
Abstract
Previously, the authors developed an adenoviral vector, Ad-PG, where transgene expression is regulated by a p53-responsive promoter. When used to transfer the p53 cDNA, a positive feedback mechanism is established. In the present study, a critical comparison is performed between Ad-PGp53 and AdRGD-PGp53, where the RGD motif was incorporated in the adenoviral fiber protein. AdRGD-PGp53 provided superior transgene expression levels and resulted in the killing of prostate carcinoma cell lines DU145 and PC3. In vitro, this effect was associated with increased production of cytoplasmic and mitochondrial oxidants, DNA damage as revealed by detection of phosphorylated H2AX, as well as cell death consistent with apoptosis. Differential gene expression of key mediators of reactive oxygen species pathways was also observed. Specifically, it was noted that induction of known p53-target genes Sestrin2 and PIG3, as well as a novel target, NOX1, occurred in PC3 cells only when transduced with the improved vector, AdRGD-PGp53. The participation of NOX1 was confirmed upon its inhibition using a specific peptide, resulting in reduced cell death. In situ gene therapy also resulted in significantly improved inhibition of tumor progression consistent with oxidant-induced DNA damage only when treated with the novel AdRGD-PGp53 vector. The study shows that the improved adenovirus overcomes limitations associated with other p53-expressing vectors and induces oxidant-mediating killing, thus supporting its further development for cancer gene therapy.
Introduction
N
Gene therapy may provide an alternative to the conventional treatment of cancer. A typical strategy involves virus-mediated transfer of a tumor suppressor gene in order to induce tumor cell death. P53, the most well-studied tumor suppressor, is a transcription factor that acts by promoting cell cycle arrest and apoptosis in response to DNA damage, ultimately preventing the propagation of cells harboring extensive genetic alterations. 2 Not only does p53 act as the “guardian of the genome,” preventing transformation of damaged cells, 3 but it also can often confer these and other desirable antitumor activities when introduced into a pre-existing tumor cell. Thus, p53 is an obvious and well-studied choice for cancer gene therapy. In fact, an adenovirus vector expressing p53 (Gendicine) has been commercially available for the treatment of head and neck cancer in China since 2003. 4,5 Even though adenoviral vectors expressing p53 have been tested in vitro and in vivo for prostate cancer treatment, 6 –13 their clinical application is still limited due, in part, to poor or variable vector performance.
Oxidants are mainly generated as sub-products of enzymatic reactions that produce, for example, hydrogen peroxide, superoxide anion, and hydroxyl radicals, which oxidize and damage proteins, lipids, and DNA. 14 In particular, nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, including NOX1, are responsible for the production of superoxide. Low-level intracellular reactive oxygen species (ROS) can cause DNA damage leading to mutation, thus promoting tumorigenesis. On the other hand, elevated levels of oxidants contribute to extensive DNA damage, mitochondrial membrane permeabilization, activation of apoptotic signaling, and induction of cell death. 15 –17 Several drugs that induce apoptosis of cancer cells rely on ROS induction. 14,18 Indeed, an essential component of p53's mechanism of action is the induction of ROS. 19 –22 ROS (peroxide and superoxide) induction mediated by p53 was shown to induce apoptosis independently of cytochrome c-release and can regulate mitochondrial membrane potential. 23 Conversely, induction of p53 by ROS has been shown to mediate necrotic cell death through PARP activation. 24 In this way, p53 plays a role in both cause and effect of the ROS response.
The use of adenoviral vectors for cancer gene therapy targeting the p53 pathway continues to be investigated. The authors' group has incorporated a p53-responsive element (PG) in a variety of viral vectors, resulting in high levels of transgene expression controlled by p53. 25 –27 It has previously been shown that Ad-PGp53—a non-replicating serotype 5 adenoviral vector expressing p53 under the control of the p53-responsive promoter—confers a positive feedback mechanism that supplies significantly higher levels of p53 protein expression than seen with a typical CMV promoter when applied to the PC3 and DU145 human prostate carcinoma cell lines. Ad-PGp53 was far superior to Ad-CMVp53 for the inhibition of proliferation and induction of cell death in vitro and in vivo. However, due to viral tropism, the effects of Ad-PGp53 on the PC3 cell line were discrete, while DU145 readily succumbed to this treatment. 28
Tropism of adenovirus serotype 5 is mediated by interaction of the fiber protein with the coxsackievirus and adenovirus receptor (CAR). Yet, incorporation of an arginine-glycine-aspartic acid (RGD) motif into the HI loop of the fiber knob domain obviates the dependence on CAR and redirects vector tropism to integrins, especially αvβ3 and αvβ5. 29,30 While the prostate cancer cell line DU145 expresses high levels of CAR and medium to low levels of integrins, PC3 presents low levels of CAR and higher levels of integrins. 31 Thus, transduction of PC3 cells may be severely hampered unless a modified vector is used.
Herein, a critical advance in the TP53 gene transfer strategy is reported: the construction and successful application of an improved adenoviral vector, AdRGD-PGp53, which features the autoregulatory expression of p53 as well as the RGD motif into the adenovirus fiber protein in order to facilitate transduction. Increased p53 expression and extensive cell death associated with ROS induction and apoptosis in the resistant PC3 cell line is shown when treated with AdRGD-PGp53 but not Ad-PGp53. Strikingly, enhanced p53 gene transfer also correlated with the differential regulation of known p53 target genes and NOX1, a novel putative p53 responsive gene. With the improved AdRGD-PGp53 vector, it was observed that induction of ROS distinguished the cellular susceptibility to TP53 gene transfer.
Materials and Methods
Cell lines
HEK293, DU145 (heterozygous p53 mutant, P223L, V274F), and PC3 (p53 null) human PCa (prostate cancer) cell lines were cultivated in Dulbecco's modified Eagle's medium with 10% fetal calf serum, supplemented with antibiotic-antimycotic (Life Technologies, Carlsbad, CA) and maintained at 37°C and 5% CO2 atmosphere.
Construction of adenoviral vectors
The Ad-CMVLacZ vector was obtained commercially (Life Technologies). The AdRGD-CMVLacZ vector was donated by Dr. Hiroyuki Mizuguchi, Osaka University, Japan.
