
Abstract
Human immunodeficiency virus type 1 (HIV-1)-based lentiviral vectors efficiently transduce genes to human, but not rhesus, primary T cells and hematopoietic stem cells (HSCs). The poor transduction of HIV-1 vectors to rhesus cells is mainly due to species-specific restriction factors such as rhesus TRIM5α. Previously, several strategies to modify HIV-1 vectors were developed to overcome rhesus TRIM5α restriction. While the modified HIV-1 vectors efficiently transduce rhesus HSCs, they remain suboptimal for rhesus primary T cells. Recently, HIV-1 variants that encode combinations of LNEIE mutations in capsid (CA) protein and SIVmac239 Vif were found to replicate efficiently in rhesus primary T cells. Thus, the present study tested whether HIV-1 vectors packaged by a packaging construct containing these CA substitutions could efficiently transduce both human and rhesus primary CD4 T cells. To accomplish this, LNEIE mutations were made in the packaging construct CEMΔ8.9, and recombinant HIV-1 vectors packaged by Δ8.9 WT or Δ8.9 LNEIE were generated. Transduction rates, CA stability, and vector integration in CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/green fluorescent protein cells, as well as transduction rates in human and rhesus primary CD4 T cells by Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors, were compared. Finally, the influence of rhesus TRIM5α variations in transduction rates to primary CD4 T cells from a cohort of 37 Chinese rhesus macaques was studied. While it maintains efficient transduction for human T-cell line and primary CD4 T cells, Δ8.9 LNEIE-packaged HIV-1 vector overcomes rhesus TRIM5α-mediated CA degradation, resulting in significantly higher transduction efficiency of rhesus primary CD4 T cells than Δ8.9 WT-packaged HIV-1 vector. Rhesus TRIM5α variations strongly influence transduction efficiency of rhesus primary CD4 T cells by both Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors. Thus, it is concluded that Δ8.9 LNEIE-packaged HIV-1 vector overcomes rhesus TRIM5α restriction and efficiently transduces both human and rhesus primary T cells.
Introduction
S
Human immunodeficiency virus type 1 (HIV-1)-based lentiviral vectors are ideal for delivering potentially therapeutic genes into human primary T cells and HSCs. Unlike Mo-MuLV-based vectors, HIV-1-based vectors can transduce both dividing and non-dividing cells, and display a tendency to integrate into actively transcribed genes. 26,27 Moreover, the new generation of self-inactivation (SIN) HIV-1-based vectors have been modified in such a way that prevents activation of genes surrounding integration sites. 28 Because of these alterations, HIV-1-based vectors are being used in a number of clinical trials for immune deficiencies, hemoglobin disorders, metabolic diseases, and HIV-1/AIDS. 29
While HIV-1-based vectors can efficiently transduce genes into human primary T cells and HSCs, they poorly transfer genes into rhesus primary T cells and HSCs. 30,31 Because rhesus macaques are an extensively utilized preclinical model for evaluating the safety and the efficacy of gene therapy approaches for human diseases, it is useful and important to develop HIV-1-based vectors that can efficiently transduce both human and rhesus macaque primary T cells and HSCs.
The poor efficiency of transduction of genes into rhesus primary T cells and HSCs by HIV-1-based vectors is due to species-specific restriction factors in rhesus macaques that inhibit HIV-1 infection. 32 Key post-entry restriction factors are the rhesus TRIM5α and TRIM5-CypA fusion proteins. 33 –35 As a member of a large family of tripartite motif (TRIM) containing proteins, TRIM5α and TRIM5-CypA contain N-terminal RING, B-box, coiled-coil, and C-terminal SPRY/B30.2 or CypA domain, respectively. Rhesus TRIM5α and TRIM5-CypA recognize the incoming HIV-1 core by binding its capsid (CA) protein. Subsequently, TRIM5α or TRIM5-CypA are poly-ubiquinated, and ubiquinated TRIM5α or TRIM5-CypA along with the HIV-1 core complex are degraded. 36,37 Simian immunodeficiency virus (SIV) CA can escape rhesus TRIM5α and TRIM5α-CypA-mediated degradation. Several strategies to modulate these factors have been developed to improve transduction to macaque cells by modified HIV-1 vectors. 38 –40 For example, Kootstra et al. 38 and Rits et al. 39 constructed modified HIV-1 vectors with mutations in the cyclophilin A (CypA)-binding domain of the CA sequence in the gag/pol packaging constructs. They demonstrated that the modified HIV-1 vectors could efficiently transduce simian cell lines CV-1 and FrHL2 cells and baboon CD34+ HSCs, however, transduction rates in rhesus peripheral blood mononuclear cells (PBMCs) were <10%. Uchida et al. 40 screened a panel of chimeric HIV-1 vectors containing various combinations of HIV-1 and SIV elements in their transduction efficiency in human and rhesus lymphocytic cell lines CEMx174 and LCL8664, and selected a chimeric HIV-1 (χHIV) vector, in which the vector genome was packaged with SIV CA. They demonstrated that the χHIV vector efficiently transduces both human and rhesus CD34+ HSCs ex vivo. In vivo competitive repopulation experiments showed that this chimeric vector had better marking levels than either an unmodified HIV-1 vector 41 or SIV-based vector. 42 However, in the rhesus lymphoblast cell line LCL8664, transduction rates with this chimeric vector were <25%, even though in CEMx174 transduction rates were >90%, 40 and no transduction efficiency in rhesus primary T cells has been reported with this χHIV vector. Besides, several groups developed SIV-based vectors and demonstrated that SIV-based vectors can efficiently transduce rhesus CD34+ HSCs, 43 –45 and non-myeloablative conditioning regimen increases engraftment of gene-modified HSCs in young rhesus macaques. 45 However, no transduction to rhesus primary T cells by SIV-based vectors has been reported in these studies. Thus, it appears that although these modified HIV-1 vectors or SIV-based vectors can efficiently transduce rhesus CD34+ HSCs, transduction rates of rhesus primary T cells by these vectors remain suboptimal.
Recently, in the development of nonhuman primate models for HIV-1 infection, Soll et al. 46 employed an “assisted evolution” approach in which individual CA mutations that reduced rhesus TRIM5α resistance without fitness penalties were randomly assorted in a library of viral clones containing synthetic CA sequences. They identified viruses that encoded combinations of five mutations in CA, namely LNEIE, that conferred complete resistance to disruptive effects of rhesus TRIM5α on incoming HIV-1 core. As a result, HIV-1 variants encoding these CA substitutions and SIVmac239 Vif replicated to high levels in rhesus primary lymphocytes. It was hypothesized that HIV-1 vectors packaged by a HIV-1 Gag/Pol expression construct with these CA substitutions could improve transduction of both human and rhesus primary T cells and HSCs.
The present study focused on testing whether HIV-1 vectors packaged by a LNEIE CA modified Gag/Pol expression construct would overcome restriction by TRIM5α and TRIM-CypA and efficiently transduce both human and rhesus primary CD4 T cells. To carry out these studies, LNEIE mutations were introduced in the CA region of the gag/pol expression construct pCEMΔ8.9, 26 and vesicular stomatitis virus glycoprotein G (VSV-G) pseudotyped HIV-1 vectors packaged using either Δ8.9 WT (wild-type pCEMΔ8.9) or Δ8.9 LNEIE (pCEMΔ8.9 containing LNEIE mutations) gag/pol expression constructs were generated. The study demonstrates that while the Δ8.9 LNEIE-packaged HIV-1 vector enables efficient transduction of human T-cell lines and primary CD4 T cells, it overcomes rhesus TRIM5α-mediated CA degradation, resulting in significantly higher transduction efficiency of rhesus primary CD4 T cells than Δ8.9 WT-packaged HIV-1 vector. It also demonstrates that variation of rhesus TRIM5α strongly influences transduction efficiency of rhesus CD4 T cells by both Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors.
