
Abstract
Cell therapies are treatments in which stem or progenitor cells are stimulated to differentiate into specialized cells able to home to and repair damaged tissues. After their discovery, endothelial progenitor cells (EPCs) stimulated worldwide interest as possible vehicles to perform autologous cell therapy of tumors. Taking into account the tumor-homing properties of EPCs, two different approaches to control cancer progression have been pursued by combining cell-based therapy with gene therapy or with nanomedicine. The first approach is based on the possibility of engineering EPCs to express different transgenes, and the second is based on the capacity of EPCs to take up nanomaterials. Here we review the most important progress covering the following issues: the characterization of bona fide endothelial progenitor cells, their role in tumor vascularization and metastasis, and preclinical data about their use in cell-based tumor therapy, considering antiangiogenic, suicide, immune-stimulating, and oncolytic virus gene therapy. The mixed approach of EPC cell therapy and nanomedicine is discussed in terms of plasmonic-dependent thermoablation and molecular imaging.
Introduction
T
EPC Description and Characterization
New blood vessel formation has long been considered to result from preexisting vessels, a process referred to as angiogenesis. Identification of EPCs in peripheral blood has highlighted an alternative mechanism based on EPC recruitment from bone marrow (BM), a process named vasculogenesis. The term EPC stands for various cell types able to migrate, proliferate, and acquire specific markers of mature endothelial cells under specific environmental conditions. Since the first identification of BM-derived EPCs by Asahara and colleagues, 1 the definition of true endothelial progenitors is still controversial and the absence of specific markers has been integrated with functional parameters, including cell morphology and clonal proliferation, replating ability, and ability to generate tubelike structures in vitro and vessels in vivo. Using various culture methods, three populations of putative EPCs have been identified, as overviewed by Basile and Yoder 2 : circulating angiogenic cells (CACs), colony-forming unit Hill (CFU-Hill) cells, and endothelial colony-forming cells (ECFCs).
CACs, 3 also called early outgrowth EPCs 4 or proangiogenic hematopoietic cells (PACs), 5 are derived from human peripheral blood mononuclear cells (PBMCs) maintained in culture plates coated with fibronectin for 5–7 days. Adherent cells take up acetylated low-density lipoprotein (acLDL) and bind Ulex europaeus agglutinin-1, both considered hallmarks of endothelial cells. However, these cells do not form colonies in vitro and do not create, in vivo, de novo vessel-like structures. CACs contribute indirectly to the formation of blood vessels in vivo by homing to damaged tissues and secreting angiogenic cytokines.
CFU-Hill cells 4,6 or CFU-ECs are early outgrowth cells, derived by plating PMBCs on fibronectin-coated culture plates for 2 days and then replating the nonadherent cell population once more on fibronectin-coated plates. After 4–9 days some colonies emerge, which display a central core of round cells and peripheral spindle-shaped cells. These cells show features similar to CACs. CACs and CFU-Hill cells do not display clonal proliferative status or replating potential; they do display low proliferative potential. Taking clonogenic and proliferative potential into account, endothelial colony-forming cells (ECFCs) or late EPCs, 7 also called endothelial outgrowth cells (EOCs) or blood outgrowth endothelial cells (BOECs), 7,8 can be isolated by plating human peripheral blood or cord blood-derived mononuclear fractions on collagen-coated plates and removing the nonadherent cells. After 2–3 weeks colonies with distinctive cobblestone morphology emerge, which can be removed and expanded. 9 When injected into animal models of ischemia these cells form vessels in vivo. Within these colonies a subpopulation of high proliferative potential-endothelial colony-forming cells (HPP-ECFCs) has been identified, which can be replaced in secondary and tertiary colonies. HPP-ECFCs can achieve up to 100 population doublings (PDs), display high telomerase activity, and are found mostly in cord blood. HPP-ECFCs give rise to low proliferative potential-endothelial colony-forming cells (LPP-ECFCs) able to proliferate for 20–30 PDs. LPP-ECFCs, present particularly in adult peripheral blood, do not form secondary colonies, but generate endothelial cell clusters and then mature differentiated ECs. Unlike the CFU-ECs isolated from peripheral blood, ECFCs/BOECs/EOCs are not contaminated with hematopoietic cell and represent the true originally defined EPCs as they are the only cells that display capacity for postnatal vasculogenesis.