Ad-PGp53, a non-replicating recombinant adenovirus serotype 5 vector encoding the human wild-type TP53 cDNA under the control of a p53-responsive promoter, termed PG, is described elsewhere. 28 The construction of Ad-PGLuc followed the similar procedure as per Ad-PGp53 and has been described previously. 32
For the construction of AdRGD-PGp53, first the pAdHM15-RGD vector 30 (kindly provided by Dr. Hiroyuki Mizuguchi, Osaka University, Japan) was modified so that it would be compatible with the Gateway recombination strategy, creating the pAdRGD-promoterless/dest construct. Next, site-directed recombination was performed using pENTR/TOPO-PGp53 and pAdRGD-promoterless/dest in the presence of Gateway L/R Clonase II Enzyme mix (Life Technologies). Further information is available upon request and as described in Hunger et al. 33
Adenovirus production
Production of Ad5 vectors (native fiber) was done using two rounds of CsCl purification, as described previously. 32 Adenoviral vectors modified with the RGD motif were purified as per Peng et al. 34 Briefly, virus containing cellular lysate was purified by iodixanol density gradient ultracentrifugation for 1 h at 10°C and 35,000 rpm (rotor Sorvall TH-G41), followed by purification on a PD-10 desalting column with Sephadex™ G-25 (GE Healthcare Life Sciences, Pittsburgh, PA).
The biological titer (expressed as transducing units per milliliter [TU/mL]) was determined functionally by detection of hexon protein in transduced cells with the aid of the Adeno-X Rapid Titer Kit (Clontech, Mountain View, CA). Multiplicities of infection (MOIs) are calculated considering the number of transducing units presented per cell.
Western blot
DU145 and PC3 cell lines were seeded in six-well plates, 1 × 105 cells/well, and transduced with adenovirus using MOIs from 50 to 1,000. Proteins were extracted, quantified by the Bradford method, loaded in 12% SDS-PAGE, transferred onto nitrocellulose, and incubated with rabbit anti-p53 (SC6243; Santa Cruz Biotechnology, Santa Cruz, CA), rabbit anti-p21Waf1 (CDKN1A; SC397; Santa Cruz Biotechnology), mouse anti-PARP (9542; Cell Signaling, Danvers, MA), or mouse monoclonal anti-β-actin (A5316; Sigma–Aldrich, St. Louis, MO) antibodies. The Amersham ECL Prime Western Blotting Detection Reagent (GE Healthcare Life Sciences) was used for chemiluminescent detection with the aid of an Image Quant LAS 4000 (GE Healthcare Life Sciences).
Cell viability, cell death assays, detection of β-galactosidase activity
Standard MTT, detection of β-galactosidase activity, cell cycle, and Annexin-V/PI procedures were used, as per previous studies. 28,32,35
Detection of caspase activity
In 12-well plates, 5 × 104 DU145 and PC3 cells were seeded and transduced as described above. After 48 or 72 h of incubation, the cells were analyzed using the CellEvent Caspase-3/7 Green Detection Reagent (Life Technologies) and observed under a fluorescence microscope (Evos FL; Life Technologies). Using ImageJ software, the positive (green) and total cells from at least 10 fields were counted.
Real-time polymerase chain reaction
One hundred thousand PC3 or DU145 cells were transduced with the adenoviral vectors, and after 48 h of incubation, total RNA was harvested using TRIzol (Life Technologies). cDNAs were generated from 1 μg of total RNA using MMLV reverse transcriptase (Life Technologies). Amplifications were carried out using SYBR Green I mix (Life Technologies) on a 7500 ABI Applied Biosystems (Life Technologies) using 0.36 μL of specific primers (Supplementary Table S1; Supplementary Data are available online at
DHE staining
One hundred thousand PC3 or DU145 cells were transduced with the adenoviral vectors, and after 48 h of incubation, the cells were stained with 50 μM of dihydroethidium (DHE) for 30 min. After the incubation period, the cells were washed twice with phosphate-buffered saline (PBS), trypsinized, and analyzed by flow cytometry (FACScan; BD Biosciences, San Jose, CA). For treatment with ROS inhibitors, cells were incubated overnight in the presence of PEG-catalase at a concentration of 200 IU/mL.
Cellular H2O2 production
One hundred thousand PC3 or DU145 cells were transduced with the adenoviral vectors, and after 48 h of incubation, the cells were trypsinized and divided into two wells for treatment or not with catalase (200 IU/mL). Together with this treatment, cells were incubated with AmplexRed reagent (0.1 mM) and horseradish peroxidase (1 IU/mL) in Krebs buffer (0.5 mM of CaCl2; 1.2 mM of MgSO4; 4.9 mM of KCl; 5.7 mM of KH2PO4; 145 mM of NaCl; 5.7 mM of Na2HPO4; 5.5 mM of glucose; pH 7.4) containing DTPA (0.1 mM) for 45 min at 37°C. Fluorescence was observed (excitation/emission wavelengths 480/560 nm) using a spectrofluorometer (SpectraMax M5; Molecular Devices, Sunnyvale, CA).
Inhibition of NOX1
For treatment with a NOX1-specific synthetic peptide inhibitor, cells were incubated daily in the presence of 10 μM of NoxA1ds or scramble (SCRMB) peptides. The authors thank Dr. Robson Lopes de Melo at the Special Laboratory of Applied Toxicology at Butantan Institute for synthesis of the peptides. NoxA1ds sequence: NH3-EPVDALGKAKV-CONH2, SCRMB sequence: NH3-LVKGPDAEKVA-CONH2. 36 After 72 and 48 h of incubation, PC3 and DU145 cells, respectively, were collected, fixed, and subjected to PI staining to reveal hypodiploid cells.
Mitochondrial membrane potential (JC-1)
One hundred thousand PC3 or DU145 cells were transduced with the adenoviral vectors and incubated for 72 or 48 h, respectively. The cells were washed with PBS, trypsinized, and incubated for 15 min with BD Mitoscreen JC-1 (BD Biosciences). After this period, the cells were washed twice with 1 × assay buffer and analyzed by flow cytometry (Attune; Life Technologies).
Mouse model of in situ gene therapy
Athymic male nude BALB/c mice, 8–12 weeks old, were used in the in vivo experiment. Animal experimentation was performed at the Gene Therapy Laboratory, Department of Cell and Developmental Biology, Biomedical Sciences Institute, University of São Paulo. The experimental protocols, as described here, were approved by the Ethics in Animal Use Committee of the Institution. PC3 cells (2 × 106) were implanted subcutaneously in the flank of the mice. Twice per week, the tumors were measured with calipers, and the volume was calculated according to the formula 0.5 × (major diameter) × (minor diameter)2. 37 After 3 weeks, the tumors reached 60 mm3, and treatment was initiated. The concentration of adenovirus injected intratumorally was 1 × 109 TU/100 μL 1 × PBS/injection, and this treatment was repeated a total of six times at 48 h intervals. Tumor progression was monitored over time, but animals were euthanized when the tumor volume reached 1,000 mm3. In all procedures, including euthanasia by CO2 inhalation, the mice were anesthetized in a chamber with 4% isoflurane.