Materials and Methods
Cell lines
The packaging cell line 293FT was purchased from Invitrogen Life Technologies (Carlsbad, CA) and maintained in complete Dulbecco's modified Eagle's medium (DMEM; i.e., high glucose DMEM supplemented with 10% fetal bovine serum [FBS], 2 mM of L-glutamine, 1 mM of sodium pyruvate, 100 IU/mL of penicillin, and 100 μg/mL of streptomycin). CEMss-CCR5 cells were generated previously, 47 and CEMss-CCR5-rhTRIM5a/green fluorescent protein (GFP) cells were generated in this study (see below) and maintained in complete DMEM.
Human and rhesus macaque primary CD4 T cells
Human PBMCs were obtained from healthy anonymous donors through the blood bank of the Changhai Hospital, Shanghai, China. Human primary CD4 T cells were enriched from PBMCs by negative selecting magnetic beads according to the manufacturer's instructions (Cat# 11346D; Thermo Fisher Scientific, Waltham, MA). Rhesus PBMCs were provided by the Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences Beijing, China (Animal Experimental Approval# XJ16006). Rhesus primary CD4 T cells were isolated from PBMCs by positive selecting magnetic beads according to the manufacturer's instructions (Cat# 130-091-102, Miltenyi Biotec, Bergisch Gladbach, Germany). Both human and rhesus primary CD4 T cells were re-suspended in the complete RPMI 1640 medium (i.e., RPMI 1640 medium supplemented with 15% FBS, 2 mM of L-glutamine, 1 mM of sodium pyruvate, 100 IU/mL of penicillin, and 100 μg/mL of streptomycin) supplemented with human rIL-2 (100 IU/mL; R&D Systems, Minneapolis, MN) before being activated and transduced with Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors (see below).
Gene constructs
Δ8.9 LNEIE gag/pol packaging construct was directly synthesized by Genewiz, a service company in Suzhou, China, in which five mutations (M10L/I91N/A92E/M96I/G116E) in the capsid protein were introduced in the pCMVΔ8.9 gag/pol packaging construct. 48 The resulting packaging constructs were designated as Δ8.9 WT and Δ8.9 LNEIE, respectively (Fig. 1A).
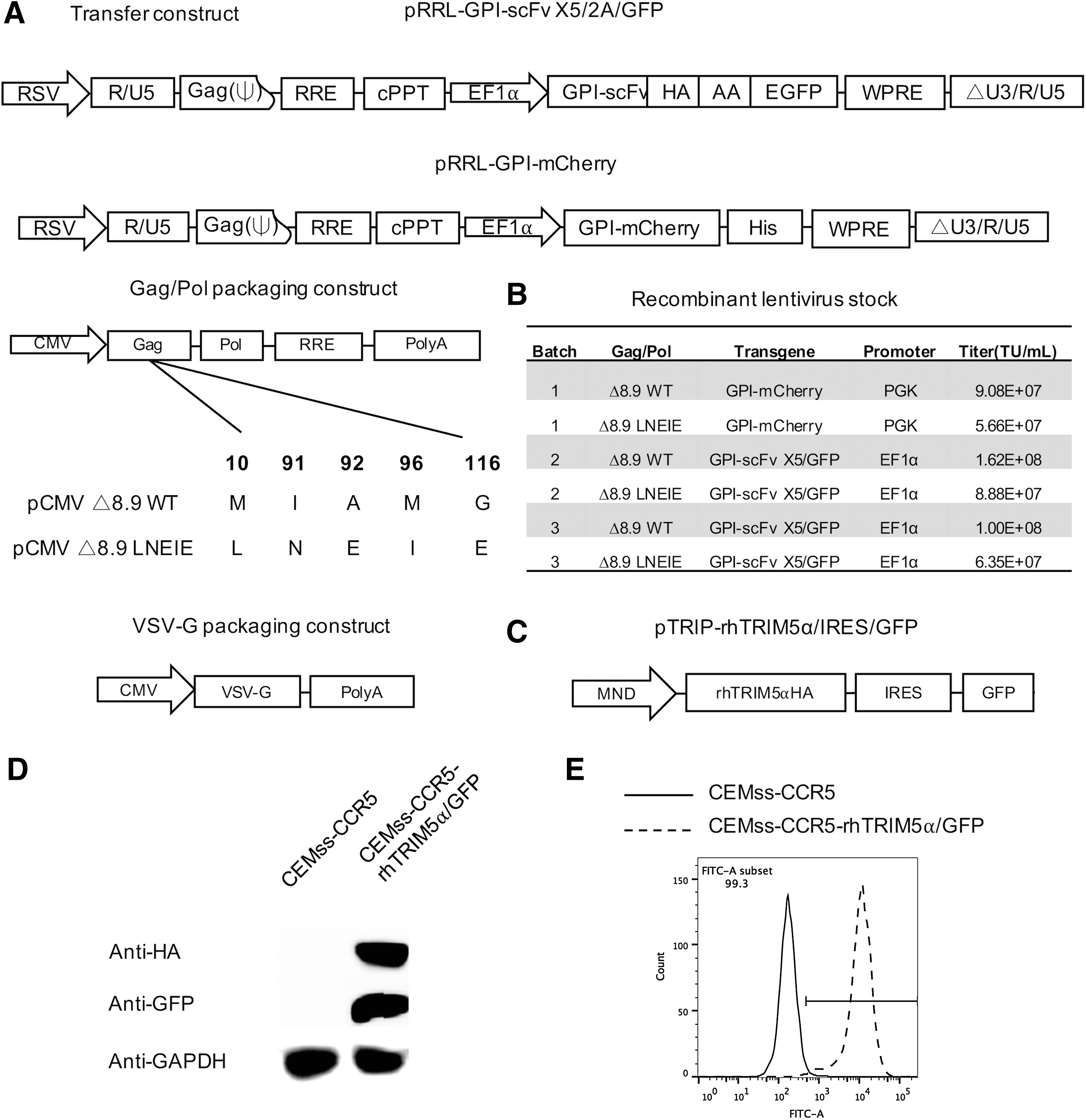
Lentiviral transfer and packaging constructs and transduction of CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/green fluorescent protein (GFP) cells with Δ8.9 wild type (WT) or Δ8.9 LNEIE-packaged human immunodeficiency virus type 1 (HIV-1) vectors. (
The fusion gene encoding the glycosyl-phosphatidylinositol (GPI)-scFv X5 was generated by polymerase chain reaction (PCR) using pRRL-GPI-scFv X547 as a template and ligated between the Bam HI and Sal I sites of a third-generation lentiviral transfer vector, pRRLsin-18.PPT.EF1α.2A.GFP.Wpre. 49 The resulting lentiviral transfer constructs were designated pRRL-GPI-scFv X5/2A/GFP (Fig. 1A).
Fusion gene encoding the GPI-mCherry/his tag was generated by overlapping PCR and ligated between the Bam HI and Sal I sites of a third-generation lentiviral transfer vector, pRRLsin-18.PPT.PGK.Wpre. 50 The resulting lentiviral transfer constructs were designated pRRL-GPI-mCherry (Fig. 1A).
