averages of the experimental data.
rAAV transduction in vivo
In vivo transduction experiments were carried out using 6–8-week-old BALB/c male and C57BL6/Svj129S female hemophilia A (HA) mice. All mice were housed in a specific pathogen-free environment, supplied a normal diet, and treated in accordance with National Institutes of Health guidelines and as approved by IACUC at Temple University (ACUP 4142). During all procedures, the mice were anesthetized using isoflurane (induction 3–5%, maintenance 1–3%) and monitored every 5 min for absence of limb withdrawal from toe pinch, absence of the eye-blink reflex, lowering of the heart or respiration rate, and absence of muscle tone. Post procedure, the animals were monitored every 15 min until ambulatory. Multiple doses of scAAV8-Gluc vector were injected into mice via the tail vein as previously described. 35 For these injections, the high dose was 6 × 1011 vg/mouse and the low dose was 2 × 1011 vg/mouse. Mouse plasma was harvested post vector administration by retro orbital eye bleeding at regular intervals as described. Plasma was diluted in 0.1 M of Tris buffer (pH 7.5) containing 0.5 M of NaCl, and Gluc expression was measured following the in vitro protocol. To terminate the experiment, mice were sacrificed by asphyxiation from inhalation of carbon dioxide followed by cervical dislocation. DNA from the mouse liver was extracted using a GeneJET genomic DNA extraction Kit for qPCR analysis.
rAAV neutralization assay
To determine the neutralizing antibody (NAb) response from mice injected with rAAVRD and rAAVHD vectors, the NAb-mediated inhibition of rAAV transduction was measured in vitro. Briefly, the sera of mice injected with either rAAV8-GlucRD or rAAV8-GlucHD were collected 6 months post injection. BHK cells were seeded in a 96-well plate at a density of 5 × 104 cells/well in 100 μL of DMEM, supplemented with 10% FBS, 4 h before inoculation. The inoculum contained rAAV-GlucRD or rAAV-GlucHD (2 × 108 particles) preincubated with 1:100 diluted mouse anti-serum for 2 h at 4°C in a total volume of 100 μL. Twenty-four hours after inoculation, Gluc expression was measure following the in vitro Gluc assay. A total of 50 human serum samples (Division of Human Genetics, Cincinnati Children's Hospital Medical Center, Cincinnati, OH) were included to study the effects of NAbs against rAAV8-GlucRD and rAAV8-GlucHD further. Serum samples at 1:100 dilution that inhibited rAAV transduction by ≥50% compared with normal mouse serum were considered as positive.
Statistical analyses
Both the two-tailed Student's t-test and one-way analysis of variance with Bonferroni multiple comparisons post test were performed. The differences were considered significant when the p-value was <0.05. The analysis was performed using SPSS v11.0 (SPSS, Inc., Chicago, IL).
Results
Comparison of the physical properties of rAAVRD and rAAVHD
It has been previously demonstrated that both regular and heavy AAV particles are present in wild-type AAV preparations when Adenovirus is used as a helper. 17 Accordingly, both particles are also found in rAAV vector preparations, as are DI particles. Fig. 1 demonstrates the presence of these three particles in a preparation of rAAV8 vector containing a 3.7 kb AAV genome. Fig. 1A illustrates the three particles with different buoyant densities. Using silver staining analysis of each fraction, the presence of three peaks corresponding to the three bands observed in the CsCl gradient was confirmed. They were identified as defective interfering rAAV (rAAVDI), rAAVRD, and rAAVHD, having buoyant densities of 1.33, 1.39, and 1.43 g/cm3, respectively (Fig. 1B).