Immunofluorescence
Tumors from the in vivo assay were collected 48 h after the final round of gene transfer, fixed with 4% paraformaldehyde for 4 h, and incubated in 30% sucrose solution for 16 h at 4°C. After this time, the tumors were frozen in OCT-Tissue Tek, cut in 4 μm sections using a cryostat, and placed on silanized glass slides. p-H2AX (ser-139) protein was detected with a mouse monoclonal antibody (05-636; Millipore, Billerica, MA). The secondary anti-mouse antibody was conjugated with Alexa 594 (Life Technologies), and nuclear staining was performed with Hoechst 33258 (Life Technologies) at a concentration of 20 mg/mL. The cells were analyzed by fluorescence microscopy (Carl Zeiss, Oberkochen, Germany).
Statistical analysis
Statistical analyses were performed using GraphPad Prism software v5.0. Statistical significance was calculated using two-way analysis of variance and Bonferroni post-test, and significance was considered when p < 0.05.
Results
Comparison of p53 expression mediated by AdRGD-PGp53 and AdPGp53
In previous studies by the authors' group, it was noted that transduction of PC3 cells with non-replicating Ad5 vectors was quite inefficient, while DU145 cells were readily transduced. 28 To explore whether a modified vector containing the RGD tripeptide in the H1 loop of the fiber protein would confer increased viral entry, DU145 and PC3 cells were transduced with Ad-CMVLacZ and AdRGD-CMVLacZ (Supplementary Fig. S1). DU145 showed similar transduction efficiency with either vector. Yet, PC3 showed higher efficiency with AdRGD-CMVLacZ, indicating that the altered fiber protein does indeed increase adenoviral transduction of PC3 cells. Next, the RGD-modified adenovirus platform was used for the construction of the AdRGD-PGp53 vector, which expresses the human wild-type TP53 cDNA under the control of a p53-responsive promoter.
The PC3 and DU145 prostate carcinoma cell lines were used to evaluate the expression of p53 mediated by AdRGD-PGp53 and Ad-PGp53. PC3 cells were transduced with a MOI of 1,000 and harvested 24, 48, and 72 h after transduction (Fig. 1A). AdRGD-PGp53 conferred superior levels of p53 expression, even at the earliest time point. Transduction with AdRGD-PGp53 confers induction of p21Waf1 at 48 h, and cleaved PARP is clearly observed after 72 h. Full-length PARP was not significantly altered (data not shown). Similarly, transduction of DU145 cells with AdRGD-PGp53 yielded greater p53 expression levels from the modified vector compared with Ad-PGp53. Yet, both conferred similar levels of cleaved PARP, and only AdRGD-PGp53 showed high levels of p21Waf1 expression (Fig. 1B). In this way, it is shown that by increasing the transduction efficiency, the level of p53 expression was also elevated, as expected.
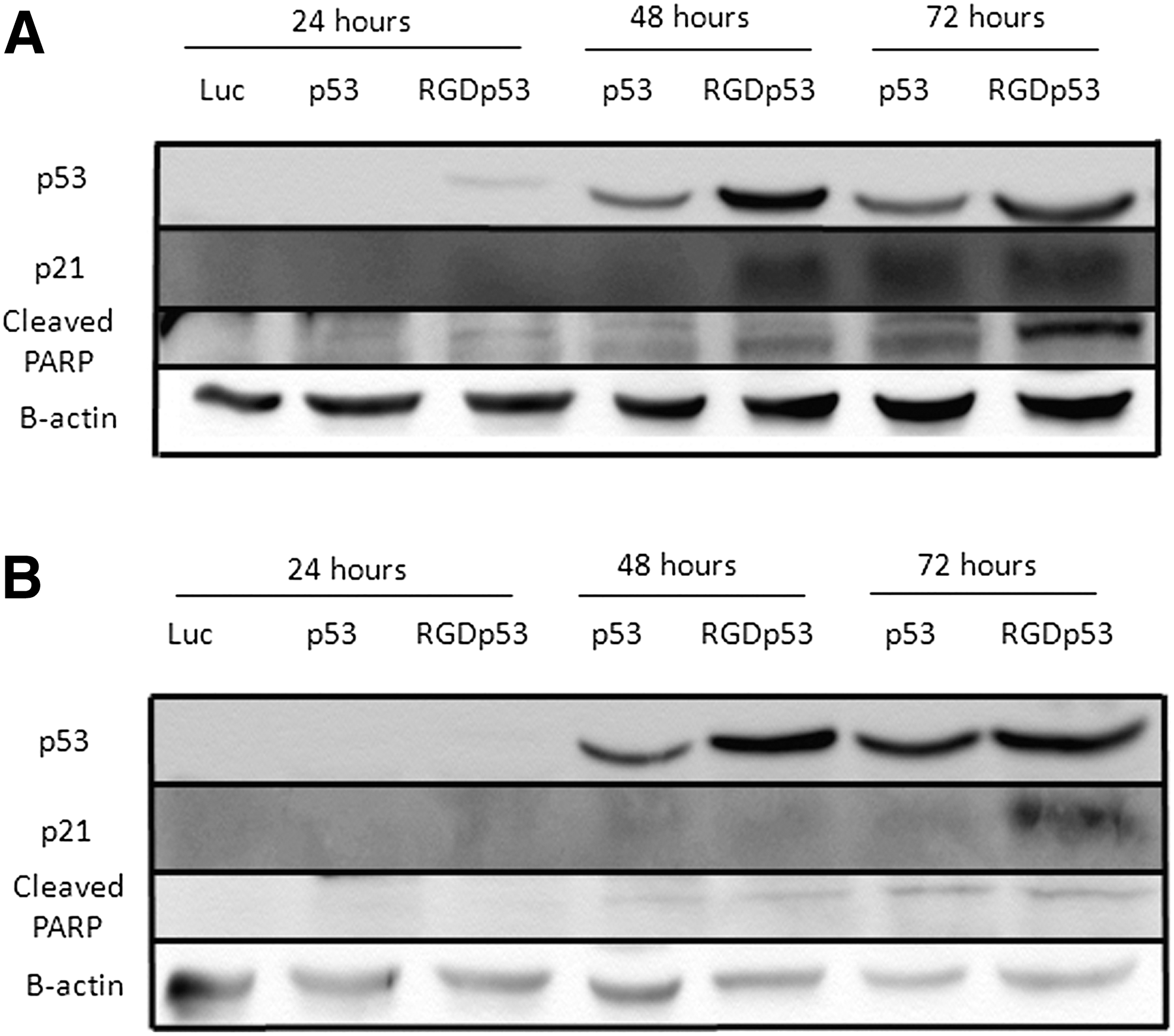
Detection of p53 protein in PCa cell lines transduced with adenoviral vectors. (
Cell cycle alterations and apoptosis mediated by the adenoviral vectors
In the authors' previous work, PC3 cells were quite resistant to cell death upon treatment with Ad-PGp53. In striking contrast, the AdRGD-PGp53 vector that confers higher levels of p53 protein was associated with reduced metabolic activity (MTT assay), impaired cell proliferation, as well as accumulation of hypodiploid and apoptotic populations—responses not seen with Ad-PGp53 at the same MOI (Fig. 2). Even when using Ad-PGp53 at a MOI five times greater than AdRGD-PGp53, the non-modified vector provided only a modest accumulation of hypodiploid cells (Supplementary Fig. S2).
