
Abstract
Mucopolysaccharidosis type II (MPS II; Hunter syndrome) is a rare X-linked recessive lysosomal disorder caused by defective iduronate-2-sulfatase (IDS), resulting in accumulation of heparan sulfate and dermatan sulfate glycosaminoglycans (GAGs). Enzyme replacement is the only Food and Drug Administration–approved therapy available for MPS II, but it is expensive and does not improve neurologic outcomes in MPS II patients. This study evaluated the effectiveness of adeno-associated virus (AAV) vector encoding human IDS delivered intracerebroventricularly in a murine model of MPS II. Supraphysiological levels of IDS were observed in the circulation (160-fold higher than wild type) for at least 28 weeks post injection and in most tested peripheral organs (up to 270-fold) at 10 months post injection. In contrast, only low levels of IDS were observed (7–40% of wild type) in all areas of the brain. Sustained IDS expression had a profound effect on normalization of GAG in all tested tissues and on prevention of hepatomegaly. Additionally, sustained IDS expression in the central nervous system (CNS) had a prominent effect in preventing neurocognitive deficit in MPS II mice treated at 2 months of age. This study demonstrates that CNS-directed, AAV9 mediated gene transfer is a potentially effective treatment for Hunter syndrome, as well as other monogenic disorders with neurologic involvement.
Introduction
T
The Sleeping Beauty (SB) transposon system and minicircles are two non-viral gene therapy platforms that have been successfully used in mice for systemic diseases such as MPS types I and VII. 5 –7 Despite being efficient and providing sustained expression in vivo, 6,8 the major drawback of these systems is the inability to penetrate the blood–brain barrier (BBB), 9 which has not yet been resolved. This limits the effectiveness of non-viral gene therapy systems for the central nervous system (CNS).
Various viral vectors have been extensively studied in gene therapy clinical trials for many diseases because of their potency and sustained expression. 10 Among these vehicles, adeno-associated virus (AAV) vectors have been shown to be promising candidates for clinical trials in mediating gene transfer for monogenic disorders. 11 –13 Unlike other AAV serotypes, AAV vector serotype 9 (AAV9) has been demonstrated in many animal models not only to transduce the CNS and peripheral nervous system (PNS) tissues efficiently, but also to penetrate the BBB and transduce various cell types in peripheral tissues. 14 –17 Thus, AAV9 outperforms other viral vectors as a candidate for systemic correction including the CNS for monogenic disorders such as MPS II. Herein we report the effectiveness of CNS-directed, AAV9 mediated human IDS gene transfer to correct IDS deficiency and prevent neurocognitive impairment in a murine model of MPS II.
Materials and Methods
AAV vector assembly and packaging
All vectors were constructed, packaged, and purified at the Penn Vector Core (Philadelphia, PA) and were provided by REGENXBIO, Inc. (Rockville, MD). In brief, the expression cassettes contained a chicken beta-actin (CB7) promoter with cytomegalovirus (CMV) enhancer followed by hIDS or human sulfatase modifying factor 1 (hSUMF1), rabbit beta-actin polyadenylation signal on the backbone of AAV2 inverted terminal repeats (ITR) on both 3′- and 5′-ends. Co-expression constructs included an internal ribosome entry site (IRES) positioned between hIDS and SUMF1 to initiate translation of SUMF1 downstream of the IRES. In this study we investigated five different vector constructs: AAV9 expressing human IDS alone (AAV9.hIDS; Fig. 1A); AAV9 expressing codon-optimized human IDS (AAV9.hIDSco; Fig. 1B); AAV9 co-expressing human IDS and human SUMF1 (AAV9.hIDS-hSUMF1; Fig. 1C); AAV9 co-expressing codon-optimized human IDS and codon-optimized human SUMF1 (AAV9.hIDSco-hSUMF1co; Fig. 1D); and AAV9 expressing human SUMF1 alone (AAV9.hSUMF1; Fig. 1E). AAV vectors were packaged by co-transfecting three plasmids—AAV cis (Fig. 1), AAV trans (pAAV2/9 rep and cap), and adenovirus helper (pAdDF6)—into HEK 293 cells. 18 AAV vector was then purified from supernatants using a Profile II depth filter and concentrated by tangential flow filtration (TFF). The concentrated feed stock was reclarified by iodixanol gradient centrifugation and then re-concentrated using a TFF cassette with a 100 kDa MWCO HyStream screen channel membrane. The purified vector was then tested for purity by SDS-PAGE and for potency by quantitative polymerase chain reaction (qPCR). 18
Adeno-associated virus (AAV) vector constructs for hIDS and hSUMF1 expression. hIDS and hSUMF1 are transcriptionally regulated by the cytomegalovirus (CMV) enhancer/chicken beta-actin promoter (CB7) and by the rabbit beta globin polyadenylation signal (RBG pA), flanked with AAV2-ITRs on both 3′ and 5′ ends. For vectors co-expressing hIDS and hSUMF1, an internal ribosome entry site (IRES) is inserted in between the two open reading frames.
Animal care and husbandry
All animal care and experimental procedures were conducted under approval of the Institutional Animal Care and use Committee (IACUC) of the University of Minnesota. NOD.SCID mice were purchased from The Jackson Laboratory and C57BL/6 wild-type mice were purchased from National Cancer Institute. C57BL/6 iduronate-2-sulphatase knockout (IDS KO) mice were kindly provided by Dr. Joseph Muenzer (University of North Carolina, NC) and maintained under specific pathogen-free conditions at the Research Animal Resources (RAR) facilities of the University of Minnesota. MPS II male pups (IDS−/0) were generated by breeding heterozygous (IDS+/–) females to wild type (IDS+/0) C57BL/6 males. All pups were genotyped by PCR.
AAV vector administration
For intrathecal (IT) injections, eight-week-old mice were injected with a dose of 5.6 × 1010 vector copies (vc) of AAV9 vector between the L5 and L6 vertebrae, as previously described. 19 The injection was performed in conscious animals in a 10–15 s duration. For intravenous (IV) injections, animals were briefly restrained and injected via the tail-vein with a dose of 5.6 × 1010 vc. Intracerebroventricular (ICV) injections were carried out in adult 8-week-old mice, as previously described. 20 Briefly, animals were injected intraperitoneally with a ketamine/xylazine mixture (100 mg/kg ketamine, 10 mg/kg xylazine) to produce deep anesthesia and then mounted in a stereotactic frame (Kopf Model 900). An incision was made to expose the cranium, a small hole was drilled as a site for the injection, and then a Hamilton syringe (Model 701) was used to carry out the infusion at a rate approximately 0.5 μL/min by hand. The syringe was left in place for an additional 3 min and then slowly withdrawn over a period of at least 2 min. The scalp was sutured after completion of the injection, and after recovery from the anesthesia, the mouse was returned to standard housing. All of the mice received a 3-day course of Ketoprofen (2.5–5.0 mg/kg) subcutaneously and Baytril 5 mg/kg intraperitoneally to prevent infection and inflammation post surgery.