The gene encoding rhesus TRIM5α was amplified by PCR using pLPCX-rhTRIM5α vector
51
as a template and a pair of primers (5′-GG
Generation of recombinant lentiviruses
Recombinant lentiviruses were generated as described previously. 53 Briefly, 4 × 106 293FT cells were seeded onto a P-100 dish in 10 mL of complete DMEM. After culturing overnight, cells were co-transfected with transfer construct (20 μg) (one of the pTRIP-rhTRIM5α/IRES/GFP, pRRL-GPI-mCherry, or pRRL-GPI-scFv X5/2A/GFP constructs), gag/pol packaging construct Δ8.9 WT (7 μg), or Δ8.9 LNEIE (20 μg; the different amounts of Δ8.9 WT or Δ8.9 LNEIE used here were based on the results of our initial assay optimization, in which various amounts of Δ8.9 WT or Δ8.9 LNEIE had been tested) and plasmids encoding the VSV-G protein envelope (pLP/VSV-G; 3 μg) using a calcium phosphate precipitation method. Sixteen hours later, culture supernatants were removed and replaced with fresh complete DMEM plus 1 mM of sodium butyrate (Cat# B58875G; Sigma–Aldrich, St. Louis, MO). Eight hours later, supernatants were again removed and replaced with fresh DMEM plus 4% FBS. After another 20 h, the culture supernatants were harvested and concentrated by ultracentrifugation as described previously. 47 The vector pellets were re-suspended in a small volume of DMEM and stored in aliquots in a –80°C freezer. Vector titers before and after the concentration were determined as previously described. 47
Generation of CEMss-CCR5-rhTRIM5α/GFP cell line
To generate a stably transduced CEMss-CCR5-rhTRIM5α/GFP cell line, 1 × 105 CEMss-CCR5 cells and 2 × 106 transducing units (TU) of the recombinant lentiviruses containing pTRIP-rhTRIM5α/IRES/GFP were added to a 24-well tissue culture plate in the presence of 8 μg/mL of polybrene (Cat# H9628; Sigma–Aldrich). Twenty-four hours later, cells were extensively washed and cultured in complete DMEM. The expression of fusion gene rhTRIM5α/IRES/GFP was determined by Western blot and by fluorescence-activated cell sorting (FACS) analysis (see below).
Comparison of transduction efficiency between Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vector in CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/GFP cell lines
To compare transduction efficiency between Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vector, CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/GFP cells (105 cells per well) were transduced with Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-mCherry at a multiple of infection (MOI) of 0.625, 1.25, 2.5, 5, and 10 in 500 μL of complete DMEM with 8 μg/mL of polybrene. The cells were centrifuged at 1,500 g for 3 h at 37°C to permit virus attachment. After overnight culture, cells were washed twice and re-suspended in fresh complete DMEM. Seven days later, the GPI-mCherry and/or GFP expression was evaluated by flow cytometry.
Western blot
To measure stability of CA proteins, 106 CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/GFP cells were transduced with Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-mCherry at a MOI of 10 in 500 μL of complete DMEM with 8 μg/mL of polybrene and centrifuged at 1,500 g for 1 h at 4°C to permit virus attachment and to synchronize the transduction. After washing cells with DMEM twice to remove unattached virus, cells were incubated at 37°C. At 0, 1, 2, and 4 h post transduction, cells were harvested and lysed using the lysis buffer (100 mM Tris-HCl, pH 8.0, and 1% NP40) containing protease inhibitor cocktail (Cat# 539134; Calbiochem, Billerica, MA). The lysis samples were incubated for 15 min at 90°C in loading buffer containing SDS and 0.6 M of dithiothreitol and loaded onto 12% SDS-PAGE. Gels were transferred onto polyvinylidene difluoride membranes, and the membranes were blocked with Tris-buffered saline containing 0.1% Tween-20 (TBST) and 5% nonfat dry milk for 1 h at room temperature. The membranes were subsequently incubated with 3 mL (0.5 μg/mL) of mouse anti-Gag p24 antibody (1:1,000) and rabbit anti-hemagglutinin (HA) antibody (1:1,000) overnight at 4°C. After being washed twice with TBST, the membranes were probed with HRP-conjugated goat anti-mouse antibody (1:5,000) and HRP-conjugated goat anti-rabbit antibody (1:4,000), respectively. Mouse anti-Gag p24 antibody (183-H12-5C) was obtained from the NIH ARRRP. Goat anti-HA antibody was purchased from Sigma–Aldrich (Cat# H6980). HRP-conjugated goat anti-mouse antibody and HRP-conjugated goat anti-rabbit antibody were purchased from ICL.
Quantitative PCR
Before the transduction, the recombinant lentiviruses were treated by DNase I (Cat# D7073; Beyotime, Haimen, China) according to the manufacturer's instructions. To synchronize the infection, CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/GFP cells were exposed to Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors at a MOI of 5, centrifuged at 1,500 g for 1 h at 4°C to permit virus attachment, and washed twice with DMEM to remove unattached virus. Cells were then incubated at 37°C. At 0, 4, 8, and 16 h post transduction, 1 × 105 cells per sample were harvested and frozen in −20°C. Genomic DNA was extracted by the MiniBEST Universal Genomic DNA Extraction Kit (Cat# 9765; TaKaRa, Otsu, Japan). Equal amounts of genomic DNA were used in triplicate and amplified with the THUNDERBIRDTM SYBR qPCR Mix provided by TOYOBO (Osaka, Japan). The level of expression of integration ratio was determined using the ABI 7900HT. Amplification efficiencies were validated and normalized against host-keeping gene GAPDH, and fold increases were calculated using the comparative CT method for quantification. The primers for transfer constructs were: WPRE F (5′-GGCACTGACAATTCCGTGGT-3′), WPRE R (5′-AGGGACGTAGCAGAAGGACG-3′); GAPDH: GAPDH-gF (5′-CTAGGGCTGCTCACATATTCTGG-3′), GAPDH-gR (5′-TAGTTGAGGTCAATGAAGGGGTC-3′).
Transduction of human and rhesus primary CD4 T cells as well as rhesus PBMCs
Human CD4 T cells (2.5 × 105 per well) were activated with anti-CD3/CD28 antibody-coated microbeads (Cat# 11131D; Thermo Fisher Scientific) at a 1:1 ratio in 500 μL of complete RPMI 1640 medium supplemented with human rIL-2 (100 IU/mL; Cat# 202-IL; R&D Systems) in 48-well plates. After 24 h, 5 × 106 TU Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-scFv x5/2A/GFP in complete RPMI 1640 supplemented with human rIL-2 (100 IU/mL) and 8 μg/mL polybrene were added into the cell suspension at a final volume of 750 μL and various indicated MOIs. The plates were centrifuged at 1,500 g for 3 h at 37°C to facilitate transduction. The anti-CD3/CD28 antibody-coated microbeads in human CD4 T cells were removed by DynaMag magnet after 4 days of activation. The transduced human primary CD4 T cells were then cultured in complete RPMI 6140 supplemented with human rIL-2. The transduction efficiency was measured by FACS analysis.