Physical properties of recombinant adeno-associated viral (rAAV) particles in cesium chloride (CsCl) gradient. (
In order to explain the density discrepancy between the rAAVRD and rAAVHD vectors, the size of the packaged genomes was evaluated. All of the vectors were treated with benzonase to remove any unprotected nucleic acid before proteinase K treatment was applied to release the genomes. Fig. 1C demonstrates the major DNA band, 3.7 kb in length, for both the rAAVRD and rAAVHD vectors. This band represents the product of annealed, plus and minus genome strands, which corresponds to the size of the AAV genome of the pAAV-hHCHL plasmid. The Southern blot analysis confirmed that the packaged DNA for both the rAAVRD and rAAVHD vectors was similar (Fig. 1D). Analysis of DNA from self-complementary vectors (scAAV8-Gluc, 1.8 kb) also supported that the regular and heavy particles packaged the same size genomes (data not shown).
Gel electrophoresis and Southern blot analyses indicated that the packaged DNA genome did not significantly contribute to the difference in density between the regular and heavy particles. Next, the capsid composition of the two vectors was examined in order to delineate the different physical properties of the two particles. Fig. 1E provides the results of SDS-PAGE analysis of the protein components for rAAVRD and rAAVHD. All three structural capsid proteins were present in both rAAVRD and rAAVHD particles. Furthermore, there is evidence of the presence of another protein (VP1.5) in addition to VP1, VP2, and VP3 in both vectors. The VP1.5 protein may be the result of a VP2 variant that occurred due to a G/C change at position 219 introducing a novel CTG start codon between VP1 and VP2. 38 Although the rAAVHD particles contained all the same classes of VP proteins, when compared, the relative amounts of VP1, VP1.5, and VP2 were less than rAAVRD. The ratio of VP1:VP1.5:VP2:VP3 was approximately 1:1.5:1:4.5 and 1:1:2:8 in rAAVRD and rAAVHD, respectively. This result was confirmed by Western blot analysis using the B1 antibody that specifically targets the AAV VP polypeptides (Fig. 1F). The phenomenon that rAAVHD had less VP1, VP1.5, and VP2 was also observed with other heavy particles containing different transgenes (data not shown).
rAAVHD particles are smaller than rAAVRD particles
As rAAVHD particles contain less VP1, VP1.5, and VP2, it is reasonable to suspect that the size of the heavy particles is smaller than the regular particles. Therefore, a DLS experiment was performed to investigate the size differences of the regular and heavy particles. Results of this experiment determined that rAAVHD had a size distribution ranging from 18.2 to 91.3 nm, whereas the size of rAAVRD ranged from 15.9 to 105.7 nm (Fig. 2A). The rAAVHD peaked at 35.2 ± 0.4 nm, which is less than the rAAVRD, which peaked at 37.8 ± 0.7 nm (Fig. 2C). Similarly, the average size of rAAVHD was 38.2 ± 0.6 nm, whereas the average size of rAAVRD was 41.2 ± 0.9 nm. Interestingly, a ∼700 nm peak was observed with the rAAVHD vectors. It likely represents large capsid aggregates, and was excluded during the average size calculation. Both rAAVRD and rAAVHD were slightly elliptic particles, close to round, when they were detected by TEM, with the mean length of the major axes being 26.1 ± 3.0 nm and 24.8 ± 3.9 nm, respectively (Fig. 2B, C, and D). Therefore, the rAAVHD was approximately 5–7% smaller in diameter than the rAAVRD, in agreement with what was observed using AAV2. 17,28 Equal amounts of AAV particles (quantified based on qPCR results) were analyzed by silver staining after separation using a SDS-PAGE gel. The results demonstrated that rAAVRD and rAAVHD had a similar amount of VP3 protein but that rAAVHD particles had less VP1, VP1.5, and VP2. Therefore, it was concluded that the DNA/protein ratio of heavy particles was higher than that of regular particles, and the total mass (DNA + Capsid) of rAAVHD was less than rAAVRD. As the rAAVHD was more compact than rAAVRD (1.43 g/cm3 and 1.39 g/cm3), it can also be speculated that rAAVHD is smaller than rAAVRD. This observation is supported by size measurement using DLS and TEM technology. However, due to the limitations of DLS and TEM, the actual physical size of the rAAVHD particles will need to be further quantified in the future.