Sample collection and preparation
Blood was collected by submandibular puncture using sterile 5 mm lancets (Goldenrode™) into Microvette® heparinized coated tubes (Sarstedt AG & Co.) and centrifuged in an Eppendorf centrifuge 5415D at 7,000 rpm for 10 min. Plasma was collected and stored at −20°C to −80°C for IDS assay. Urine was collected and stored at −20°C until used for creatinine and GAG assay. Organs were harvested by first determining animal weight using an OHAUS® CS 200 scale before euthanasia using a CO2 fume chamber at 2 L/min for 3 min. The animals were perfused with 60 mL of 1 × phosphate-buffered saline (PBS) in a 60 mL syringe (BD) with a SURFLO® winged infusion set (TERUMO®) size 23G × ¾" by hand pressure. The heart, lung, liver, spleen, kidney, and spinal cord were harvested and weighed using a Sartorius BP 61S scale. The brain was micro-dissected into left and right cerebellum, cortex, hippocampus, striatum, olfactory bulb, and thalamus/brainstem. The organs were immediately snap frozen and stored at −70°C until further tissue processing.
For tissue processing, the cerebellum, hippocampus, striatum, and olfactory bulb were added into preassigned 1.5 mL locked-cap microtubes (Eppendorf) containing one scoop (0.2 g/scoop) of 0.5 mm glass beads (Next Advance) in 250 μL of sterile saline solution. The thalamus/brainstem, cortex, and spinal cord were added into assigned locked-cap microtubes containing two scoops of 0.5 mm glass beads in 400 μL of sterile saline solution. Half of the lung and the whole spleen were added into assigned tubes containing two scoops of 0.9–2.0 mm stainless steel blend (0.6 g/scoop) in 400 μL of saline solution. The heart, ∼0.3 g of liver, and one kidney were added into assigned tubes containing three scoops (a mixture of two scoops of 0.9–2.0 mm stainless steel blend [0.6 g/scoop] and one scoop of 3.2 mm stainless steel beads [0.7 g/scoop]) in 600 μL of sterile saline solution. All of the prepared samples in the bead tubes were then homogenized using a Bullet blender® STORM bead mill homogenizer (Next Advance) at speed 12 for 5 min to generate tissue homogenate. Fifty microliters of tissue homogenates were transferred into 1.5 mL microtubes (GeneMate) and stored at −20°C to −80°C for qPCR. The remaining tissue homogenates were clarified using an Eppendorf centrifuge 5424R at 13,000 rpm for 15 min at 4°C. All of the supernatants (tissue lysates) were transferred into new microtubes and stored at −20°C to −80°C until used for IDS, GAG, and protein assays.
Iduronate sulfatase assay
IDS enzyme activity was measured in tissue lysates using 4-methylumbelliferyl-α-L-iduronide-2-sulphate disodium (4-MU-αIdoA-2S; Toronto Research Chemical Incorporation; cat. # M334715) as substrate in a two-step assay. Tissue lysates were mixed with 1.25 mM MU-αIdoA-2S (in 0.1 M sodium acetate buffer pH 5.0 + 10 mM lead acetate +0.02% sodium azide) and incubated at 37°C for 1.5 h. The first-step reaction was terminated with PiCi buffer to stop IDS enzyme activity (0.2 M Na2HPO4/0.1 M citric-acid buffer, pH 4.5 + 0.02% Na-azide). A final concentration of 1 μg/mL Iduronidase (IDUA; R&D Systems; cat. #4119-GH-010) was added into the tubes to start the second-step reaction. The tubes were incubated overnight at 37°C to cleave 4-MU-IdoA into 4-MU. The second-step reaction was terminated by adding 200 μL of stop buffer (0.5 M Na2CO3 + 0.5 M NaHCO3, 0.025% Triton X-100, pH 10.7). The tubes were centrifuged using an Eppendorf centrifuge 5415D at 13,000 rpm for 1 min. Supernatants were transferred into a round-bottom black 96-well plate and fluorescence measured at excitation 365 nm and emission 450 nm, 75 sensitivity using a Synergy MX plate reader and spectrophotometer (Bio Tek) with Gen5 plate reader program. Enzyme activity is expressed in nmol/h/mL plasma for plasma samples and in nmol/h/mg protein for tissue extracts. Protein was determined using the Pierce™ 660 nm Protein Assay Reagent with bovine serum albumin as a standard (cat. # 23208; Thermo Scientific).
Glycosaminoglycan assay
Tissue lysates were incubated overnight with Proteinase K, DNase1, and RNase, as previously described, 21 then, GAG contents were assessed using the Blyscan™ Sulfated Glycosaminoglycan Assay kit (Biocolor Life Science Assays; Accurate Chemical). Blyscan glycosaminoglycan standard 100 μg/mL (cat. # CLRB 1010; Accurate Chemical) was used to make a daily standard curve. Absorbance was measured at 656 nm using a Synergy MX plate reader and spectrophotometer (Bio Tek) with the Gen5 plate reader program. The blank value was subtracted from all readouts. Tissue GAG content is reported in micrograms GAG per milligrams protein, and urine GAG content is reported as micrograms GAG per milligrams creatinine. Urine creatinine was measured using the Creatinine Assay Kit (Sigma–Aldrich®) according to the manufacturer's instructions.
qPCR for IDS vector sequences
Tissue homogenates were mixed with 300 μL cell lysis buffer (5 Prime) and with 100 μg proteinase K, gently rocking overnight at 55°C. DNA was isolated from the sample by phenol/chloroform extraction. Reaction mixtures of 20 μl contained 60 ng of DNA template, 10 μL of FastStart Taqman Probe Master mix (Roche), 200 nM each of forward and reverse primers, and 100 nM of Probe36 (#04687949001; Roche). A C1000 Touch™ Thermo Cycler (Bio-Rad) equipped with CFX manager software v3.1 was used for qPCR reaction. The PCR conditions were: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. IDS primers used were: forward primer—5′-TCCCTTACCTCGACCCTTTT-3′; IDS reverse primer—5′-CACAAGGTCCATGGATTGC-3′. To prepare the standard, pENN.AAV.CB7.hIDS was linearized by digestion with SalI restriction enzyme (New England BioLab, Inc.). The linearized plasmid DNA was then purified using the 5Prime DNA Extraction kit. The plasmid DNA concentration was measured using a NanoDrop 1000 spectrophotometer (Thermo Scientific) with the NanoDrop 1000 3.7.0 program. The purified linearized plasmid DNA was then diluted to prepare the qPCR standard curve. UltraPure™ distilled water (Invitrogen) was used as negative control. A 10-fold dilution series of linearized plasmid was used to generate a standard curve with a range of 1–108 plasmid copies per assay in duplicate with amplification efficiencies between 90% and 110% and R 2 of 0.96–0.98. Vector copy was calculated based on a daily standard curve and expressed as vector copies per cellular genome equivalent (vc/ge).