To transduce rhesus primary CD4 T cells or PBMCs, anti-CD3 antibody (clone# FN-18; Thermo Fisher Scientific) and anti-CD28 antibody (clone# CD28.2; eBioscience, San Diego, CA) were pre-coated at a 1:1 ratio in 500 μL of phosphate-buffered saline (PBS) in 24-well non-tissue culture plates (Cat# 3738; Corning, Corning, NY) at 4°C overnight. The coated plates were then blocked with 1% bovine serum albumin (BSA) in PBS for 30 min at 37°C, washed twice with PBS, and used for stimulation. Rhesus CD4 T cells (5 × 105) or PBMCs were added onto the anti-CD3/CD28-coated plate in 1 mL of complete RPMI 1640 medium supplemented with human rIL-2 (100 IU/mL). After 24 h, 10 × 106 TU Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-scFv x5/2A/GFP in complete RPMI 1640 supplemented with human rIL-2 (100 IU/mL) and 8 μg/mL of polybrene were added into the cell suspension at a final volume of 2 mL and various indicated MOIs. The plates were centrifuged at 1,500 g for 3 h at 37°C to facilitate transduction, and were cultured for an additional 3 days. The transduced rhesus primary CD4 T cells or PBMCs were then cultured in complete RPMI 6140 supplemented with human rIL-2. The transduction efficiency was measured by FACS analysis.
FACS analysis
To study the transgene expression, CEMss-CCR5 or CEMss-CCR5-rhTRIM5α/GFP cells transduced with Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-mCherry or human and rhesus primary CD4 T cells transduced with Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-scFv X5/2A/GFP (2 × 105 per sample) were harvested, washed twice with FACS buffer (PBS containing 1% BSA), and fixed with 1% formaldehyde in 0.5 mL of FACS buffer. FACS analysis was performed on Fortessa (Becton Dickinson, Mountain View, CA).
To study the transgene expression in transduced rhesus PBMCs, rhesus PBMCs transduced with Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-scFv X5/2A/GFP (2 × 105 per sample) were harvested, washed twice with FACS buffer, and stained with APC-conjugated anti-CD4 and PE-conjugated anti-CD8 antibodies (BD Biosciences, San Jose, CA). After the staining, cells were washed twice with FACS buffer and fixed with 1% formaldehyde in 0.5 mL of FACS buffer. FACS analysis was performed on Fortessa.
Rhesus TRIM5α genotyping
Rhesus TRIM5a genotyping was performed using genomic DNA. Genomic DNAs from 37 rhesus macaques were extracted from PBMCs using the MiniBEST Universal Genomic DNA Extraction Kit (Cat# 9765; TaKaRa). A 525 bp fragment was generated by PCR using a pair of primers SPRY-006 (5′-CAGTGCTGACTCCTTTGCTTG-3′) and T5α1087R (5′-GCTTCCCTGATGTGATAC-3′), as previously reported by de Groot et al. 54 The PCR products were then purified and sequenced directly.
Statistics
Statistical analyses were performed using the PRISM v6 software program. Transduction rates and integration ratio were compared using Student's t-test. Correlations were evaluated by R 2 in linear regression and p-values in correlation coefficient. A p-value of <0.05 was deemed significant. Standard errors of the mean are shown as error bars in all figures.
Results
Δ8.9 LNEIE-packaged HIV-1 vector efficiently transduces human T-cell lines and overcomes rhesus TRIM5α restriction
To evaluate whether Δ8.9 LNEIE-packaged HIV-1 vector could overcome restriction by rhesus TRIM5α, first Δ8.9 LNEIE packaging construct was constructed, in which M10L/I91N/A92E/M96I/G116E mutations were introduced into the CA sequence. The Δ8.9 WT or Δ8.9 LNEIE and VSV-G packaging constructs along with one of two SIN transfer vectors pRRL-GPI-mCherry or pRRL-GPI-scFv X5/2A/GFP (Fig. 1A) were co-transfected into the packaging cell line 293FT. Several batches of recombinant HIV-1 vectors were generated, and they were concentrated and titrated using 293FT cells as targets. Figure 1B shows that the titers of concentrated Δ8.9 WT-packaged HIV-1 vector stocks ranged from 9.08 × 107 to 1.62 × 108 TU/mL, whereas 8.9 LNEIE-packaged HIV-1 vector stocks ranged from 5.66 × 107 to 8.88 × 107 TU/mL. Thus, while high titers of recombinant virus were obtained with Δ8.9 LNEIE-packaged HIV-1 vectors, the titers of Δ8.9 LNEIE-packaged HIV-1 vectors were only half of those of Δ8.9 WT-packaged HIV-1 vectors.
Meanwhile, a bicistronic expression vector was constructed containing both HA-tagged rhesus TRIM5α and GFP (Fig. 1C), and the recombinant viruses were produced and used to transduce CEMss-CCR5 cells. The Western blot analysis shows that in transduced CEMss-CCR5 cells, both GFP and HA-tagged rhesus TRIM5α proteins expressed at detectable levels compared to parental CEMss-CCR5 cells (Fig. 1D). FACS analysis shows that 99.3% of transduced CEMss-CCR5 cells were GFP positive (Fig. 1E).
To compare transduction efficiency of Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors, CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/GFP cells were transduced with various doses of Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-mCherry (MOI from 0.625 to 10). Figure 1F shows the gating strategy of representative CEMss-CCR5 and CEMss-CCR5-rhTRIM5α/GFP cells transduced with Δ8.9 LNEIE-packaged HIV-1 vector expressing GPI-mCherry at a MOI of 10. Clearly, both Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors efficiently and comparably transduced CEMss-CCR5 cells. At a MOI of 0.625, 1.25, 2.5, 5, and 10, both Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors had a 60%, 85%, 95%, 98%, and 99% transduction rate, respectively (Fig. 1G). In contrast, in CEMss-CCR-rhTRIM5α/GFP cells, the transduction efficiency by Δ8.9 WT-packaged HIV-1 vector was much lower than that of Δ8.9 LNEIE-packaged HIV-1 vectors. At a MOI of 0.625, 1.25, 2.5, 5, and 10, Δ8.9 WT-packaged HIV-1 vectors had a 6%, 25%, 34%, 38%, and 55% transduction rate, respectively, whereas Δ8.9 LNEIE-packaged HIV-1 vectors had a 24%, 55%, 68%, 91%, and 97% transduction rate, respectively (Fig. 1G), indicating that while it maintains efficient transduction of human T-cell lines, the Δ8.9 LNEIE-packaged HIV-1 vector overcomes rhesus TRIM5α restriction.