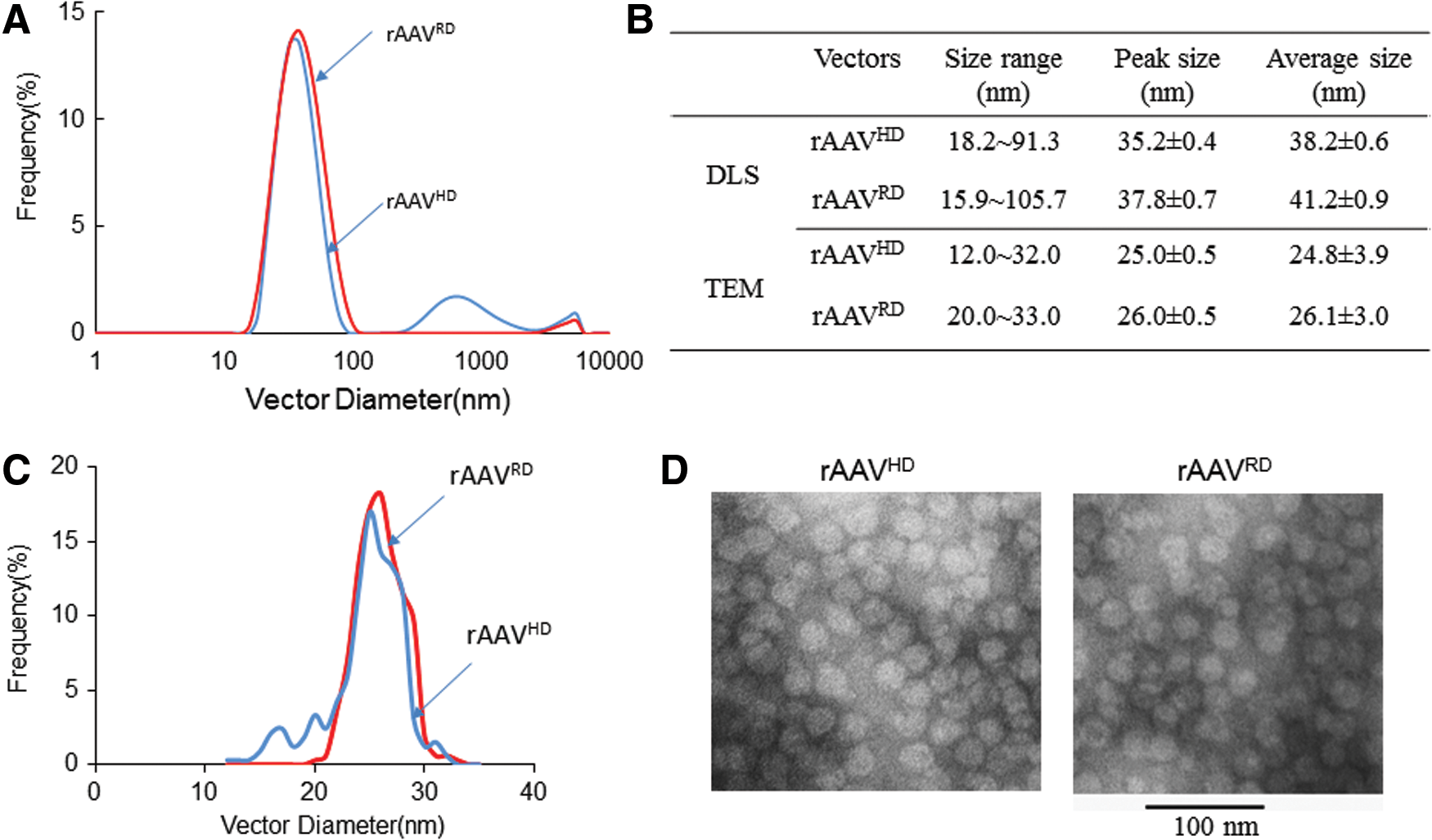
Size distribution of rAAVRD and rAAVHD particles. (
Different in vitro transduction efficiency between rAAVRD and rAAVHD
It is well known that the N-termini of VP1 contains a phospholipase A2 domain (PLA2), nuclear localization signals (NLSs), and several protein sequence motifs (PSMs) that are necessary for efficient infection in vitro. 39 –44 Therefore, since it was determined that the rAAV8HD particles contain less VP1, VP1.5, and VP2, it is logical to hypothesize that they will also have a lower transduction efficiency compared with rAAV8RD. Using the viral preparations described above, the transduction efficiency of rAAVRD and rAAVHD vectors was characterized in the different cell lines HEK 293, BHK, GM16095, Hela, CV1, and Cos-1. The transduction efficiency of the two vectors was measured using EGFP (scAAV8-EGFP) and Gluc (scAAV8-Gluc) as reporter genes. The EGFP and Gluc expression patterns were examined 24–72 h post infection. As illustrated in Fig. 3A, the EGFP positive cells were significantly lower in cultures infected with scAAV8-EGFPHD than those infected with scAAV8-EGFPRD. Among the cell lines tested, BHK produced the highest EGFP expression. Similar results were observed when Gluc was used as reporter; AAV8HD transduction was fourfold less efficient than rAAV8RD in 16095 cells (Fig. 3B). In other cell lines, there was a calculated twofold difference between the rAAV8RD and rAAV8HD transduction. These results indicate that the difference in transduction efficiency between rAAVRD and rAAVHD is less than what was previously reported using an AAV2 infection, which demonstrated a 10–100-fold difference. 