Although flow cytometric analysis has been used to identify various peripheral blood EPC subsets, there are no unique phenotypic markers that are not shared with mature ECs or hematopoietic cells. 10 All the various populations of putative human EPCs express many EC antigens and functional molecules, including CD31, CD34, CD105, vascular endothelial growth factor receptor type 1 (VEGFR1) and VEGFR2 (KDR/Flk1), Tie2, von Willebrand factor (vWF), CXCR4, and aldehyde dehydrogenase (ALDH)bright, and exhibit acLDL uptake. Importantly, ECFCs do not express the hematopoietic markers CD45, CD14, and CD115. 2,11 Moreover, ECFCs do not express CD133, 11 which was once considered a specific EPC marker. At present, it is accepted that early EPCs originate from CD45+ hematopoietic cells whereas late EPCs derive from the CD34+CD45– cell fraction. 12,13 Also, resident ECFCs have been demonstrated 14 and identified within the vascular wall. 15 After injury or during postnatal vasculogenesis, circulating angiogenic cells are recruited to damaged endothelium, where they cooperate to form new vessels, secrete angiogenic molecules, and create a proangiogenic microenvironment. 2 Table 1 summarizes the features of the various EPC subtypes. We refer to endothelial progenitor cells as EPCs in the rest of this review, and clearly highlight the various EPC subtypes used in preclinical and/or clinical therapeutic applications.
Functional and antigenic features of main endothelial progenitor cell subtypes as determined in various assays
Note: Shown here are the phenotypic and functional characteristics of “circulating” EPCs present in the blood of cord and peripheral vessels. EPCs may also be isolated directly from bone marrow (BM). Many investigators have begun to describe such BM-derived EPCs according to the same terminology used for circulating EPCs, namely “colony-forming unit-Hill” (CFU-Hill) cells or “circulating angiogenic cells” (CACs), highlighting their role as hematopoietic myeloid cells involved in promoting new vessel growth (see Refs. 2 and 4).
acLDL, acetylated low-density lipoprotein; ALDH, aldehyde dehydrogenase; BOECs, blood outgrowth endothelial cells; CACs, circulating angiogenic cells; CFU, colony-forming unit; ECFCs, endothelial colony-forming cells; ECs, endothelial cells; EPCs, endothelial progenitor cells; OECs, outgrowth endothelial cells; PACs, proangiogenic hematopoietic cells; UEA-1, Ulex europaeus agglutinin-1; VEGFR, vascular endothelial growth factor receptor; vWF, von Willebrand factor.
Since the discovery of EPCs strong evidence supports their crucial role in angiogenesis and in vasculogenesis, both in vascular diseases and cancer. EPCs are recruited to ischemic tissues, where they are able to form new vessels and, therefore, have been used to treat ischemic diseases with favorable results. 16 Also, the hypoxic tumor microenvironment recruits EPCs, which contribute to tumor growth and progression. In this case, EPCs can be used as vectors of antitumor agents, indicating the possible use of autologous cells, mobilized from the bone marrow of each patient with cancer.