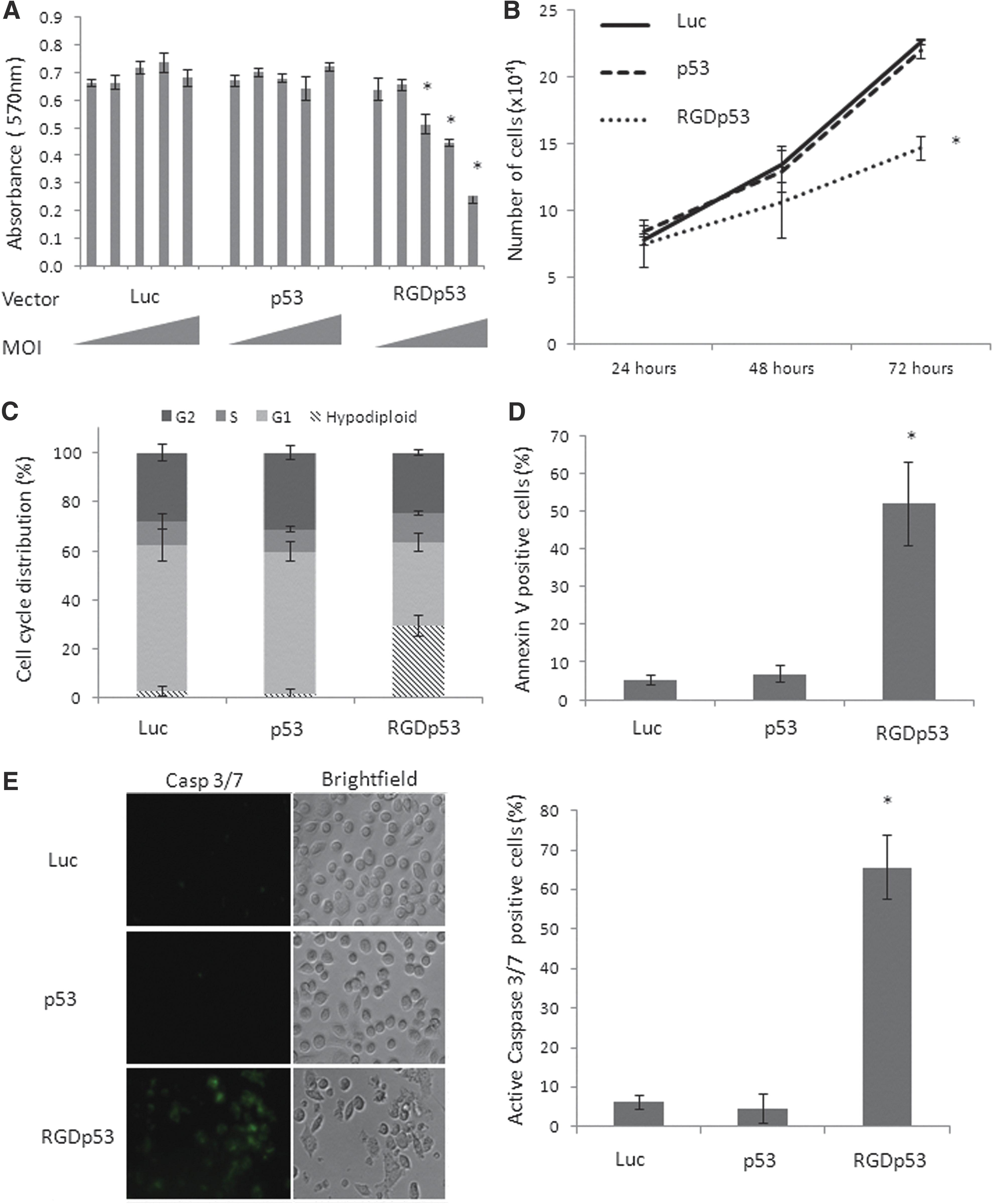
Functional assays reveal the impact of adenovirus-mediated gene transfer in PC3 cells. (
DU145 showed reduction of metabolism, decreased growth, and an increase in apoptosis mediated by both AdRGD-PGp53 and Ad-PGp53 (Fig. 3), consistent with previous findings, indicating that DU145 cells are quite susceptible to treatment with p53, even when using the p53-responsive vector without fiber modification. 28
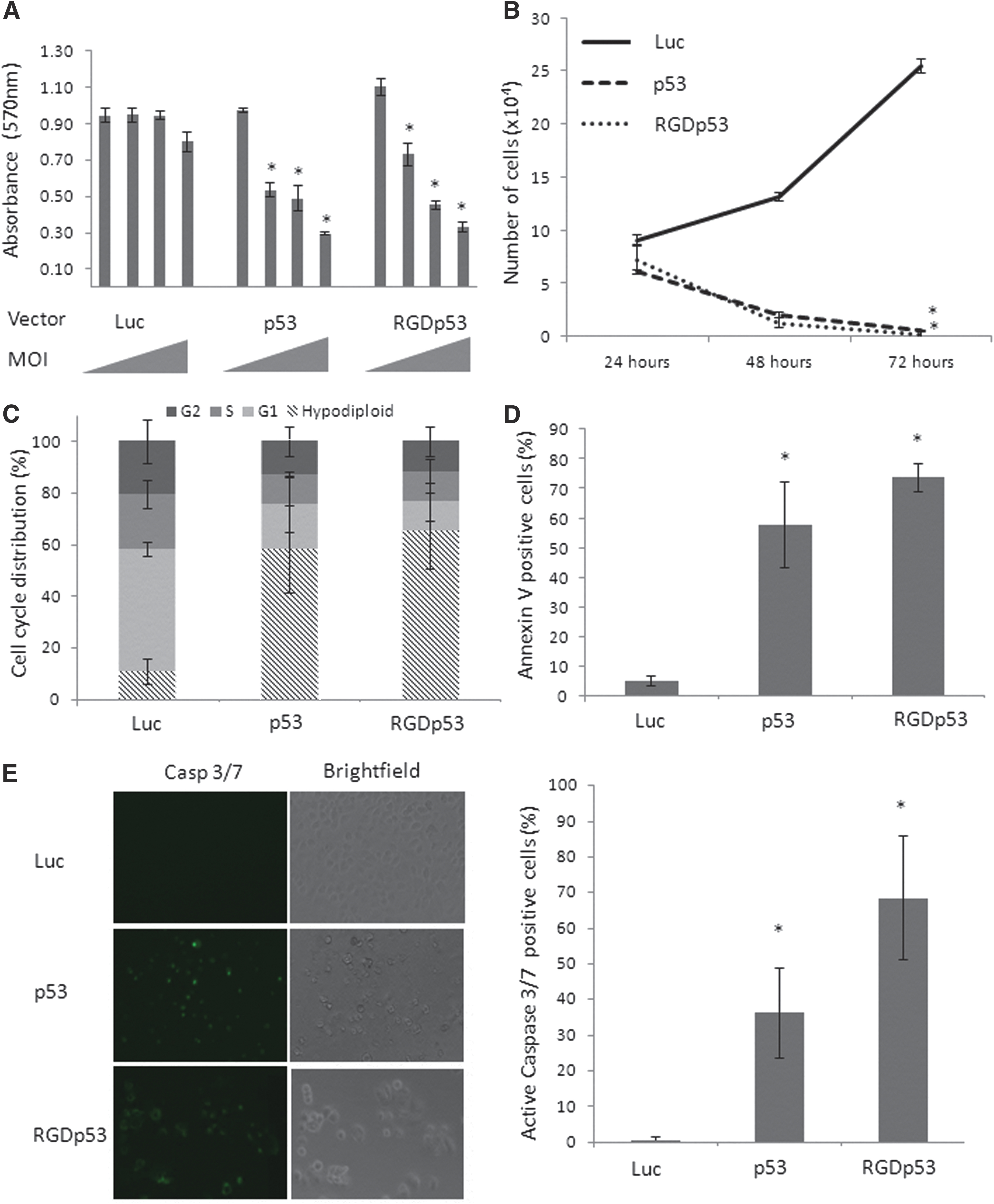
Functional assays reveal the impact of adenovirus-mediated gene transfer in DU145 cells. Legend as per Fig. 2, except that MOIs of 10, 100, 250 and 500 were used in panel (
Induction of oxidants mediated by p53 overexpression
Since p53 may act through a variety of mechanisms in order to induce cell death, the study aimed to characterize further the functional responses upon gene transfer with the adenoviral vectors, including the production of oxidants. DHE, a blue, non-fluorescent, cell permeable dye that reacts with superoxide and forms red fluorescent ethidium and 2-hydroxyethidium compounds that intercalate with DNA, was used. Figure 4A shows that treatment with AdRGD-PGp53 conferred an increase in red fluorescence for both PC3 and DU145, while Ad-PGp53 only increased superoxide in DU145 cells. The low-intensity peaks seen in the treated DU145 cells were associated with the low complexity and small-sized cell populations, likely representing hypodiploid cells (data not shown). Cellular H2O2 production was measured with Amplex Red Reagent (Fig. 4B), showing markedly increased levels upon gene