17 Previous reports included the co-infection of Ad. Therefore, the effect of Ad on rAAVRD and rAAVHD transduction was examined. It was demonstrated that the co-infection of Ad dramatically potentiated transduction of both rAAV vectors (Fig. 3C and D). However, Ad increased the transduction of the rAAVHD and rAAVRD by the same degree. Together, these results suggest that rAAVRD and rAAVHD particles only produce a moderate difference in transduction activity in vitro.

Transduction efficiency of rAAVHD and rAAVRD particles in vitro. Different cell lines including HEK-293, HeLa, Cos-1, BHK, GM16095, or CV1-T cells were infected with the rAAVHD or rAAVRD vectors (50,000 vg/cell). (
Similar in vivo transduction efficiency of rAAVRD and rAAVHD
In previous studies, the infection activity of regular and heavy AAV particles was only compared using in vitro conditions 23 ; the relative transduction efficiency of rAAVRD and rAAVHD vectors was never evaluated in vivo. In this study, BALB/c male HA mice were injected with 2 × 1011 vg of either scAAV8-GLucHD or scAAV8-GlucRD vectors via the tail vein (N = 6). Since heavy particles have two- to fourfold less activity in vitro, a high-dose group (6 × 1011 vg/mouse) of heavy particles was included. Every 2 weeks post injection, the luciferase activity was quantified. The relative luciferase values for both vectors were similar (Fig. 4A). The high-dose group of heavy particles produced a two- to threefold increase in activity compared with the regular and heavy particle low-dose groups. In order to confirm these results, the same vectors were administered intravenously to 4–8-week-old C57BL6/Svj129S female HA mice (N = 6). Similar to the previous observation, the scAAV8-GlucRD and scAAV8-GlucHD produced comparable Gluc expression (data not shown). Taken together, these data suggest that rAAV8HD vectors are as efficient as rAAV8RD vectors in vivo, which is opposite of the in vitro results.