EPCs in Tumor Vascularization and Metastasis
Many studies suggest an important role for EPCs in tumor vascularization, progression, and metastasis. 17 –19 The amount of circulating EPCs positively correlates with tumor progression, and their levels decrease after initiation of chemotherapy. 20 Some authors report a substantial presence of EPCs in tumor endothelium, whereas others suggest lower or undetectable involvement. 21 In any case EPCs, recruited into the tumor mass, contribute to the “angiogenic switch” both directly by their incorporation in cancer vessels or indirectly via paracrine secretion of proangiogenic cytokines. 22
In vitro and in vivo evidence show that mesenchymal stem cells (MSCs) are also involved in tumor growth by providing stromal support for both cancer cells and vasculature. Moreover, tumor cells transplanted subcutaneously have elevated capabilities of proliferation, angiogenesis, and metastasis promotion when they are mixed with MSCs. 23 Thus, EPCs and MSCs serve two different strategies in tumors: EPCs are involved in tumor angiogenesis and MSCs contribute to the maintenance of both tumor stroma and connective tissue. Melero-Martin and Dudley 24 reviewed the important cross-talk between tumor cells, cancer stem cells, and all the types of progenitor cells (EPCs, hematopoietic progenitor cells [HPCs], and mesenchymal progenitor cells [MPCs]), which constitute the tumor microenvironment and work in concert to form vessels.
Under normal conditions, EPCs reside in the BM within a stem cell niche, where they interact with bone marrow stromal cells (BMSCs). EPCs are retained in the niche by high levels of the chemokine SDF1 (stromal-derived factor-1), some integrins such as α4β1 and β3, and low levels of c-Kit ligand. In response to tumor growth, the niche microenvironment is perturbed and EPCs can leave the BM, enter the peripheral blood, and migrate into the tumor to form new vessels. All the steps of this process are summarized in comprehensive reviews by Laurenzana and colleagues 18 and De la Puente and colleagues. 25 Hypoxia, present at the site of tumor mass, induces the production and release from the BM of many factors involved in EPC mobilization (VEGF, the most important regulator of EPC mobilization, granulocyte colony-stimulating factor [G-CSF], basic fibroblast growth factor [bFGF], SDF1), which reach the stem cell niche in the BM, where they activate BMSCs and induce matrix metalloproteinase (MMP)-9 upregulation. In addition, neutrophil elastase and cathepsin G impair the interaction between α4β1 and β3 integrins and stromal cells, thus causing detachment of EPCs from the BM. Proteases play an important role in EPC mobilization and intravasation into the peripheral blood, as exhaustively reviewed. 18 The migration of circulating EPCs toward the tumor mass is regulated by chemotactic tumor-secreted cytokines that interact with their cognate receptor on EPC membrane. The subsequent EPC extravasation and tumor homing depend on the interaction of EPC integrins with the adhesion molecules expressed by ECs of the vessel wall. Afterward, EPCs invade the extracellular matrix (ECM) to reach the tumor site, where they contribute to form new vessels and thus favor cancer progression. This process requires the activity of proteases able to degrade the ECM and to enable EPCs to invade host tissue. 18 The last phase of tumor angiogenesis is EPC differentiation, which takes place in three steps: adhesion to ECM mediated by integrins, secretion and activation of proangiogenic factors, and overexpression of transcription factors (histone deacetylases [HDACs] and HoxA), which regulate the expression of typical EC genes such as those encoding endothelial nitric oxide synthase (eNOS), VEGFR-2, and vascular endothelial (VE)-cadherin. 26
Because of their ability to home within tumors, EPCs can be used in anticancer cell therapy as cellular vehicles for delivering molecules able to impair cancer progression, as well as imaging probes when properly labeled, 27 thus providing a so-called “theranostic” approach. In addition to their use in cancer cell therapy, EPCs can be a therapy target, by reducing their mobilization and/or paracrine angiogenic activity, a topic that is not evaluated in this review. Unfortunately, the number of EPCs that can be isolated from various sources is low (fewer than 1 per 10,000 circulating mononuclear cells and less than 1% of bone marrow cells 1 ). A solution to overcome this obstacle is the possibility to increase cell mobilization in vivo after injection of some chemotactic or growth factors, 18 to expand isolated EPCs in vitro, and to transfect telomerase reverse transcriptase (TRT), allowing proliferation without experiencing senescence and thereby increasing functional competence. 28
EPCs in Cell-Based Therapy
Cell-based therapies are becoming more and more relevant. Taking into account the tumor-homing properties of EPCs, two different approaches to control cancer progression can be evaluated by combining cell-based therapy with gene therapy or with nanomedicine. The first approach is based on the possibility to engineer EPCs to express various transgenes, and the second approach is based on the capacity of EPCs to take up nanomaterials. The EPC subtype that is currently used in almost all experimental therapeutic strategies of tumor targeting is represented by BOECs (ECFCs), which are assumed to appear from the bone marrow but also from the endothelial lining of blood vessels, 29 and that can be prepared from the blood of umbilical cord and peripheral vessels. Their tumor-homing properties have been repeatedly assessed, including a specific study using 111In-labeled BOECs from human umbilical cord injected into mice with a C3H mammary carcinoma foot tumor. 30 In that study 111In was detected not only in tumor but also in other, healthy organs, even if off-target activity was ascribed mainly to 111In released from BOECs. 111In activity was revealed in the tumor rim and microscopy demonstrated that such activity originated from human BOECs not located in the vessel wall.