Neurocognitive testing in the Barnes maze
The Barnes maze 22 is a circular platform measuring approximately 4 feet in diameter and is elevated approximately 4 feet from the floor with 40 holes spaced equally around the perimeter. All of the holes are blocked except for only one hole that is open for the mouse to escape the platform. Different visual cues were attached to each of the four walls for the mouse to use as spatial navigators. At 6 months of age, test mice were placed in the middle of the platform with an opaque funnel covering the mouse. The cover was lifted, releasing the mouse and exposing it to bright light. The animal is expected to complete the task by escaping the platform using the one open hole within 3 min. Each mouse was subjected to four trials per day for a total of 6 days. The time that the mouse required to escape the platform in each trial was recorded, and the average was calculated for each day in each group.
Statistical analysis
Data are reported as mean ± standard error (SE). Statistical analyses were performed using Prism 6. Two-way analysis of variance with Tukey's post test was used to evaluate the significance of differences among test groups for IDS assay, GAG assay, and neurobehavioral assay, with a p-value of <0.05 considered significant. A two-tailed t-test on Microsoft Excel was used to evaluate differences in IDS activities between the left and the right hemispheres of the microdissected brain.
Results
Pilot studies: comparison of vector constructs and route of administration to achieve IDS expression in the CNS
A pilot study was conducted to compare several AAV vector constructs and to find a suitable route of administration resulting in IDS expression in the CNS. NOD.SCID mice were used for this study to circumvent the potential complication of an anti-IDS immune response. The four vectors shown in Fig. 1A–D (AAV9.hIDS, AAV9.hIDSco, AAV9.hIDS-hSUMF1, and AAV9.hIDSco-hSUMF1co) were delivered by IT administration, as described in the Materials and Methods. SUMF1 encodes an enzyme which post-translationally oxidizes an active site cysteine in lysosomal sulfatases, including IDS, converting the enzyme into a catalytically active form. 23 The addition of SUMF1 to some of the vectors was to determine if SUMF1 activity is rate-limiting in producing active IDS protein when IDS is overexpressed. Five untreated IDS+ NOD.SCID mice were used as a control group. Six weeks post injection, the mice were euthanized, and the brain was microdissected into different portions. Our parallel studies in MPS I have demonstrated supraphysiological activities of IDUA in the CNS post-IT injection of AAV9 vector encoding hIDUA. 24 We thus expected to see high levels of IDS, exceeding the endogenous level in IDS+ NOD.SCID mice administered AAV9.hIDS vector. Surprisingly, we observed no significant increase of IDS activity in the CNS exceeding the endogenous level of uninjected NOD.SCID mice, regardless of vector construct (data not shown). AAV9.hIDS was also injected into two groups of three wild type C57BL/6 (IDS+ C57BL/6) mice at 8 weeks of age, one group via IT administration and the other group via IV administration. Again, no significant increase in the level of IDS activity in the CNS above the endogenous level of untreated controls was observed (data not shown). Thus, neither IT nor IV injection of IDS-encoding AAV vector appeared to be a suitable route of administration.
Another unexpected result from the initial pilot study described above is that while there was undetectable increase in IDS activity in the CNS, plasma IDS activity in both IV- and IT-treated groups was increased up to approximately 140-fold above the untreated wild-type level and persisted for at least 12 weeks post treatment (Fig. 2A). This result suggests that AAV vector was distributed to the peripheral circulation after IT injection into the cerebrospinal fluid (CSF). The presence of sustained enzyme activity for at least 12 weeks post injection (either IT or IV) also suggests that hIDS is presumably non-immunogenic for C57BL/6 mice.
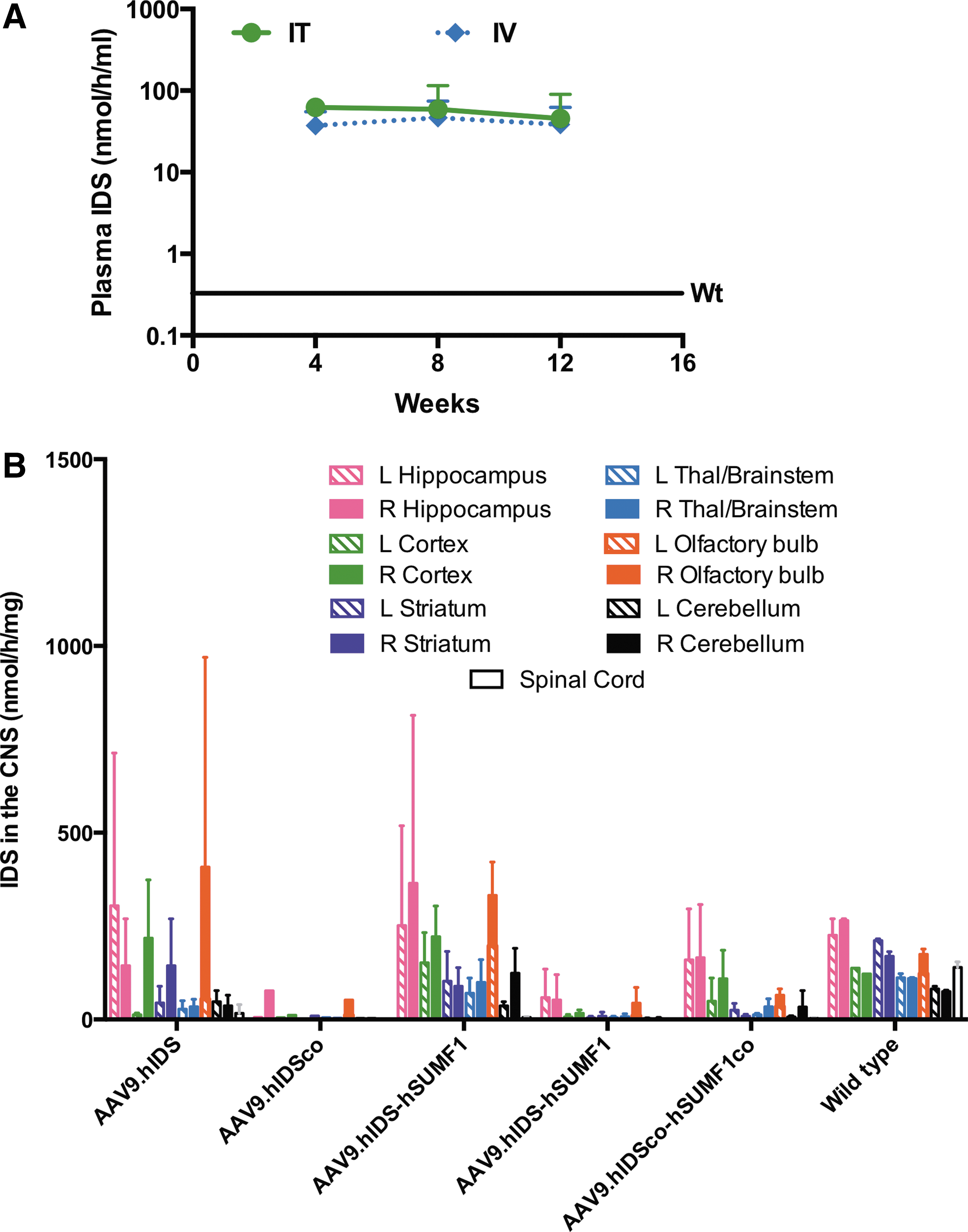
Iduronate-2-sulfatase (IDS) expression after intrathecal (IT), intravenous (IV), or intracerebroventricular (ICV) administration of AAV9 IDS vectors.