transfer that were significantly impeded in the presence of catalase. Next, the study evaluated the functional effect of H2O2 inhibition, using PEG-catalase, on the cellular response to viral transduction. As measured by PI staining, PEG-catalase reduced the hypodiploid population provoked by the treatment of PC3 and DU145 cells with Ad-PGp53 or AdRGD-PGp53 (Fig. 4C). These findings suggest that superoxide/H2O2 levels induced by p53 expression are associated with cell death.
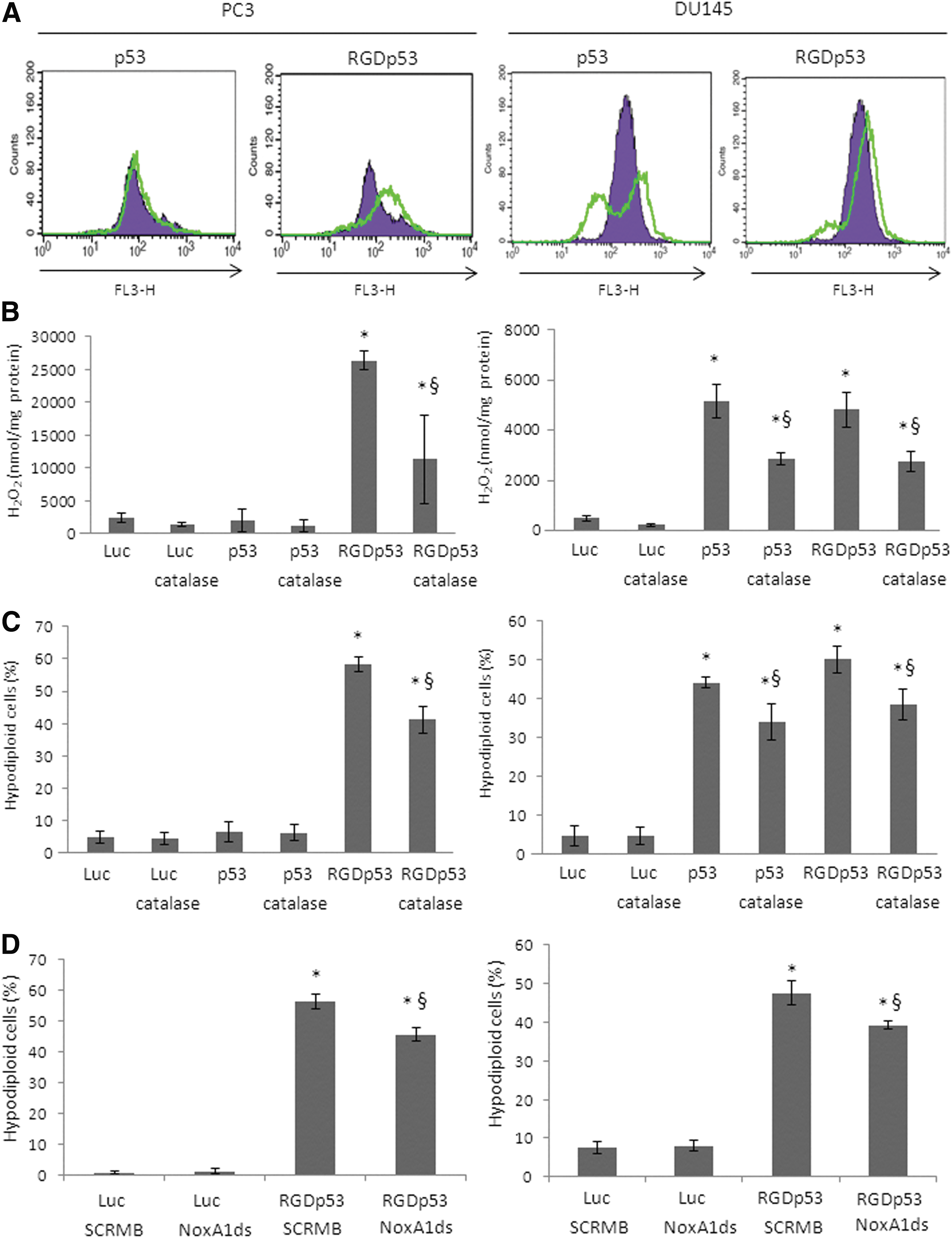
Reactive oxygen species (ROS) induction in response to p53 gene transfer. PC3 (left panel) or DU145 cell lines (right panel) were transduced with Ad-PGLuc (Luc), Ad-PGp53 (p53), or AdRGD-PGp53 (RGDp53) with a MOI of 500 (PC3) or 50 (DU145). (
Since NOX1 plays a critical role in generating superoxide/H202, next the study asked whether inhibition of NOX1 would reduce cell death associated with the gene transfer approach. Using a NOX1-specific synthetic peptide inhibitor (NoxA1ds), again a reduction in the hypodiploid population induced by AdRGD-PGp53 in PC3 and DU145 cells was observed (Fig. 4D). This finding corroborates the data with PEG-catalase and indicates that NOX1 plays a pivotal role in the induction of oxidants and, as a result, cell death.
Alteration of the mitochondrial membrane potential (Δψ) was conferred by AdRGD-PGp53 in PC3 and DU145, while Ad-PGp53 only altered potential in DU145 cells, as observed as a reduction in the JC-1 aggregate and an increase in JC-1 monomers (Fig. 5A). Consistent with the induction of oxidants, AdRGD-PGp53 caused DNA