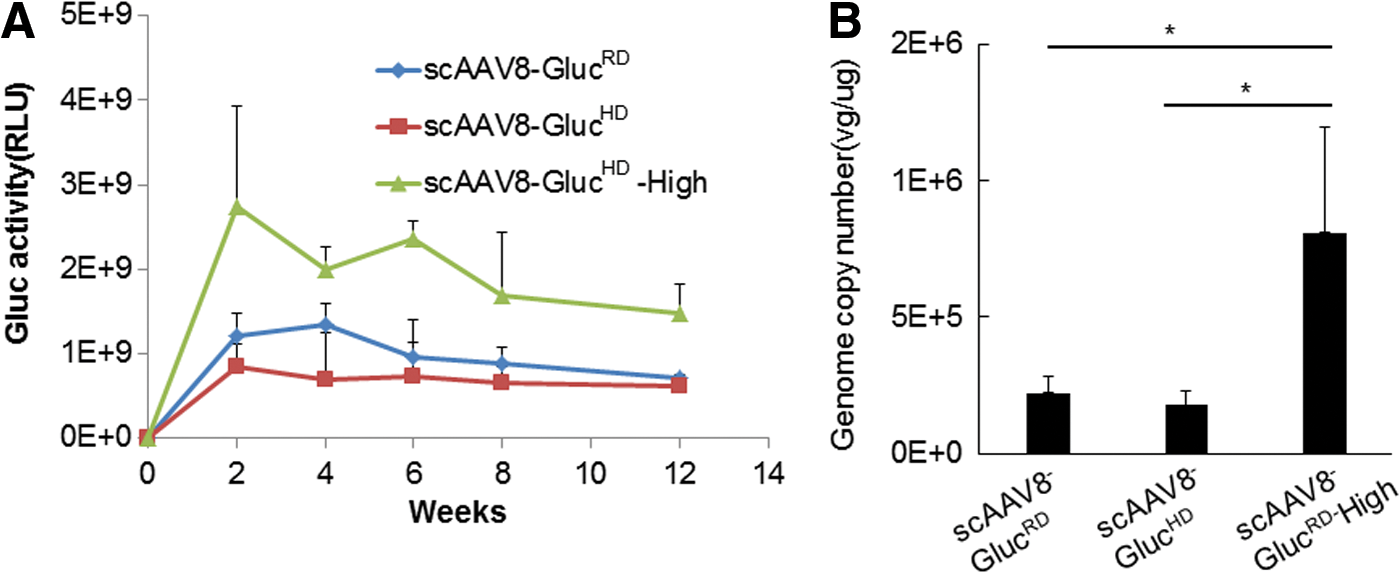
rAAVRD and rAAVHD achieved equal transduction in vivo. Multiple doses of scAAV8-Gluc vectors (high dose: 6 × 1011 vg/mouse; low dose: 2 × 1011 vg/mouse) were injected into C57BL6/Svj129S female mice via the tail vein. (
Next, the viral genome copy numbers remaining in the tissue samples were compared using qPCR (Fig. 4B). In the low-dose group, the viral genome copy number in the liver was approximately 2 × 105 vg/μg. There was no significant difference between mice injected with scAAV8-GLucHD or scAAV8-GlucRD vectors. The viral genome copy number in the high-dose group of scAAV8-GLucHD was four times higher than that in the low-dose group, which is consistent with the input amount of vectors. This result further demonstrates that the transduction efficiency and genome incorporations of rAAVRD and rAAVHD vectors are similar.
NAb responses of rAAVRD and rAAVHD particles
To define the antigenic properties of rAAVRD and rAAVHD vectors, their reactions to anti-AAV sera were tested. For this assay, the sera from scAAV8-GlucRD-injected HA mice was used. BHK cells were plated 24 h before infection with rAAV vectors at a multiplicity of infection of 5,000. The scAAV8-GlucHD and scAAV8-GlucRD vectors were incubated with anti-AAV sera at a dilution of 1:100 before infection. The control groups were incubated with normal mouse sera at the same dilution. Gluc activity was determined 24 h post infection. The results of this experiments demonstrated that all of the mice developed anti-AAV8 NAbs (Table 1). NAbs induced by the scAAV8-GlucRD vector dramatically inhibited the transduction capability of both scAAV8-GlucRD and scAAV8-GlucHD vectors. Similar results were found when sera from mice injected with scAAV8-GlucHD were tested, with 5/6 mice developing anti-AAV NAbs to both vectors (Table 1). When scAAV8-GlucRD and scAAV8-GlucHD vectors were incubated with different dilutions of anti-AAV sera, similar inhibition was observed (data not shown). These results indicate that the antigenic properties of rAAV8RD and rAAV8HD are similar. As the NAb reaction may vary between mouse strains, the same experiments were performed using sera of Balb/c mice injected with either scAAV8-GlucRD or scAAV8-GlucHD. Again, the transduction of both scAAV8-GlucRD and scAAV8-GlucHD was inhibited by the sera of mice injected with either vector.