We then administered the same four vector constructs (AAV9.hIDS, AAV9.hIDSco, AAV9.hIDS-hSUMF1, and AAV9.hIDSco-hSUMF1co) into immunocompetent MPS II mice by ICV injection—a procedure that supports a much higher level of transduction in the CNS than IT injection. 21 Immunosuppression of the MPS II test animals was not necessary, since we found that expression of human IDS does not elicit an immune response in C57BL/6 mice. An additional group of MPS II mice was injected ICV with a combination of two vectors—AAV9.hIDS (Fig. 1A) and AAV9.hSUMF1 (Fig. 1E)—at a 1:1 ratio (AAV9.hIDS + AAV9.hSUMF1; at a dose of 5 × 1010 vc total) to determine if there would be additional IDS activity when SUMF1 and IDS are both translated independently rather than relying on translation of SUMF1 from a downstream position by employing an IRES. Untreated wild-type littermates were used as controls. Six weeks post injection, the animals were euthanized, organs were harvested, and brains were microdissected to determine IDS activity. Animals injected with AAV9.hIDS, AAV9.hIDS-hSUMF1, or AAV9.hIDS + AAV9.hSUMF1 showed levels of IDS activity approximately 10–40% of the wild-type level in most portions of the brain (Fig. 3). IDS activity was undetectable in all areas of the brain in MPS II mice (Fig. 3C). Animals injected with codon-optimized vector constructs showed mostly <10% of the wild-type level, so codon optimization of the IDS sequence did not result in a higher level of IDS activity in transduced tissues. There was no significant difference between AAV9.hIDS-injected animals and animals injected with AAV9.hIDS plus hSUMF1. Thus, in our hands co-delivery of hSUMF1 either on the same vector or on a separate vector did not enhance the level of IDS activity assessed. Vector AAV9.hIDS (Fig. 1A) was subsequently used for more extensive efficacy studies in ICV-administered MPS II mice, as described below, as neither the addition of SUMF1 nor the codon-optimization algorithm resulted in increased IDS activity compared to the native hIDS cDNA sequence.
IDS expression and metabolic correction after ICV injection of AAV9.hIDS in MPS II mice.
Prevention of CNS and peripheral lysosomal disease by ICV administration of AAV9.hIDS vector
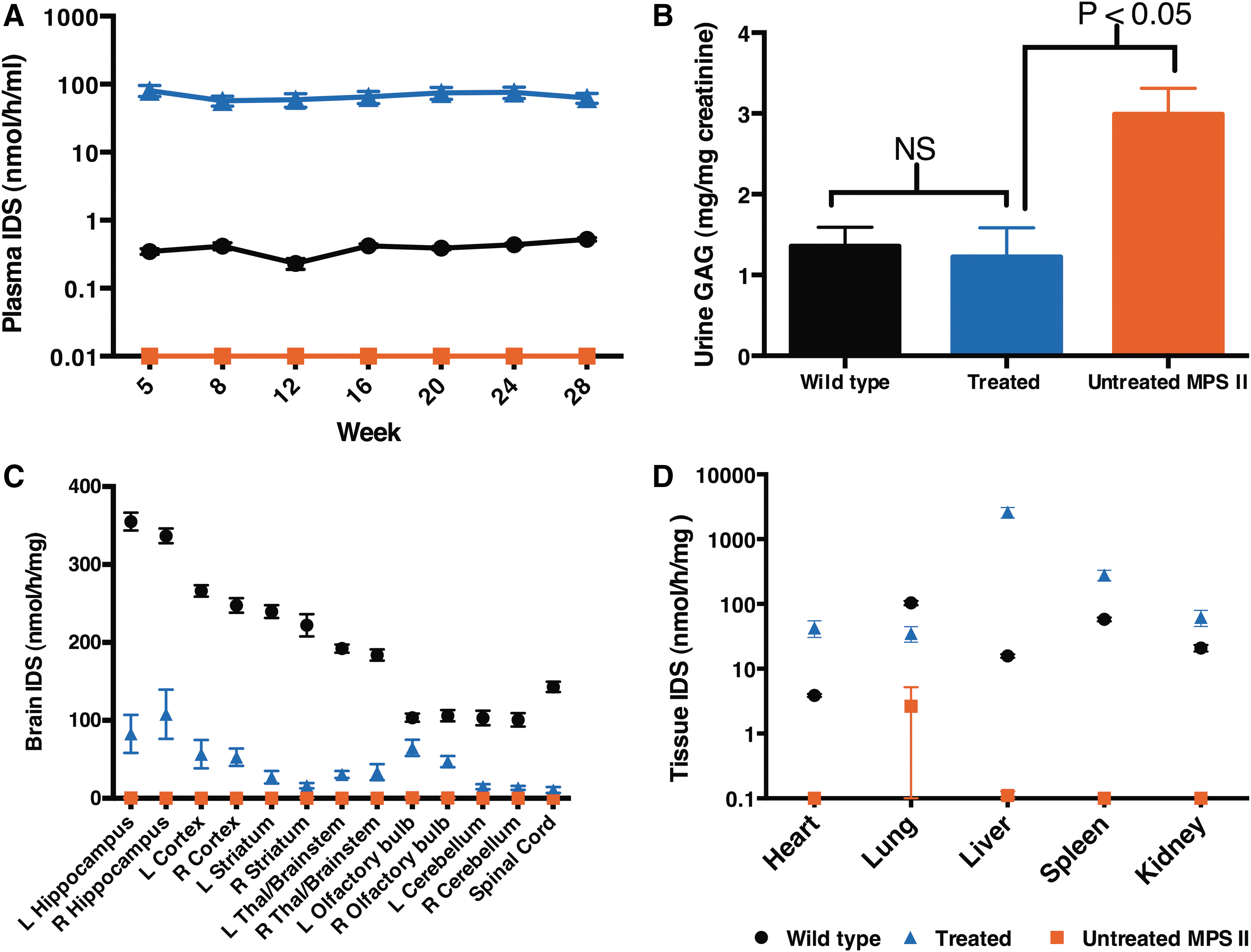
A dose of 5.6 × 1010 AAV9.hIDS vc was infused into 8-week-old MPS II mice by ICV injection to achieve widespread CNS distribution of the vector through the CSF. As in the pilot study, plasma IDS activities up to 160-fold higher than wild type were observed in this larger cohort of ICV-treated MPS II animals, and this expression persisted throughout the experiment (28 weeks post injection; Fig. 3A). Urine was collected at the end of the study (week 40 post injection) to evaluate the effect of long-term IDS expression on GAG excretion in the treated animals compared to wild-type and untreated MPS II mice. Urine GAG was significantly elevated in MPS II animals when compared to wild-type littermates (Fig. 3B; p < 0.05). The treated animals demonstrated a significant reduction in urine GAG content (p < 0.05) when compared to untreated littermates and were normalized when compared to the wild-type level (p > 0.05).