damage, as indicated by the increase in p-H2AX in both cell lines, and Ad-PGp53 caused DNA damage only in DU145 cells (Fig. 5B). Together, these functional assays implicate that oxidant induction distinguishes the cellular susceptibility to p53 gene transfer and point to AdRGD-PGp53 as an effective means to bring about this effect.
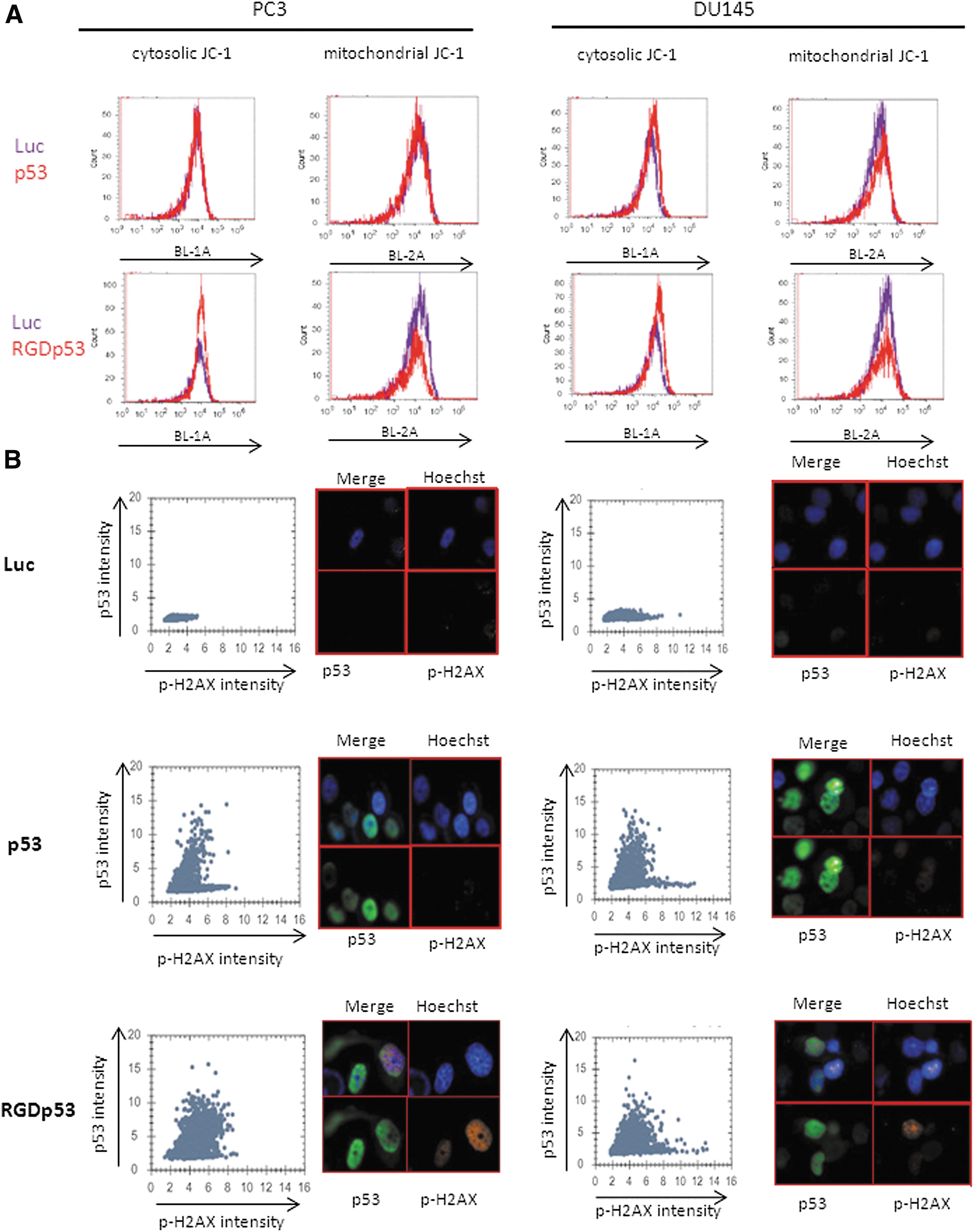
Mitochondrial and DNA damage induced by p53 gene transfer. (
Impact of gene transfer on the mRNA expression from p53 target genes
Often, p53 exerts its effects through transcriptional regulation of a variety of target genes. In this study, quantitative PCR was used to examine the expression of key p53-responsive genes related to cell death and ROS production (Fig. 6). Strikingly, in PC3 cells, differential gene expression of p21 Waf1 (CDKN1A), NOXA (PMAIP1), Sestrin2 (SESN2), PIG3 (TP53I3), and NOX1 was seen depending on the vector used. The induction of NOX1 is especially intriguing not only due to its importance in the ROS response, but also since it is not a known transcriptional target of p53. In contrast, the induction of these genes in DU145 was confirmed but was not significantly influenced by the choice of vector. Induction of GADD45α or suppression of Bcl-2 (BCL2) tended toward the vector-specific pattern in PC3 but not DU145 (Supplementary Fig. S3). Downregulation of Survivin (BIRC5) and MDM4 was seen in both cell types and did not appear to correlate to the vector used (Supplementary Fig. S4). No significant alteration in OKL38 (OSGIN1) or PIG8 (TP53I8) expression was seen (Supplementary Fig. S5).