NAbs reaction difference between rAAVRD and rAAVHD particles in BALB/c male HA and C57BL6/Svj129S female HA mice
NAbs, neutralizing antibodies.
To analyze further and to provide a translational implication of the NAb response of rAAVRD and rAAVHD, 50 samples of human sera were collected and incubated with either scAAV8-GlucRD or scAAV8-GlucHD. The results demonstrated that NAbs for both scAAV8-GlucRD and scAAV8-GlucHD were present in nine human samples (Table 2), and that the NAbs neutralized both rAAV8RD and rAAV8HD in a similar fashion. This indicates that the rAAV8RD and rAAV8HD vectors are capable of generating cross-reactive immune responses, suggesting that the two types of rAAV8 vectors harbor similar capsid proteins on their surface and process similar antigenicity.
Prevalence of NAbs against rAAVRD and rAAVHD in human serum samples
Discussion
The results of this study demonstrate that both regular and heavy bands of rAAV8 after CsCl purification contain infectious rAAV particles, which parallels previous results obtained using wild-type AAVs and other parvoviruses. 17,23,25,26,29,35 The AAV particles purified from these bands were reported to have significantly different infectious properties in vitro. De la Maza and Carter found a 16-fold difference in infectivity between the regular and heavy AAV2 particles, 17 whereas Roy et al. reported a 300-fold difference in infectivity. 28 In contrast, another previous study of AAV4 proposed that the infectivity of heavy AAV4 was almost the same as regular AAV4. 19 In the current study, a two- to fourfold difference in transduction activity was demonstrated between rAAV8RD and rAAV8HD particles in vitro. This difference in transduction activity was confirmed in six different cell lines.
Previously, Clinton and Hayashi demonstrated that regular and heavy particles from MVM were equally infectious after maximum adsorption was reached, but that the heavy particles had a slower rate of adsorption. 29 Paradiso found that the regular and heavy H1 parvovirus particles had similar particle/infection ratio. 26 For MVM and H-1, there is evidence indicating a precursor–product relationship between the regular and heavy density particles. However, a subsequent study did not demonstrate a precursor–product relationship between regular and heavy AAV particles. 17 Therefore, the dissimilarity between AAV and other parvoviruses may explain the activity difference between rAAVRD and rAAVHD particles. It is suggested that the discrepancy between the work presented here and that of others may be due to the contribution of Ad. Previously it had been proposed that Ad exerted multiple effects on AAV transduction. 45 –47 Co-infection of Ad with rAAVHD and rAAVRD in Hela and GM16095 cells dramatically potentiated transduction (Fig. 3). However, Ad increased the transduction of the rAAVRD and rAAVHD to a similar degree. The decreased transduction activity of rAAVHD in vitro is not due to its genome, as agarose gel and Southern blot analysis of DNA from both rAAVHD and rAAVRD did not indicate any genomic differences. The present results are consistent with previous findings showing no difference in genomic DNA using velocity sedimentation and restriction endonuclease digestion analyses. 17
The capsid compositions may contribute to the difference in transduction activity between rAAVRD and rAAVHD. Normally, AAV's capsid is composed of the three structural proteins: VP1, VP2, and VP3. For AAV2, VP1 and VP2 differ from VP3 by an N-terminal extension of 65 amino acids, 39 –44 and VP1 contains an additional 137 unique amino acids. The N-termini of VP1 is buried within the capsid interior but becomes externalized during the passage of AAV through the endosomal compartment. It has been previously determined that the N-termini of VP1 contains a PLA2, several NLSs, and PSMs that are necessary for efficient infection in vitro. These elements are conserved among all parvoviruses, and mutation of these sequences decrease infectivity in vitro. 40 As presented in Fig. 1, the rAAV8 has an additional capsid protein (VP1.5) as well as VP1, VP2, and VP3 for both regular and heavy particles. VP1.5 is 72 amino acids shorter than VP1. 38 The novel capsid composition observed in rAAV8 still retains normal structure and functional activity. 48 Since results consistently demonstrated that rAAV8HD has less VP1 and VP1.5 than rAAV8RD, it is logical to conclude that the low transduction activity of rAAVHD particles may be due to the presence of VPs that contain less PLA2, NLSs, and/or PSMs. The mutant BR3_K, which contains an alanine-to-lysine substitution in the third basic region of VP1, was 10- to 100-fold-less infectious than rAAV2 in transformed cell lines. In contrast, it produced only a twofold difference in several nontransformed cell lines, such as CV1 and C2C12. 40 However, in current study, the performance of rAAVRD and rAAVHD in non-transformed cell lines and transformed cell lines is very similar.