At 10 months of age (40 weeks post injection), all mice were euthanized, and organs were harvested for analysis. IDS activity was undetectable in all areas of the brain and spinal cord of untreated MPS II mice (Fig. 3C). AAV9.hIDS-injected animals had IDS activity in all regions of the brain at approximately 9–28% of wild type, 53% in the olfactory bulb and 7% in the spinal cord (Fig. 3C). Although the vector was infused into the right ventricle of the brain, no significant difference in IDS activity was observed between the left and right hemispheres (p > 0.05). Unlike the CNS, supraphysiological levels of enzyme activity were observed in all tested peripheral organs such as the heart, liver, spleen, and kidney (11-, 166-, 5-, and 3-fold, respectively; Fig. 3D), except in the lung where 34% of wild type was observed (Fig. 3D). This suggests that the vector was able to cross the BBB from the CNS into the circulation whereby it was taken up by and expressed in peripheral organs.
DNA was isolated from the same tissue homogenates and evaluated for vector distribution by qPCR. Consistent with Wolf et al., 21 ICV infusion of AAV vector resulted in global distribution of vector in the CNS (Fig. 4A) and in all of the tested peripheral tissues (Fig. 4B). The highest vc number was observed in the right hippocampus (49 vc/ge), while similar vc were observed between the left and right hemispheres for all regions of the brain (Fig. 4A). Unlike the CNS, relatively low copy numbers were detected in most tested peripheral tissues, including the heart, lung, spleen, and kidney of the treated animals (<0.6 vc/ge; Fig. 4B), while high vc numbers were observed in the liver (44 vc/ge; Fig. 4B). This suggests that enzyme produced by the liver was released into the circulation where the tested peripheral organs took up the circulating enzyme (i.e., metabolic cross-correction).
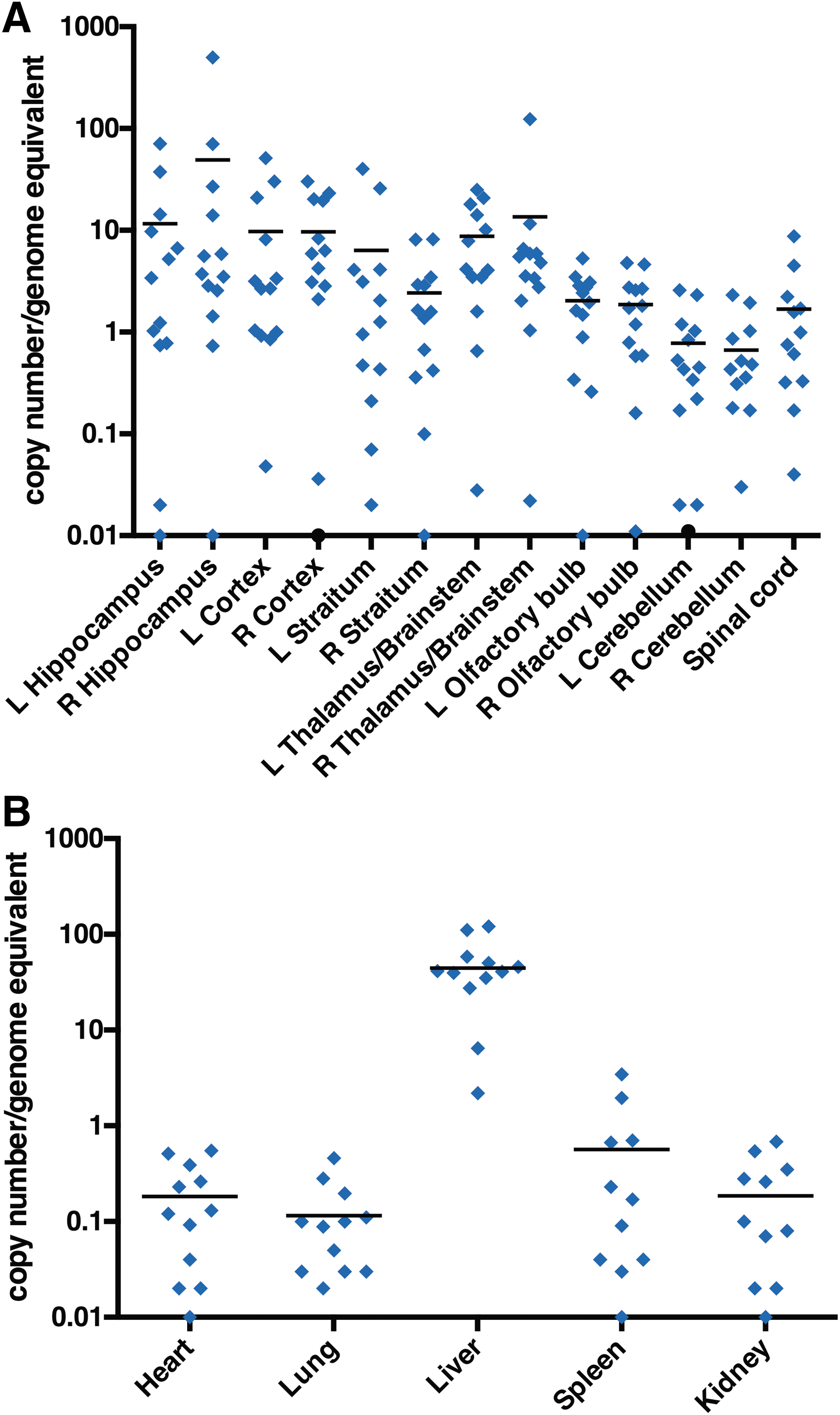
AAV9.hIDS vector biodistribution after ICV injection in MPS II mice.
The global distribution and expression of IDS had a significant effect on accumulation of lysosomal storage materials. Elevation of lysosomal GAG content was observed in the CNS of untreated MPS II mice when compared to wild-type littermates (Fig. 5A; p < 0.0001). Even though there was only 10–40% of wild-type IDS level in the CNS, GAG content in the CNS was normalized when compared to wild type (p < 0.01; Fig. 5A). Similar to the CNS, significant elevation of lysosomal GAG content was observed in all tested peripheral organs of the untreated MPS II animals when compared to wild type (p < 0.01; Fig. 5B). In contrast, significantly decreased GAG levels were observed in all tested peripheral tissues (Fig. 5B) of the treated mice when compared to the untreated group (p < 0.01). Statistical analysis showed no significant difference in GAG content between wild-type animals and the treated group (p > 0.05), which indicates that GAG content was normalized in the treated group.