EPCs and Gene Therapy
From the 1980s to 1990s many studies have been performed to exploit gene therapy by using viral vectors carrying therapeutic genes 31 (Fig. 1A). One of the major challenges in the use of viral vectors in gene therapy is their distribution in healthy organs and tissues, thus causing off-target toxicity. Thus, gene therapy combined with cell therapy has been developed to deliver therapeutic genes to the pathological sites. 31,32 This approach consists of the transfection of human stem cells with viral vectors, such as lentivirus, that express the relevant therapeutic gene. Expanded engineered cells can be injected into patients. EPCs have been studied as a potential anticancer delivery system. 33 The ability of EPCs to reach high transduction efficiency and their use in cell therapy have been investigated, resulting in the demonstration that late EPCs display higher lentiviral vector transduction efficiency than early EPCs. Particular attention to the potential modification of EPCs after viral transduction must be paid before clinical use. 34 Moreover, the transduction efficiency of EPCs strictly depends on the choice of the appropriate gene promoter. 35 To control in vivo gene expression the Tet-on/off system has been used. 36
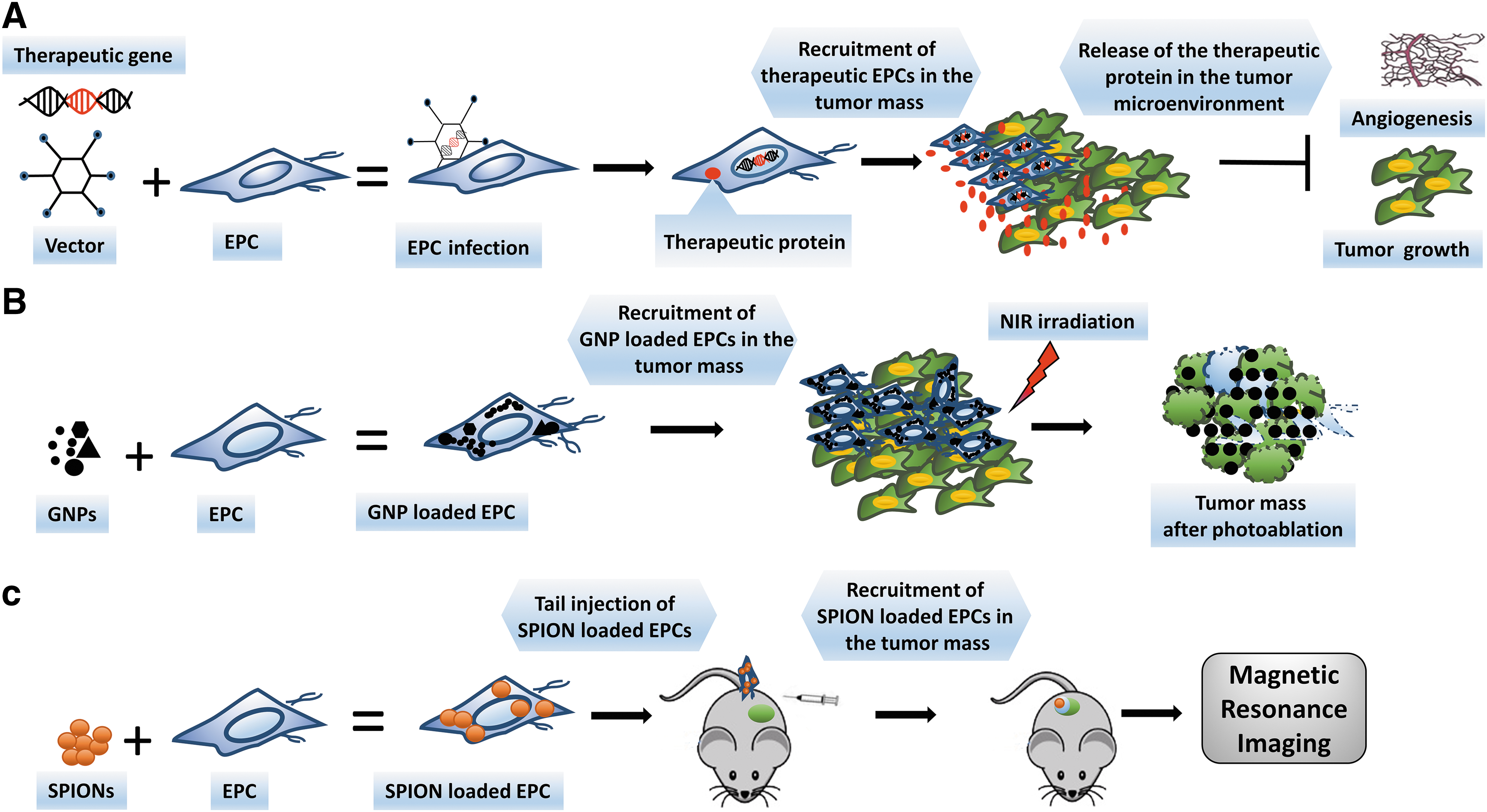
Endothelial progenitor cells (EPCs): a multifunctional tool for cancer therapy and imaging.
EPCs and antiangiogenic/antitumor gene therapy
An effective anticancer approach relies on the inhibition of angiogenesis and tumor growth. It has been demonstrated that the soluble, truncated VEGFR-2 (sFlk1) acts as a decoy, by binding with high affinity to VEGF and preventing it from interacting with the cellular, fully functional, VEGF receptor. Actually, gene-modified EPCs, expressing sFlk1, inhibit angiogenesis and tumor growth in mice. 37 The same antitumor effect has been obtained in Lewis lung carcinoma by engineering EPCs with a retroviral vector expressing the angiogenetic inhibitor endostatin. 