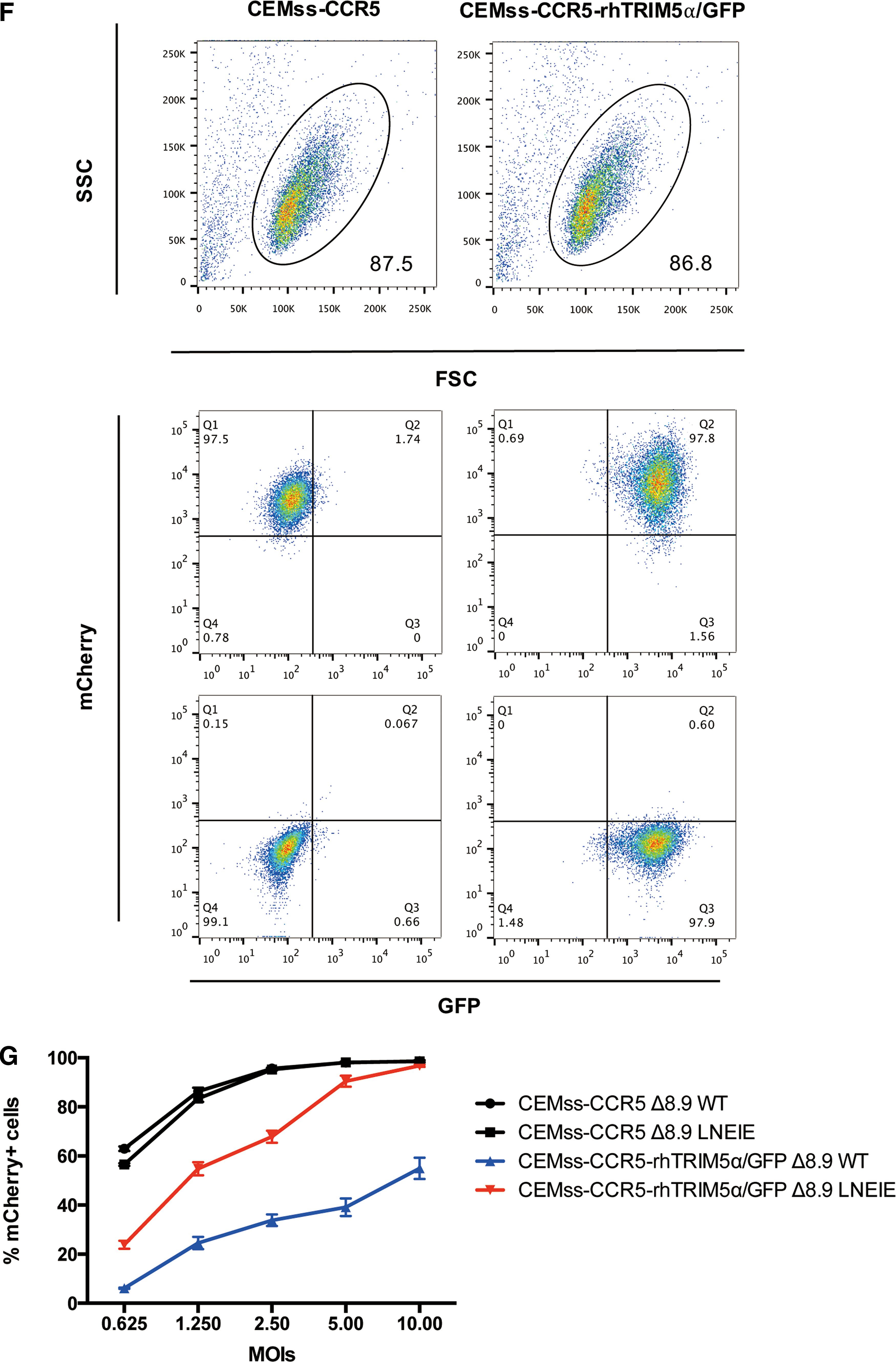
Δ8.9 LNEIE-packaged HIV-1 vector is resistant to TRIM5α-mediated capsid degradation resulting in high levels of proviral DNA
Rhesus TRIM5α redirects incoming HIV-1 CA for proteasomal degradation by directly binding CA with its B30.2/SPRY domain. Next, the study tested whether Δ8.9 LNEIE-packaged HIV-1 vector was able to avoid the disruptive effect of rhesus TRIM5α. To accomplish this, equivalent amounts of Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors were applied to CEMss-CCR5-rhTRIM5α/GFP cells in a synchronized infection protocol. At 0, 1, 2, and 4 h post transduction, cells were harvested and lysed. The amounts of CA, rhesus TRIM5α and actin were measured by Western blot analysis. Figure 2A and B shows that while similar amounts of rhesus TRIM5α and actin were detected in cells transduced with either Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors at all time points, the amounts of HIV-1 capsid protein were significantly higher and more stable in cells transduced with Δ8.9 LNEIE-packaged HIV-1 vector than with Δ8.9 WT-packaged HIV-1 vector. At 1 and 2 h post transduction, much higher amounts of CA were detected in cells transduced with Δ8.9 LNEIE-packaged HIV-1 vector than with Δ8.9 WT-packaged HIV-1 vector. Moreover, at 4 h post transduction, CA was undetectable in cells transduced with Δ8.9 WT-packaged HIV-1 vector, while the amount of CA remained relatively high in cells transduced with Δ8.9 LNEIE-packaged HIV-1 vector.
Resistance of Δ8.9 LNEIE-packaged HIV-1 vector to TRIM5α-mediated capsid degradation. (
To compare proviral DNA between cells transduced with Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors, equivalent amounts of Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors were applied to CEMss-CCR5 or CEMss-CCR5-rhTRIM5α/GFP cells in a synchronized infection protocol. At 0, 4, 8, and 16 h post transduction, genomic DNA was extracted. The amounts of proviral DNA were measured by quantitative PCR. Figure 2C shows that as expected, in CEMss-CCR5 cells transduced with Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors, similar amounts of proviral DNA were detected. In contrast, in CEMss-CCR5-rhTRIM5α/GFP cells transduced with Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors, significantly higher amounts of proviral DNA were detected in cells transduced with Δ8.9 LNEIE-packaged HIV-1 vector than in cells transduced with Δ8.9 WT-packaged HIV-1 vector. Thus, taken together, these results indicate that Δ8.9 LNEIE-packaged HIV-1 vector is resistant to rhesus TRIM5α-mediated CA degradation, resulting in higher levels of proviral DNA.
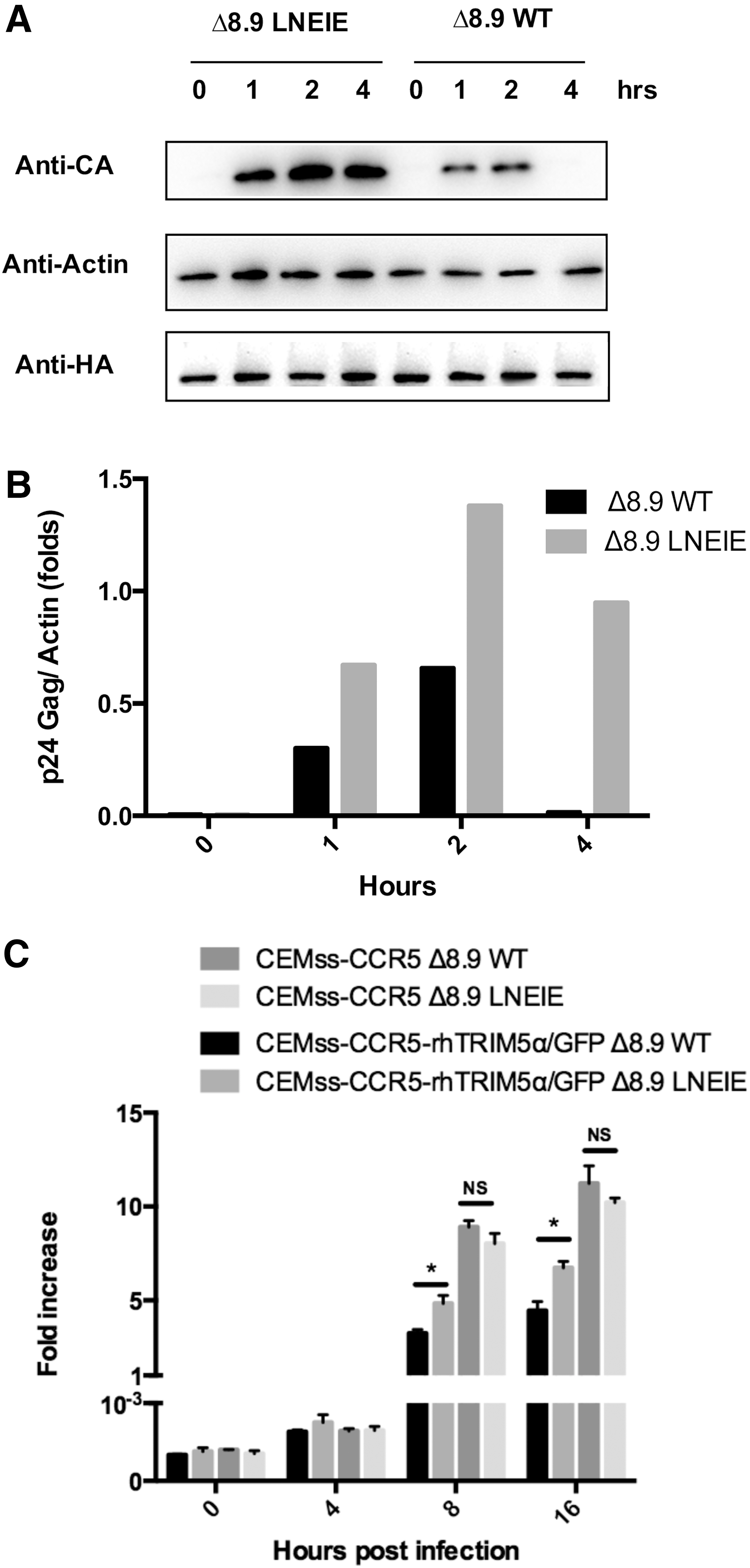
Δ8.9 LNEIE-packaged HIV-1 vector retains transduction efficiency in primary human CD4 T cells but significantly increases transduction efficiency of rhesus primary CD4 T cells