Differential gene expression in response to treatment with the adenoviral vectors in PC3 and DU145 cell lines. The cells were transduced with Ad-PGLuc (Luc), Ad-PGp53 (p53), or AdRGD-PGp53 (RGDp53) at a MOI of 500 (PC3) or 50 (DU145). After 24 or 48 h of incubation, respectively, cells were harvested and subjected to real-time polymerase chain reaction analysis. Graphs show relative expression of the genes compared to the control group (Luc). *Indicates samples that show significant statistical difference compared to the control. §Indicate samples that show significant statistical comparing AdRGDPGp53 and AdPGp53.
These results indicate that the resistance of PC3 cells to transduction with the non-modified vector were overcome when the AdRGD-PGp53 vector was employed, causing alteration in oxidant levels, differential expression of key regulatory genes in the oxidant pathways (Sestrin2, PIG3, NOX1), and induction of DNA damage. In contrast, the oxidant response in DU145 cells was induced by either the modified or non-modified adenoviral vector encoding p53. In this way, induction of oxidants distinguished the cellular responsiveness to p53 gene transfer.
In vivo model of gene therapy
Previously established subcutaneous PC3 tumors in the flanks of nude mice were subjected to in situ gene therapy using AdRGD-PGp53 and Ad-PGp53. As seen in Fig. 7A and Supplementary Fig. S6A, AdRGD-PGp53 conferred superior inhibition of tumor progression compared with either Ad-PGLuc or Ad-PGp53. The experiment was halted on day 92 with the observation that half of the animals in the group treated with AdRGD-PGp53 had not yet reached maximum tolerated tumor volume (1,000 mm3), while all of the animals treated with Ad-PGp53 and Ad-PGLuc had already been euthanized because of tumor burden. Median survival times with sub-maximum tumor volume were 36, 53, and 92 days for the Ad-PGLuc, Ad-PGp53, and Ad-RGDPGp53 groups, respectively, and showed significant statistical difference (Fig. 7B). Immunohistochemical staining of tumor sections with an antibody against pH2AX indicated that Ad-RGDPGp53 increases the DNA damage of the PC3 cells compared with Ad-PGp53 (Fig. 6C and Supplementary Fig. S6B). While the effects of Ad-PGp53 were notable, the impact of AdRGD-PGp53 correlated with long-term survival in 50% of the animals due, in part, to the induction of DNA damage consistent with ROS activation.

In situ gene therapy model. Athymic nude mice were subcutaneously inoculated with 2 × 106 PC3 cells and, once tumors were palpable, treated intra-tumorally with six doses (indicated by arrows) of 1 × 109 transducing units of AdRGD-PGp53 (RGDp53), Ad-PGp53 (p53), or Ad-PGLuc (Luc); n = 6 animals per group. Tumor volumes were measured every 2 days, and mice were euthanized when tumors reached a volume of 1,000 mm3. (
Discussion
The initial reports of treating prostate carcinoma cell lines with an adenoviral vector expressing p53 under control of a constitutive promoter date back some 20 years. 10,12 Even though such a vector was approved for clinical use in China more than 10 years ago, its application has been limited to the treatment of head and neck cancer. 4,5 Transfer of the TP53 cDNA is an obvious choice for cancer gene therapy, though often its effectiveness has not lived up to expectations. The authors have long proposed that improvements to the gene transfer vector may offer renewed hope for this cancer gene therapy approach.
Previously, viral vectors were developed that utilize a p53 responsive promoter, termed PG, which provides levels of transgene expression that are superior to those seen with the CMV promoter or retroviral LTR. 25 –27,32 By inserting the TP53 cDNA in the Ad-PG vector, high-level expression of p53 is supported by a positive feedback mechanism. Here, the Ad-PGp53 vector was used to show that DU145 cells were permissive to transduction and sensitive to the effects of p53. In contrast, PC3 cells were resistant to p53 activity and were not killed, even though they presented high-level expression of p53 upon transduction with Ad-PGp53. In fact, only at a very high MOI could apoptosis be induced in the PC3 cells. Even so, the AdPGp53 vector was shown to be superior, in both DU145 and PC3, to a typical Ad-CMVp53 vector that offers constitutive expression of p53. 28
The RGD-modified vector presented here has resolved a serious issue that was encountered in the authors' previous work. The difficulty in transducing PC3 cells seems to be related to a lack of CAR expression, since an RGD-modified vector, capable of CAR-independent cell entry, efficiently transduced PC3 cells. The next step was to construct an adenoviral vector with the RGD modification, as well as expression of p53 controlled by the positive feedback mechanism. The new adenoviral vector described here, AdRGD-PGp53, was shown to express higher levels of p53 compared with the parental adenoviral vector, Ad-PGp53, even though both utilize the autoregulatory transcriptional mechanism. Having resolved the issues of transduction efficiency, the new AdRGD-PGp53 vector was shown to induce cell death that correlated with the induction of oxidants due, in part, to the impact of high levels of p53 on the expression of p53-target genes in both DU145 and PC3 cell lines.