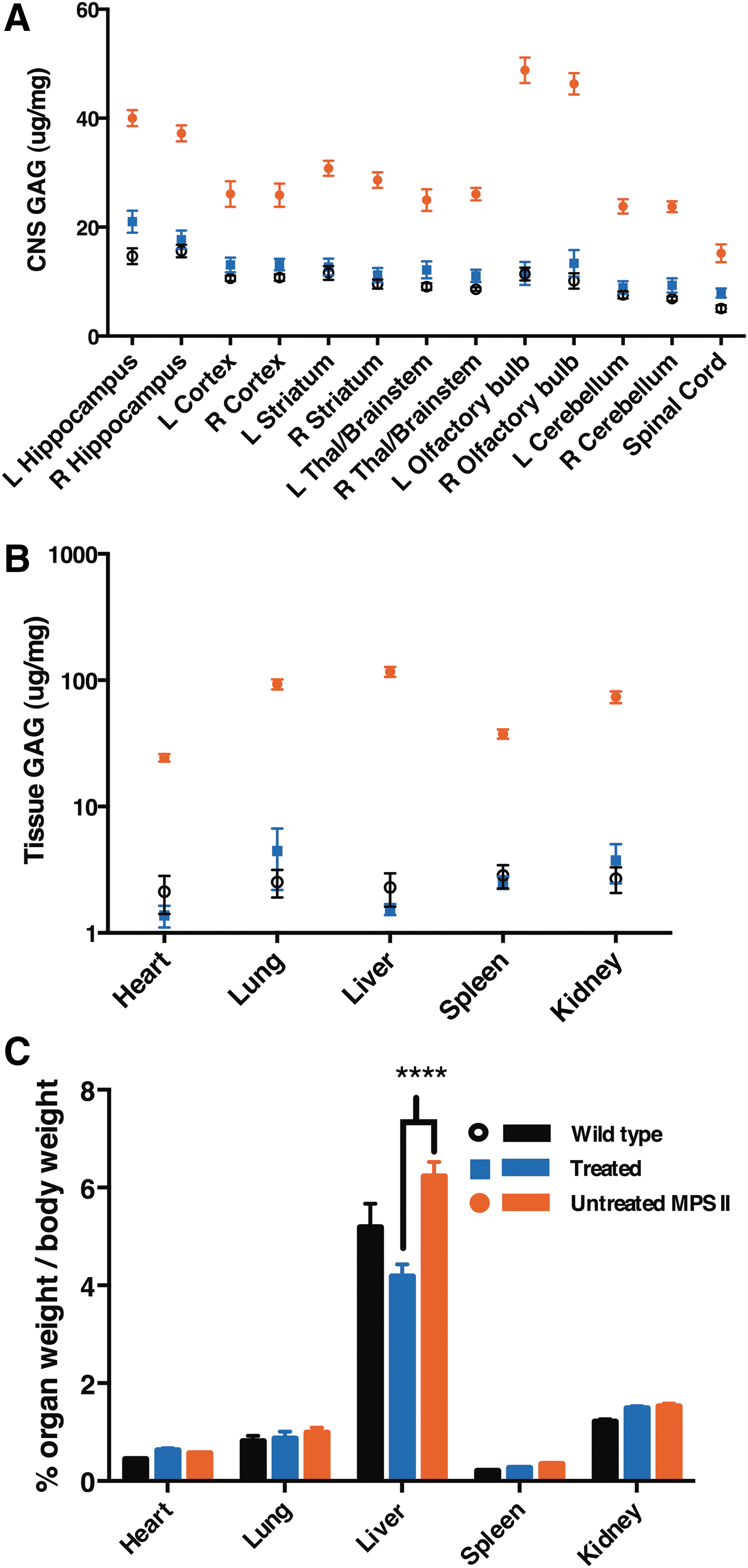
Correction of storage disease in AAV9.hIDS-treated MPS II mice.
The body weights of all mice were measured before sacrifice, and organs were weighed after the animals were perfused with 1 × PBS, calculating the percentage of organ weight to body weight immediately post sacrifice. No significant difference was observed in the size of the heart, lung, spleen, or kidney among all groups. However, the liver of untreated MPS II mice was 20% larger than that of wild-type animals (6.2% and 5.2% of total body weight, respectively; p < 0.001; Fig. 5C). In contrast, the liver of the treated MPS II mice was 68% smaller than the untreated group (4.2% and 6.2% of total body weight, respectively; p < 0.0001; Fig. 5C). This result shows that normalization of GAG content in the liver in turn prevented hepatomegaly in the treated mice.
Sustained expression of IDS in the CNS leads to prevention of neurocognitive deficit in MPS II mice
At 6 months of age (4 months post treatment), untreated MPS II mice, AAV9.hIDS-treated MPS II mice, and control normal littermates were evaluated for neurocognitive function in the Barnes maze, a test for spatial navigation and memory. The animals were subjected to a series of 3-min trials, four trials per day for a course of 6 days. Wild-type littermates showed reduced latency to escape (30 s) on day 6, while untreated MPS II mice exhibited a significant deficit in learning this task (latency to escape reduced only to 71 s; p ≤ 0.05 vs. normal littermates; Fig. 6). In contrast, the AAV9.hIDS-treated mice showed a marked reduction in latency to escape (25 s), significantly outperforming the untreated MPS II mice (p ≤ 0.01) on days 5 and 6. In addition, there was no significant difference between the treated animals and wild-type littermates (p > 0.05; Fig. 6). We conclude that sustained expression of IDS in the CNS prevented the emergence of neurocognitive deficits in MPS II mice when treated at a young age.