Next, the transduction efficiency was compared between Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors in human and rhesus primary CD4 T cells. To accomplish this, human and rhesus primary CD4 T cells were isolated from human and rhesus PBMCs stimulated with anti-CD3 and anti-CD28 antibody-coated beads or plates and transduced with Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors expressing GPI-scFv X5/2A/GFP. Figure 3A and B shows the transduction rates of Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors of representative human and rhesus primary CD4 T cells. Both Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors transduced human primary CD4 T cells very well, even though the transduction rates by Δ8.9 LNEIE-packaged HIV-1 vector were slightly lower than those by Δ8.9 WT-packaged HIV-1 vector. At a MOI of 2.5, 5, 10, 20, and 40, the transduction rates of Δ8.9 WT-packaged HIV-1 vector were 39.6%, 61.4%, 86.5%, 92.3%, and 93.1%, respectively, while the transduction levels of Δ8.9 LNEIE-packaged HIV-1 vector were 28%, 48.5%, 76.5%, 85.6%, and 86.2%, respectively (Fig. 3A). In contrast, although Δ8.9 WT-packaged HIV-1 vector transduced rhesus primary CD4 T cells poorly, Δ8.9 LNEIE-packaged HIV-1 vector significantly increased transduction efficiency. At a MOI of 2.5, 5, 10, 20, and 40, the transduction rates of Δ8.9 WT-packaged HIV-1 vector were 3.1%, 5.9%, 9.8%, 12.1%, and 12.3%, respectively, while those of Δ8.9 LNEIE-packaged HIV-1 vector were 28.1%, 37.9%, 43.2%, 53.9%, and 54.8%, respectively (Fig. 3B).
The comparison of transduction efficiency between Δ8.9 WT and Δ8.9 LNEIE-packaged HIV-1 vectors. The rates of transduction to human (
In addition, preliminary testing was done on transduction efficiency in anti-CD3 and anti-CD28 antibody-stimulated rhesus PBMCs transduced with Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vector at a MOI of 10. Figure 3C–E shows that Δ8.9 LNEIE-packaged HIV-1 vector significantly increased transduction efficiency in both rhesus CD4 and CD8 T cells compared with Δ8.9 WT-packaged HIV-1 vector. With Δ8.9 LNEIE-packaged HIV-1 vector, 44% of CD4 T cells and 47.7% of CD8 T cells were GFP+, whereas with Δ8.9 WT-packaged HIV-1 vector, only 6.1% of CD4 T cells and 9.4% of CD8 T cells were GFP+.
Because in both human and rhesus primary CD4 T cells the transduction levels plateaued at a MOI of 20, a MOI of 20 was used to compare the transduction efficiency between Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors in rhesus primary CD4 T cells from a cohort of 37 Chinese rhesus macaques tested. Figure 4A summarizes transduction rates between Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors in primary CD4 T cells of a cohort of 37 individual macaques. The average transduction rate by Δ8.9 WT-packaged HIV-1 vector was 8.4% (ranging from 1.1% to 18.4%), whereas the average transduction rate by Δ8.9 LNEIE-packaged HIV-1 vector was 26.5% (ranging from 10.5% to 57.8%). In each animal, Δ8.9 LNEIE-packaged HIV-1 vector outperformed Δ8.9 WT-packaged HIV-1 vector. The average fold increase in the percentage of GFP+ cells was 4.04 (ranging from 2.1% to 16.9%). Statistically, the difference in transduction rates between these two groups was extremely significant (p = 5.2 × 10–11; Fig. 4B). Taken together, these results clearly indicate that while the CA modified packaging vector maintains efficient transduction of human primary CD4 T cells, it significantly increases transduction of rhesus primary CD4 T cells.