The interplay of oxidants, DNA damage, and p53 activity are critical for determining cell fate in response to many anti-cancer treatments. Low levels of oxidants can induce DNA damage, cause mutations, and promote tumorigenesis. However, high levels of ROS cause cell death. 15,16 DNA damage induced by oxidants such as the radical •OH, formed by H2O2 and reduced metals, induce DNA damage and can activate p53, which can both inhibit and promote oxidant production. 22 In PC3 cells, it was observed that the AdRGD-PGp53 vector was indeed able to provide p53 expression at a level that conferred superoxide/H2O2 activation and DNA damage. Yet, the Ad-PGp53 vector did not match this result. In the more permissive DU145 cells, both vectors performed essentially the same in inducing ROS and cell death. In this way, the induction of oxidants distinguishes the cellular susceptibility to p53 gene transfer, and only the improved vector was able to achieve this effect in the more resistant PC3 cells.
As seen in the quantitative PCR analyses, key regulators of cell death and oxidant production were impacted by p53 gene transfer. Interestingly, the degree of activation of several genes, including p21Waf1 , NOXA, PIG3, and NOX1, was much higher with AdRGD-PGp53 than it was with Ad-PGp53 in PC3 cells. The more permissive DU145 cells showed similar alteration of target gene expression when either vector was used. Even though high levels of p53 protein were observed in both cell lines when transduced with either vector, only the RGD-modified virus was able to provoke a strong response in PC3 cells.
The induction of NOX1 in PC3 cells by AdRGD-PGp53 was several orders of magnitude greater than that seen with the Ad-PGp53 vector. This finding is striking due not only to the intensity of the response and correlation with oxidant activation, but also to the lack of prior evidence that p53 may control the expression of NOX1, directly or indirectly. This point was not examined experimentally here and will be explored in future studies. However, NOX1 has been shown to contribute to the deacetylation of p53 and, as a result, reduces p53's capacity to transactivate target genes and induce apoptosis, 38 an effect that may be reverted with gentian violet. 39 In endothelial cells, nutrient deprivation was associated with increased NOX2 and correlated with p21Waf1 and p53 activation, but this effect was abrogated in NOX2-deficient cells. 40 In lung cancer cells, wild-type p53 was shown to suppress TGF-β-induced NOX4. Yet, mutant p53 enhanced NOX4 expression. 41
Clear examples of the impact of p53 transcriptional function on ROS have been published. P53 is known to induce the expression of PIG3 (NQO1-quinone oxidoreductase) in response to genotoxic stress, 42 and PIG3 is known to contribute to ROS production. 43 Here, induction of PIG3 upon TP53 gene transfer was also observed. PIG8 (Ei24) induces ROS after activation by etoposide in a p53-dependent manner and induces apoptosis, 44 although alteration in PIG8 mRNA levels in response to the introduction of exogenous p53 was not observed.
P53 can also partner with other proteins to permeabilize the mitochondrial membrane. 45 This study showed the reduction of the mitochondrial membrane potential upon p53 gene transfer. It also showed the increase in NOXA transcript levels concomitant with the reduction in BCL2 mRNA. OKL3846 regulates mitochondrial structure and function, and has been shown to be regulated by p53 in association with peptidylarginine deiminase 4 following DNA damage, 47 although in this study, p53 gene transfer was not associated with alterations in OKL38 transcript levels.
Although the use of catalase and a NOX1-specific inhibitor did not prevent p53 mediated cell death, their presence reduced cell death, in accordance with another report showing that Ad-catalase reduced DU145 cell death. 48 These data indicate that ROS induction is important in prostate carcinoma cell death mediated by adenoviral delivery of the p53 gene and may serve to distinguish cellular sensitivity to p53.
In addition, this study ventured to explore the mRNA expression level of some additional p53 target genes that may have contributed to the cellular response to gene transfer. As expected, the well-known p53 target gene, p21Waf1 , which can promote G1 arrest, 49,50 was activated by p53 gene transfer, though more so with AdRGD-PGp53. In DU145, it was observed that p53 significantly increased the expression of an important gene regulated by DNA damage, Growth arrest and DNA damage (GADD45α), which is known to induce cell cycle arrest and apoptosis. 51 Sestrin2 is involved in mTOR inhibition 52 and p53-mediated autophagy. 53 PC3 only showed significant increase of these mRNAs in the presence of AdRGD-PGp53. However, their levels were even higher in DU145 cells.
In addition, genes involved in cell survival were downregulated by p53 overexpression. Survivin associates with caspases and confers resistance to apoptosis. 54,55 MDM4 interacts with p53 and inhibits its transactivation domain. 56 In DU145 and PC3 cells, the levels of Survivin and MDM4 transcripts were downregulated by both adenoviral vectors. However, as PC3 cells treated with Ad-PGp53 did not show evidence of cell death, reduction of Survivin and MDM4 alone may not be sufficient to induce killing. In all, the evaluation of these p53 target genes suggests that the response to treatment with AdRGD-PGp53 favors cell death, even in the more resistant PC3 cell line.
In a mouse model of in situ gene therapy, it has been observed that intra-tumoral injection with AdRGD-PGp53 was more effective in retarding tumor progression and significantly increased overall survival compared with treatment with Ad-PGp53. Median survival was 92 days for the group treated with AdRGD-PGp53, which is nearly three times longer than the control and almost two times longer than the Ad-PGp53 group. Consistent with the induction of ROS that was observed in vitro, this in vivo assay was associated with the induction of pH2AX, a marker of DNA damage. Although AdRGD-PGp53 did offer significant improvement in tumor inhibition, further refinements, such as alteration of the treatment regime and inclusion of chemotherapeutics, may offer even better control over tumor progression.
In summary, the inclusion of the RGD motif increased adenoviral transduction in PC3 cells, which, in turn, supported higher levels of p53 protein expression and dramatic alterations in p53 target genes, including those involved in apoptosis, cell cycle arrest, autophagy, and ROS. Employing the AdRGD-PG vector for p53 gene transfer was an effective means of overcoming the cellular barriers to treatment, resulting in superior inhibition of tumor growth in vitro and in vivo.
Footnotes
Acknowledgments
We thank Roger Chammas and his staff (ICESP, FM-USP) for ongoing support and critical discussions. We thank Luciana Pescatore Alves (InCor, FM-USP) for expert technical assistance. Financial support was received from the São Paulo Research Foundation, FAPESP (RET, 2011/21256-8; BES, 2013/25167-5).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.