38 We have engineered BOECs (ECFCs) with lentivirus encoding the uPAR (urokinase plasminogen activator receptor)-degrading MMP-12. As the native full-length uPAR plays a central role in the invasive step of angiogenesis and spreading of cancer cells, delivery of MMP-12 into the tumor mass cleaves uPAR on tumor cells and on ECs and strongly inhibits the angiogenesis, growth, and development of lung metastases in nude mice xenografted with human melanoma cells. 39
EPCs and suicide gene therapy
The suicide gene therapy approach consists of arming EPCs with viral vectors expressing genes that produce enzymes able to metabolize prodrugs into anticancer drugs that kill tumor cells by a “bystander effect.” EPCs that express the prodrug-metabolizing enzyme will be killed in the presence of the prodrug that, on activation, will also kill neighboring nontransduced bystander tumor cells. BOECs (ECFCs) are usually exploited for such therapy, and the most extensively studied suicide genes are those encoding thymidine kinase (TK), cytosine deaminase (CD), and nitroreductase, which transform initially harmless drugs (ganciclovir, 5-fluorocytosine [5-FC], and dinitroaziridinyl benzamide, respectively) into highly toxic ones (reviewed by Dudek et al 40 ). Several limitations are, however, highlighted in the suicide approach. 40 First, the effect of suicide therapy is not optimal when the suicide cell-to-tumor cell ratio is lower than 1:10. Second, the ability of BOECs transfected with the cytosine deaminase gene to home to tumor sites is modest, resulting in failure to improve the survival of mice displaying multiple organ metastases. Wei and coworkers 41 have shown that BOECs home to lung metastases of Lewis lung carcinoma (LLC), although to different extents for single metastases, while liver and kidney metastases underwent BOEC homing to a low extent. When expressing a suicide gene, BOECs exerted a bystander effect on LLC cells even if they did not increase animal survival. Homing efficacy was not the only important parameter to judge BOEC efficacy. In fact, their susceptibility to suicide gene-induced cell death, their growth-promoting activity on LLC cells, and their low proliferation rate compared with that of LLC cells must also be taken into consideration.