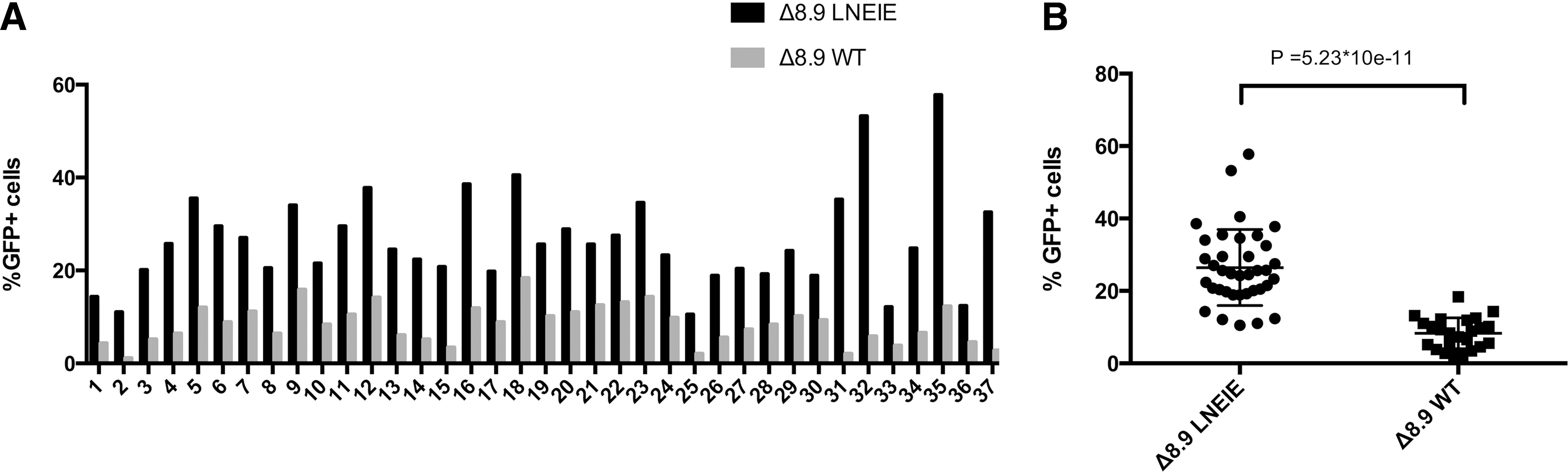
The transduction rates to rhesus primary CD4 T cells from a cohort of 37 Chinese rhesus macaque by Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors. (
Influence of TRIM5α variations in transduction efficiency of rhesus primary CD4 T cells by Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vector
Based on amino-acid variations at positions 307, 327, 333, and 339–340, rhesus TRIM5α consists of six genotypes (Manu-1, 2, 3, 4, 5, and TRIMCyp), which cluster into three types: TFP (Manu-1, 2, and 3), Q (Manu-4 and 5), and TrimCyp. 54,55 The rhesus TRIM5α variations have been shown to affect SIV infectivity and clinical outcomes in rhesus macaques. 56,57 It was hypothesized that rhesus TRIM5α variations might influence the variable transduction rates by Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors in primary CD4 T cells of the cohort of 37 Chinese rhesus macaques shown in Fig. 4. To test this hypothesis, TRIM5α alleles of these 37 Chinese rhesus macaques were evaluated using direct sequencing of 525 bp PCR product (see Materials and Methods). Among the 37 Chinese rhesus macaques, 25 had TFP/TFP (23 with Manu-3/Manu-3 and two with Manu-1 or 2/Manu-3) alleles, 11 had TFP/Q (seven with Manu-3/Manu-4 and four with Manu-3/Manu-5), and one has Q/Q (Manu-4/Manu-4) alleles. Not animals were found with a TRIMCyp genotype (Table 1). Next, the transduction rates were compared between TFP/TFP and TFP/Q groups. The average transduction rate by Δ8.9 WT-packaged HIV-1 vector in the TFP/Q group was 11.0%, while in the TFP/TFP group it was 7.38%. The difference in transduction rates between these two groups was significant (p = 0.014; Fig. 5A and Table 2). The average transduction rate by Δ8.9 LNEIE-packaged HIV-1 vector in the TFP/Q group was 35.0%, while in the TFP/TFP group it was 23.2%. The difference in transduction rates between these two groups was highly significant (p = 0.00089; Fig. 5B and Table 2). Thus, these results clearly indicate that rhesus CD4 T cells with TFP/Q alleles are more susceptible than rhesus CD4 T cells with TFP/TFP alleles to the transduction by both Δ8.9 WT and Δ8.9 LNEIE-packaged HIV-1 vectors.
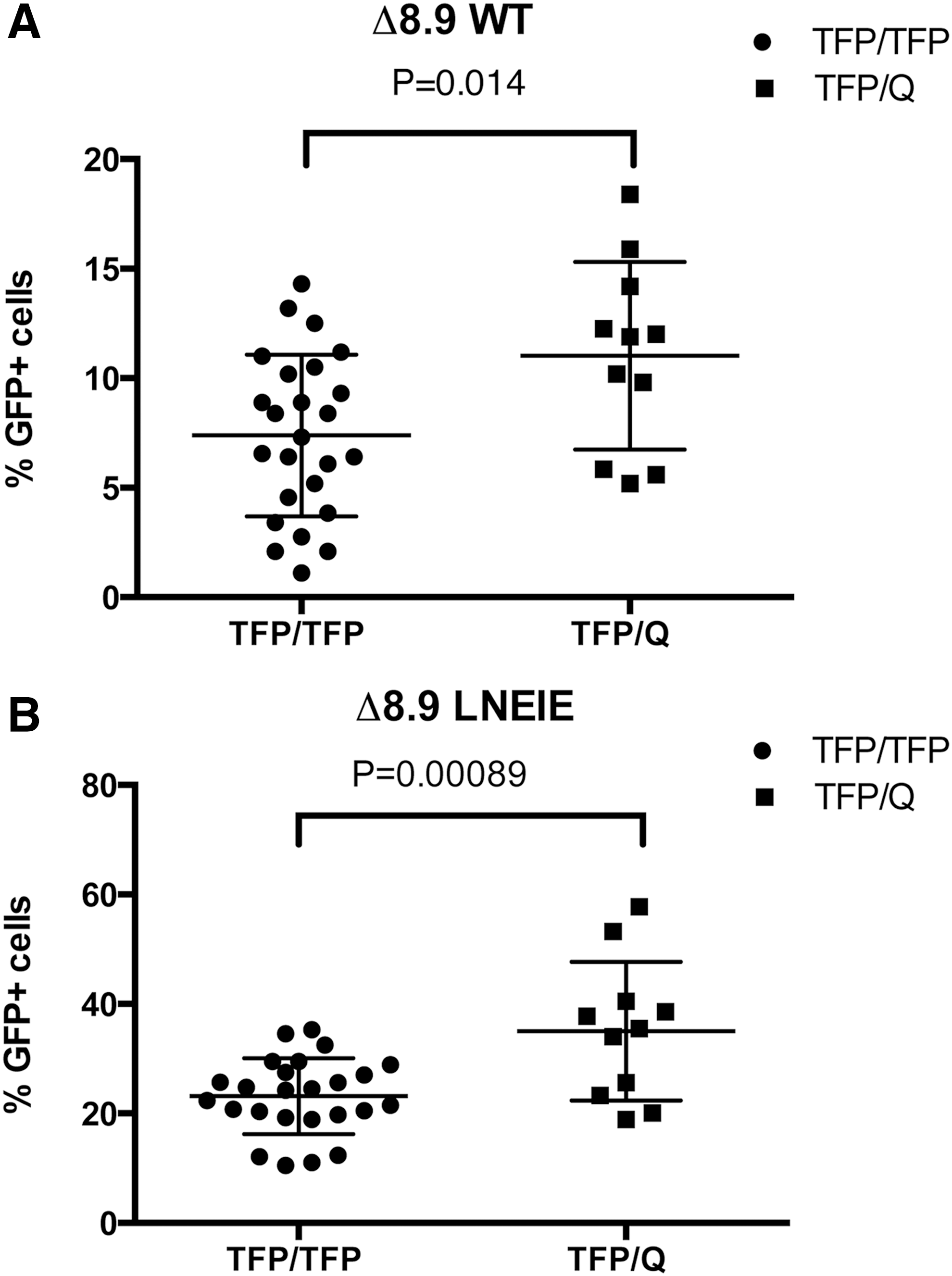
Statistical analysis of transduction rates between the TRIM5α TFP/TFP group and the TFP/Q group. (
Age, sex, weight, TRIM5α genotypes, and alleles and transduction rates of rhesus primary CD4 T cells by Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vector among a cohort of 37 individual Chinese rhesus macaques
Statistical analyses of the correlation between age, sex, weight, or TRIM5α alleles and transduction rates of rhesus primary CD4 T cells by Δ8.9 WT- or Δ8.9 LNEIE-packaged HIV-1 vector among a cohort of 37 Chinese rhesus macaques
For further statistical analysis, other host factors were also assessed that potentially affect transduction efficiency by Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vector, including the age, sex, and weight of these 37 rhesus macaques (Table 1). The results of univariate analysis show that there was no statistical difference in transduction efficiency between the different age and weight groups or between male and female animals (Table 2). Thus, taken together, it is concluded that while rhesus TRIM5α alleles has a strong influence on transduction efficiency of rhesus primary CD4 T cells by either Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vector, the age, sex, or weight of the donor animal did not influence transduction efficiency.