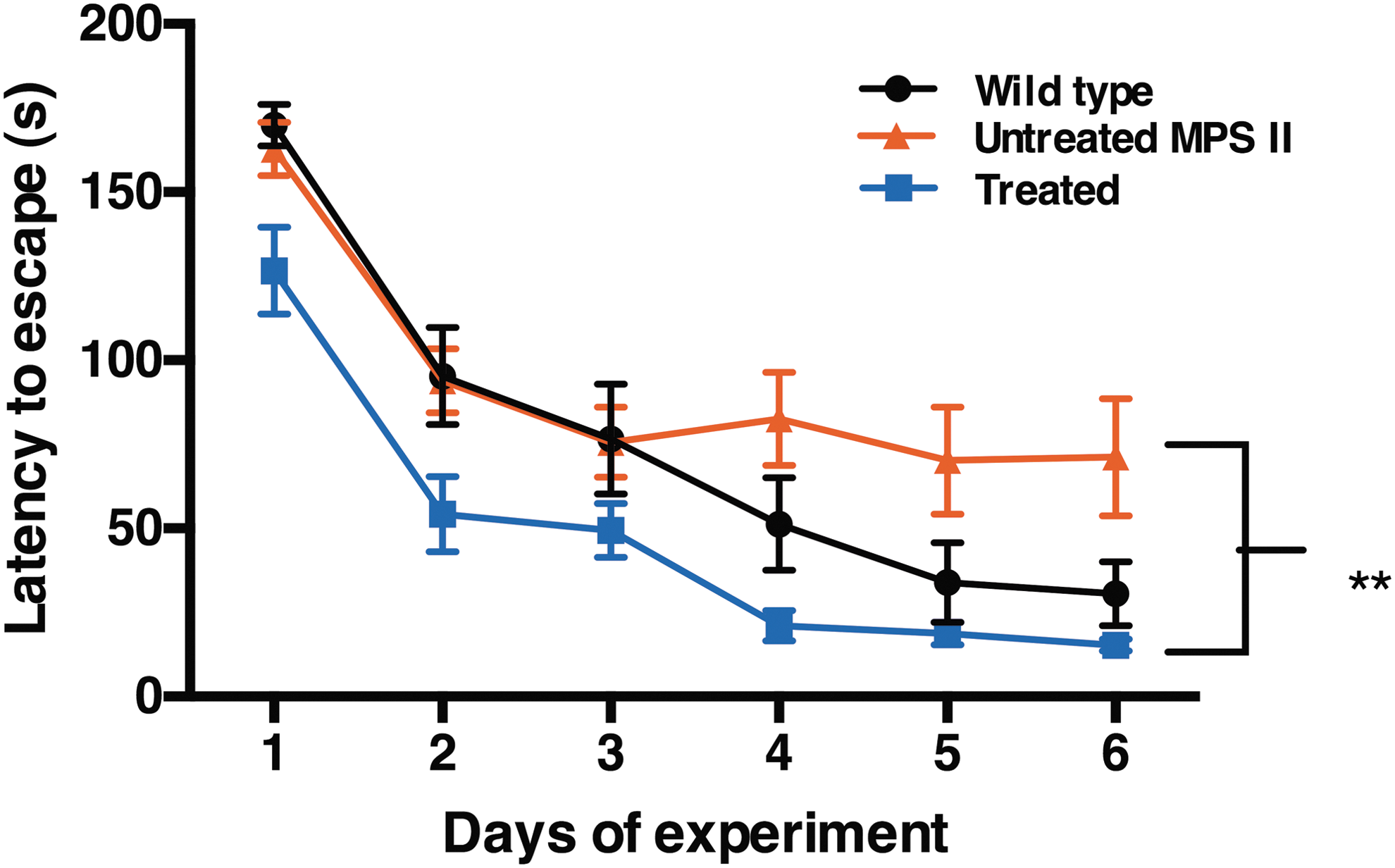
Neurocognitive function assessed in the Barnes maze. Animals in all three groups were tested in the Barnes Maze, as described in the Materials and Methods. The graph depicts the average latency to escape (seconds) that each group of mice required during four trials conducted on each day for six consecutive days. The average latency to escape required for wild-type and treated groups decreased over the course of 6 days of the experiment. In contrast, there was no improvement observed for the MPS II mice from day 3 to day 6. No significant difference in the performance of treated mice versus wild-type littermates was observed, while the treated mice significantly outperformed the untreated MPS II mice on days 5 and 6 (p ≤ 0.01).
Discussion
In this study, we administered AAV9.hIDS using a strong promoter to the CNS of MPS II mice. We found that levels of IDS activity in the CNS were only 7–28% of wild type. In contrast, levels of IDS activity in the circulation and in tested peripheral organs were at least 2- and up to 170-fold higher than wild-type levels. We also observed that sustained IDS expression leads to global normalization of GAG content. Finally, we observed that sustained levels of IDS activity had a profound effect on preventing neurologic deterioration.
No significant increase in IDS activity in the CNS above the endogenous level was observed in IV- or IT-treated IDS+ mice, regardless of vector construct, route of administration, or mouse strain, even though IDS was expressed under regulation of the strong CB7 promoter (data not shown). However, the level of plasma IDS activity in IT-treated IDS+ mice was an average of 100-fold higher than the wild-type level, indicating successful IT administration of vector. Similar high levels of plasma IDS were observed when AAV9.hIDS was infused into MPS II mice via ICV injection (Figs. 2B and 3C), although in this case IDS activity in the CNS above baseline was observed in all 12 assayed portions of the brain (Fig. 3C). Similar IDS activities were reported from Motas et al. (in coronal sections of the brain) 25 and Hinderer et al. (whole brain), 26 in which they observed approximately 20–40% of wild-type levels in the CNS. Here, a more extensive analysis of vector distribution and expression is reported in all different micro-dissected areas of the brain after ICV administration of AAV9.hIDS in MPS II mice.
Unlike Motas et al. 25 and Hinderer et al., 26 we emphasize the limited level of IDS activity achieved in the CNS after ICV administration of AAV9.hIDS. These limited levels of IDS activity are in stark contrast with the levels of IDUA activity observed in the CNS of MPS I mice after ICV injection of AAV9-IDUA vector, in which 100- to 1,000-fold higher than wild type levels of IDUA activity are observed. 24 Supraphysiological levels of IDS (>1,000 nmol/h/mg) were also observed in the liver of ICV-administered animals in our study at a similar vc number that yielded 10- to 100-fold less IDS expression (10–100 nmol/h/mg) in the CNS. Highly relevant to the goal of CNS-directed gene therapy for MPS II, therefore, is this question: what is it that limits expression of IDS in the brain after highly efficient AAV-mediated IDS gene delivery?
One possibility is that SUMF1 activity might be rate limiting for the generation of active IDS in the brain. SUMF1 is required for the post-translational activation of lysosomal sulfatases, including IDS. 23 hIDS enzyme clearly was activated in tissues of the MPS II mouse, presumably by mouse SUMF1, but this process could be rate limiting in the brain if AAV-encoded hIDS protein is expressed in a large quantity but only a limited amount of hIDS becomes activated. For example, Fraldi et al. 27 demonstrated that N-sulfoglucosamine sulfohydrolase activities were increased when the enzyme was co-expressed with SUMF1 in their MPS IIIA studies. Anticipating a potential limitation of SUMF1, hIDS and hSUMF1 were co-transduced either on the same construct (Fig. 1C) or on two separate vectors (Fig. 1A and E), but no significant increase in hIDS activity was found compared to delivery of hIDS alone (Fig. 2B). We concluded that hIDS activity was not enhanced by co-delivery with hSUMF1 in these experiments. Further studies evaluating SUMF1 mediated activity of IDS in the CNS are nonetheless warranted.
Another possibility is that the CB7 promoter might be limiting when compared to the endogenous IDS promoter. To further investigate this possibility, IDS activity (nmol/h/mg protein = unit) per vc was calculated for each tissue and compared to IDS activity per endogenous copy in male mice. IDS activities in the CNS of the treated mice were observed between 2 and 32 (mean of 8) units per vc compared to wild-type male mice, which express an average of 200 units per genome equivalent (approximately only 1–31% of wild-type level). By comparison, in the liver there was an average of 55 units of IDS activity per vc in the ICV-treated mice. From this, one might conclude that the CB7 promoter is not as robust as the endogenous IDS promoter in the brain. However, the lower level of vector-mediated IDS expression per vc versus endogenous expression most likely results from the presence of excess transcriptionally inactive AAV genomes remaining in the intracranial space post injection, possibly due to inefficient intracellularization. Nonetheless, the MPS II results contrast greatly with the results from our MPS I studies in which levels of IDUA activity at approximately 10- to 100-fold higher than wild type were observed in the CNS of mice administered AAV-hIDUA intrathecally, 24 while heterozygous MPS I animals express approximately 6 units per genome equivalent in the brain. 28 Therefore, it is feasible to achieve or exceed wild-type IDUA levels in the CNS of MPS I mice. The promoter that we used in the MPS II study was similar but not identical to the promoter used in the MPS I studies, 21,24 and the possibility that relative promoter strength may have contributed to the observed differences in outcome cannot be excluded.