EPCs and immune-stimulating gene therapy
EPCs have also been used in immunotherapy. In two studies bone marrow-derived EPCs were transfected with a viral vector encoding interleukin (IL)-12, 42 and mouse microvascular endothelial cells (MECs) were isolated from the lungs of BALB/c and C57BL/6 mice and transfected with IL-2. 43 On delivery into the tumor mass, activation of cytotoxic lymphocytes and natural killer cells was observed, inducing a decrease in tumor burden and prolonging animal survival. In cancer immune therapy, modification of the immune-suppressive cancer microenvironment is of paramount importance. The injection of EPCs transduced with the immune-stimulatory gene CCL19 exerted an antitumor effect in a mouse model of ovarian cancer, in terms of lung metastasis and peritoneal dissemination. 44 The CD40–CD40 ligand (CD40L) system is of interest in antitumor immune therapy. CD40 is located on antigen-presenting cells (APCs) and, on interaction with CD40L on helper T cells, stimulates APC activation. However, many normal and tumor cells also express CD40 (reviewed in Ref. 45). In particular, cancer cells use the CD40–CD40L pathway (which is activated to a low level) to drive tumor proliferation and survival. When intense activation of CD40 occurs, it produces growth inhibition and apoptosis of cancer cells. Apoptosis depends on increased expression of Bcl2-associated X protein and of membrane-associated cytotoxic ligands of the tumor necrosis factor (TNF) family, such as FasL, TNF, and TNF-related apoptosis-inducing ligand (TRAIL). 45 CD40L alone or combined with chemotherapy inhibits the development of xenografted human breast tumors and prolongs animal survival. 46 Therefore, CD40 activation can exert a direct antitumor effect. On these bases, EPCs derived from human induced pluripotent stem cells (iPS-EPCs) have been transduced with a baculovirus expressing the CD40 ligand. Intravenous injection of such modified EPCs prolonged the survival of breast cancer-bearing mice by stimulating the secretion of TNF-α and interferon (IFN)-γ and by increasing caspase-3/7 activity in lung metastases. 47 As suggested by the authors, such effects may depend on a iPS-EPCs local activation produced by CD40L–CD40 interactions in the tumor through a bystander mechanism, involving the intercellular transfer of CD40L from iPS-EPCs to tumor cells.
EPCs and oncolytic viral therapy
EPCs combined with oncolytic viral therapy has been hypothesized in place of classic oncolytic viral therapy, which presents many problems concerning therapeutic levels of oncolytic viruses carried to metastases. 48 Wei and colleagues 49 demonstrated that, after intravenous and peritumoral injection into irradiated orthotopic U87 gliomas in mice, BOECs transfected with attenuated measles virus of the Edmonston B strain (MV-Edm) produce a decrease in tumor size.