Discussion
Gene transfer to autologous HSCs and primary T cells utilizing HIV-1 vectors is a promising modality for the treatment of human diseases. Among nonhuman primates, the rhesus macaque has been proven to be a suitable preclinical model for safety and efficacy studies. However, species-specific restriction factors such as TRIM5α and TRIM5-CypA in rhesus macaques block HIV-1 infection, as well as significantly impair transduction efficiency of HIV-1 vectors. 33 –35 Rhesus TRIM5α and TRIM5-CypA recognize the incoming HIV-1 core by binding HIV-1 capsid and mediate its degradation, whereas SIV capsid escapes rhesus TRIM5α and TRIM5-CypA-mediated degradation. 36,37 Previously, several strategies such as the CypA-binding mutant-modified HIV-1 vectors 38,39 and a SIV capsid-packaged chimeric HIV-1 (χHIV) vector 40 have been developed to improve transduction by HIV-1-based vectors. While these strategies significantly improved transduction efficiency of rhesus CD34+ HSCs, transduction rates for rhesus primary T cells by these modified HIV-1 vectors remain suboptimal. Therefore, the present study focused on testing whether HIV-1 vectors packaged by an HIV-1 Gag/Pol expression construct containing five LNEIE mutations in CA could efficiently transduce both human and rhesus primary CD4 T cells.
One interesting finding in the present study is that while it maintains efficient transduction to human T-cell lines and human primary CD4 T cells (Figs. 1F and G and 3A), Δ8.9 LNEIE-packaged HIV-1 vector indeed overcomes rhesus TRIM5α restriction (Fig. 1F and G), resulting in significantly higher transduction efficiency to rhesus primary CD4 T cells compared with Δ8.9 WT-packaged HIV-1 vector (Figs. 3B and 4). Moreover, the preliminary data also show that Δ8.9 LNEIE-packaged HIV-1 vector can also efficiently transduce both rhesus primary CD8 and CD4 T cells (Fig. 3C–E). Thus, taken together, it is concluded that Δ8.9 LNEIE-packaged HIV-1 vector can indeed efficiently transduce human and rhesus primary T cells.
Several lines of evidence indicate that the HIV-1 capsid remains intact as it traverses the cytoplasm of newly infected cells. Prematurely uncoated incoming HIV-1 core triggers cytosolic sensors and nucleases, 58 assembled capsid proteins are required for properly engaging the nuclear pore complex, 59 and intact capsids have been observed at the nuclear envelope. 60 More recently, it has been shown that each capsid hexamer contains a size-selective pore bound by a ring of six arginine residues and a “molecular iris” formed by the N-terminal β-hairpin, which import nucleotides and fuel encapsidated DNA synthesis. 61 Rhesus TRIM5α and TRIM5-CypA recognize the incoming HIV-1 core by binding HIV-1 capsid with their C-terminal B30.2/SPRY and cyclophilin A (CypA) domains and then mediate the degradation of HIV-1 core complex. 36,37 The present study shows the significantly higher and more stable HIV-1 capsid, along with significantly higher proviral DNA detected in CEMss-CCR5-rhTRIM5α/GFP cells transduced with Δ8.9 LNEIE-packaged HIV-1 vector than with Δ8.9 WT-packaged HIV-1 vector (Fig. 2A–C). Thus, the results not only indicate the resistance of LNEIE CA mutant to rhesus TRIM5α/TRIM5-CypA-mediated degradation, but also suggest that the intact HIV-1 core complex of Δ8.9 LNEIE-packaged HIV-1 vector leads to a higher amount of proviral DNA.
Another interesting finding in the present study is that allelic variation in TRIM5α strongly influences transduction efficiency of rhesus primary CD4 T cells by both Δ8.9 WT or Δ8.9 LNEIE-packaged HIV-1 vectors. Among the cohort of 37 Chinese rhesus macaques tested in the present study, there were 25 TFP/TFP, 11 TFP/Q, one Q/Q, but no TrimCyp/TrimCyp, TrimCyp/TFP, or TrimCyp/Q alleles (Table 1). The allelic frequency found in these 37 Chinese rhesus macaques is consistent with two previous reports. 54,62 When transduction efficiency was compared between TFP/TFP and TFP/Q groups, the transduction efficiency by both Δ8.9 WT and Δ8.9 LNEIE-packaged HIV-1 vectors was significantly higher in the TFP/Q group than it was in the TFP/TFP group (p = 0.014 and 0.00089, respectively; Fig. 5A and B and Table 2). Although at this time there is no explanation for this observation, the results are consistent with the study reported by Evans et al. 42 They transduced cell lines expressing six different rhesus TRIM5 genotypes (Manu-1, -2, -3, -4, -5, and TRIM5-CypA) with chimeric χHIV-1 vector expressing GFP and found that transduction efficiency is much higher in cell lines expressing Q (Manu-4 and -5) and TrimCyp alleles than in it is in cell lines expressing the TFP allele (Manu-1, -2, and -3). In addition, they also found higher transduction rates in CD34+ HSCs from TFP/Q (Manu-3/Manu-4) alleles than in TFP/TFP (Manu-3/Manu-3) alleles.
Finally, although significant improvement of transduction efficiency of rhesus primary CD4 T cells has been achieved with Δ8.9 LNEIE-packaged HIV-1 vector, transduction rates of rhesus primary CD4 T cells are still lower than those of human primary CD4 T cells (Fig. 3). It is speculated that this could be due to the presence of other species-specific restriction factors in rhesus primary CD4 T cells that partially block the Δ8.9 WT-packaged HIV-1 vector. Besides rhesus TRIM5α/TRIM5-CypA, rhesus APOBEC3G and tetherin proteins have been shown to block HIV-1 infection in macaque cells, which can be antagonized by SIV Vif and Vpu, respectively. 32,63 –65 Interestingly, Nomaguchi et al. 66 showed that a HIV-1 variant MN4/LSDQgtu, which combines LSDQ CA mutations and the substitution of the transmembrane domain of SIVgsn166 Vpu, grew comparably to SIVmac239 in rhesus PBMCs. Soll et al. 46 showed that a HIV-1 variant encoding the LNEIE CA substitutions and SIVmac239 Vif replicated efficiently in rhesus primary lymphocytes. Thus, it may be worthwhile to modify Δ8.9 WT-packaged HIV-1 vector further so as to improve transduction efficiency of rhesus primary T cells further. Nevertheless, the Δ8.9 LNEIE packaging construct provides improved transduction of rhesus cells, which should enhance the utility of HIV-1-based vectors in this nonhuman primate species.
Footnotes
Acknowledgments
The authors wish to thank Dr. L. Naldini at the University Torino Medical School, Torino, Italy, for lentiviral transfer vector pRRLsin-18.PPT.PGK.Wpre. This work was supported by research grants from the Chinese National Science Foundation (#31170871) and National Science and Technology Major Project (#2014ZX10001-001) to P.Z.; by research grants from Research Program of Chinese Ministry of Health (#201402001) to C.Q.; by research grants from National Science and Technology Major Project (#2014ZX10001001-001-004 and #2014ZX10001001-002-006) to J.X.; by research grants from National Institutes of Health (R56 AI108467) to J.T.K. and the Baylor-UTHouston CFAR (P30 AI036211); and by Chinese National Science Foundation—National Institutes Health joint grant (#81361120406—R01 AI106574) to P.Z. and J.T.K. The funders had no role in the study design, data collection and interpretation, or the decision to submit the work for publication.
Author Disclosure
None of the authors have any conflicts of interest to report.