Although rAAVHD was less efficient than rAAVRD in transduction in vitro, its infectivity was indistinguishable from rAAVRD in vivo (Figs. 3 and 4). The underlying cause of infectivity differences between in vitro and in vivo testing remains unclear. The mutant BR3_K was less infectious than AAV2 in vitro, but not in vivo. 40 This is similar to what was previously documented for AAV2 and AAV8. AAV2 can more efficiently transduce cells in tissue culture compared with other serotypes, including AAV8. However, AAV8 consistently achieves a 10- to 100-fold improvement in gene transfer efficiency over AAV2 in the liver. 30 Moreover, the preclinical animal studies on rAAV transduction efficiency are not always predictive of the human outcome. It is well known that rAAV8 vectors are very effective in many animal models, including non-human primates. However, human hepatocytes are ∼20 times less efficiently transduced compared with mouse hepatocytes. 31 This phenomenon indicates that current knowledge of AAV transduction is limited. Therefore, it is important to test AAV-based vector transduction utilizing a variety of targets in vivo.
Humoral immunity against AAV vectors represents an important barrier to intravascular gene transfer, resulting in the clearance of the vector before it enters the target cell. 49 Antibodies directed against the AAV capsid are highly prevalent in humans, a natural host for this virus, and cross-react with a wide range of serotypes because of the degree of homology between the capsid proteins. As a result, even relatively low titers of NAbs can block AAV transduction when the vector is introduced into the bloodstream. Studies in humans have found that antibodies to some AAV serotypes are less prevalent than others. 50 AAV serotype switching and capsid engineering strategies have produced promising results. 51,52 Because the capsid composition is different between rAAVRD and rAAVHD particles, it can be hypothesized that humoral immunity may also have some difference. Immunoelectron microscopy studies indicated that rAAVRD and rAAVHD may exhibit some antigenic differences. 53 This study investigated the humoral immunity of rAAV in two ways. First, HA mice were injected with rAAVRD and rAAVHD vectors. Then, the sera from injected mice were incubated with rAAVRD and rAAVHD vectors. However, no difference was found between rAAVRD and rAAVHD NAbs, confirming that both NAbs neutralized the regular and heavy particles equally. Second, NAbs against rAAVRD and rAAVHD were screened using healthy subjects. Again, there was no significant difference in the prevalence of NAbs against either rAAVRD or rAAVHD particles. Thus, rAAVHD failed to display an altered antigenic profile compared to rAAVRD. This is a reasonable outcome, since the differences between rAAVRD and rAAVHD particles are the amounts of VP1, VP1.5, and VP2, which may not change the outside of the capsid but may change the interior.
Collectively, the work herein is the first report providing evidence that the rAAVHD is as efficient as rAAVRD for in vivo gene delivery, and there is no difference in NAb response to these two kinds of rAAV particles.
Footnotes
Acknowledgments
This work was supported by NIH grants (R01HL080789, R01HL114152, and HL130871) and the Natural National Science Foundation of China (81271691,81371669, 81371672).
Author Disclosure
No competing financial interests exist.