EPCs and tumor microenvironment targeting
Modified EPCs have also been employed to target the cross-talk between EPCs and cells of the tumor microenvironment, such as tumor-associated macrophages (TAMs). Transfection of a soluble form of CD115 (the macrophage colony-stimulating factor receptor) via EPCs produced a decrease in TAMs in xenografts, as well as a reduction of tumor size of human prostatic cancer cells after tumor localization of EPCs. 50 In this study authors used circulating endothelial precursors (CEPs), cells that closely resemble circulating endothelial progenitors (EPCs/CEPs), endothelial colony-forming cells (ECFCs), and blood outgrowth endothelial cells (BOECs).
EPCs and Nanomedicine
Nanomaterial-mediated tumor therapy has significantly advanced and is becoming a pillar of modern medicine. In particular, gold nanoparticles (GNPs) are promising therapeutic tools for the treatment of cancer because of the following features: (1) their noncytotoxic nature, (2) reliable synthesis and functionalization, and (3) tunable plasmonic properties. This allows the production of gold nanostructures able to generate local heating on exposure to near-infrared (NIR) light. 51 Although multifunctional nanoparticles (NPs) have been used as photothermal agents or as vehicles of chemotherapeutics in cancer treatment, at the current stage of development the majority of administered NPs end up also in healthy organs and tissues. This happens even with the assistance of active targeting with tumor-tropic molecules, a feature that does not eliminate clearance by macrophages, inaccessibility to the hypoxic areas of tumors that lack blood flow, and off-target distribution, thus limiting the treatment efficacy. 51 In light of these considerations, a possible way to improve delivery of NPs into tumors could take advantage of the natural tumor-homing activity of EPCs (Fig. 1B). In our laboratory we have optimized ECFC uptake of GNPs. 52 Both in vitro and in vivo assays have shown the excellent thermotransductive properties of GNP-enriched ECFCs and their ability to kill melanoma cancer cells at moderate NIR light intensities, 52 providing a rationale for efficient tumor ablation by a mixed approach (cell therapy and nanomedicine).
EPCs and Molecular Imaging
EPCs have also been proposed for diagnostic purposes. The possibility to label tumor-tropic cells may allow monitoring to localize primary and metastatic tumors and a parallel follow-up of the same lesions after therapy. EPCs, labeled with superparamagnetic iron oxide nanoparticles (SPIONs) or gadolinium, have been successfully used in magnetic resonance imaging of human breast cancer animal models 53 and CD133+ cells, freshly isolated from cord blood, in a rat model of human glioma 54 (Fig. 1C), as well as in in vivo studies of tumor angionenesis. 53,55 After 99Tc or 111In uptake and single-photon emission tomography (SPECT), EPCs have been shown to localize in xenografted human glioma 54 and human melanoma. 39 In the near future, the possibility of using ECFCs or other tumor-tropic stem cells carrying a nanoparticle payload with both therapeutic and diagnostic properties will allow a “theranostic” approach for personalized tumor medicine.
Clinical Translation and Conclusions
A list of the cancer clinical trials initiated, completed, or withdrawn is available at
Footnotes
Acknowledgments
Work by the authors cited in this review (Refs. 39 and ![]() ) were financially supported by Associazione Italiana Ricerca sul Cancro (AIRC) grant IG 2013 N. 14266 (M.D.R.), by Ente Cassa di Risparmio di Firenze and by Istituto Toscano Tumori (L.C.). A.C. was supported by a postdoctoral fellowship of the Federazione Italiana Ricerca sul Cancro (FIRC). The authors thank Dr. Benedetta Mazzanti for fruitful discussion and suggestions.
) were financially supported by Associazione Italiana Ricerca sul Cancro (AIRC) grant IG 2013 N. 14266 (M.D.R.), by Ente Cassa di Risparmio di Firenze and by Istituto Toscano Tumori (L.C.). A.C. was supported by a postdoctoral fellowship of the Federazione Italiana Ricerca sul Cancro (FIRC). The authors thank Dr. Benedetta Mazzanti for fruitful discussion and suggestions.
Author Disclosure
No competing financial interests exist.