Surprisingly, we observed supranormal levels of enzyme activity in all tested peripheral organs of the treated animals. The highest levels of IDS activity (164-fold wild type) were observed in the liver, which correlated with the highest number of vc found (48 vc/ge). In contrast, the tested peripheral organs (other than the liver) contained low levels of vector, but levels were higher than wild-type levels of IDS activity. The exceptionally high levels of IDS in the liver (Fig. 3D) lead to sustained high levels of IDS activity (up to 172-fold higher than wild type) in the circulation (Figs. 2A and 3A). Similar results have been reported in a MPS IIIA study and several MPS I studies using different types of vectors, 5,7,29 along with two studies in MPS II mice. 25,26 These results suggest that in our study, the liver acts as an enzyme factory that produces an enormous amount of IDS and releases it into the circulatory system where it is subsequently taken up by the other organs in the periphery. Even though higher than wild-type levels of IDS enzyme activity were observed in the heart, spleen, and kidney in the treated mice, the vector levels in these organs were unexpectedly low (<0.6 vc/ge). This low transduction rate suggests that these organs took up circulating IDS, leading to metabolic cross-correction with higher levels of IDS than wild type.
Polito et al. observed no detectable IDS activity in the CNS after a single IV injection of AAV2/5 CMV-hIDS. However, they did observe partial correction of GAG content in the CNS. 30 They speculated that only a minute fraction of high-level circulating hIDS produced from the liver penetrated the BBB into the CNS, with subsequent partial correction of GAGs. Similarly, in the current experiments we found exceptionally high levels of IDS in the circulation of AAV9.hIDS-treated mice, and yet only subphysiological activities of enzyme were observed in the CNS. This finding indicates that an insignificant amount of enzyme produced from the liver after ICV injection of vector was able to penetrate the BBB from the circulation into the CNS. This is consistent with the lack of increased IDS over wild-type levels in the CNS after IV administration of AAV9.hIDS in IDS+ C57BL/6 mice (Fig. 2A). Therefore, the level of IDS observed in the brain most likely relied on transduction of the vector construct inside the CNS after ICV injection rather than from circulating enzyme. Further investigation is needed to achieve IDS levels comparable to or higher than the wild type in the CNS.
Sustained levels of enzyme expression after direct vector injection into the CNS have been shown to have profound effects on GAG reduction in several MPS studies irrespective of whether wild-type levels of those enzymes were achieved. 20,22,24,25,28 Similarly, although only subphysiological levels of IDS were observed in the CNS after ICV injection of AAV9.hIDS, there was a significant effect of the enzyme on GAG reduction that was greater than expected. Desnick et al. and Polito et al. speculated that only <5% of the wild-type level of lysosomal enzyme is needed to correct a GAG storage defect. 30,31 The present results are consistent with this speculation. After AAV9.hIDS treatment, all areas of the brain (Fig. 5A) and even the spinal cord (the organ with the lowest enzyme measured; 10.7 nmol/h/mg protein; 7.4% of wild type; Fig. 3C) showed significantly lower GAG content than in untreated MPS II mice (Fig. 5A). Statistical analysis revealed striking results, as GAG content in the CNS of the treated mice was normalized when compared to wild type. We also observed normalization of GAG content in the urine and in the heart, lung, liver, spleen, and kidney of the treated animals. These results indicate a sustained effect of IDS expression on correction of GAG content.
Roberts et al. demonstrated a direct relationship between GAG accumulation and liver size when injecting rodamine B, a GAG synthesis inhibitor, into MPS IIIA mice. They found that GAG content was decreased in the liver, leading to normalization of liver size. 32 Motas et al. observed a preventive effect on hepatomegaly after ICV administration of AAV9.hIDS into MPS II mice. 25 Similarly, we also observed that the weight of the liver in treated mice was normalized after ICV injection of AAV9.hIDS (Fig. 5C). This result indicates a profound effect of sustained IDS expression leading to normalization of GAG content in the liver, which in turn prevents hepatomegaly in the treated mice.
Several MPS studies have demonstrated prevention of neurologic deficits after AAV-mediated gene transfer in mice. 21,25,33 We observed neurocognitive deficiency in untreated MPS II mice at 6 months of age. We also observed that sustained IDS expression in the CNS prevented the emergence of neurocognitive dysfunction in MPS II mice after ICV infusion of AAV9.hIDS (Fig. 6). Several studies have shown that the hippocampus is associated with neurocognition in rodents. 34 –36 However, the possibility that physical impairment such as vision, olfactory sense, or motor neuron defects could also affect the results in the Barnes maze cannot be excluded, as rodents require all of the aforementioned physical capabilities in order to perform the required task in this test. 37 Additional behavioral analyses would support the observation that neurocognitive deficit plays a pivotal role in learning impairment of untreated MPS II mice and its prevention by AAV-mediated IDS gene transfer.
In conclusion, this paper characterizes the benefit of direct AAV9-mediated hIDS gene transfer to the CNS. The most important challenge emerging from this study is the limited level of AAV-mediated IDS expression achieved in the CNS in comparison with other tissues such as the liver, and in comparison with the expression of other therapeutic genes introduced into the CNS by AAV-mediated transduction such as IDUA. 21,24 Nonetheless, the MPS II data indicate that direct injection of AAV9.hIDS vector into the CNS resulted in efficient gene transfer that is key to treatment of MPS II and prevention of neurocognitive deficits. We also found that AAV9.hIDS vector was capable of crossing the BBB from the CNS into the circulation, resulting in global transduction of the vector outside of the CNS, providing long-term expression of IDS enzyme systemically. Even though the IDS activities in the brain were lower than expected, this study nonetheless supports the notion from previous studies 30,31 that <10% of the wild-type level of IDS is needed to prevent GAG storage accumulation. In addition, sustained IDS expression corrected the accumulation of GAG in liver and subsequently prevented the emergence of hepatomegaly. Finally, our results reinforce the importance of sustained IDS expression in the CNS in preventing the emergence of neurologic deficits when animals are treated at a young age. We anticipate that this study that this study will contribute to the field in developing a long-term effective treatment with neurological benefits for MPS II patients.
Footnotes
Acknowledgments
The authors thank Dr. Joseph Muenzer for providing the IDS KO strain, and the University of Minnesota NINDS Behavioral Phenotyping Core.
Author Disclosure
The work described in this manuscript was supported by REGENXBIO. Karen Kozarsky is a recent employee of REGENXBIO, and currently consults for and receives compensation from REGENXBIO.