
Abstract
Investigations based on mesenchymal stem cells (MSCs) for osteoporosis have attracted attention recently. MSCs can be derived from various tissues, such as bone marrow, adipose, umbilical cord, placenta, and dental pulp. Among these, dental pulp–derived MSCs (DPSCs) and hepatocyte growth factor (HGF)-modified DPSCs (DPSCs-HGF) highly express osteogenic-related genes and have stronger osteogenic differentiation capacities. DPSCs have more benefits in treating osteoporosis. The purpose of this study was to investigate the roles of HGF gene-modified DPSCs in bone regeneration using a mouse model of ovariectomy (OVX)-induced bone loss. The HGF and luciferase genes were transferred into human DPSCs using recombinant adenovirus. These transduced cells were assayed for distribution or bone regeneration assay by transplantation into an OVX-induced osteoporosis model. By using bioluminogenic imaging, it was determined that some DPSCs could survive for >1 month in vivo. The DPSCs were mainly distributed to the lung in the early stage and to the liver in the late stage of OVX osteoporosis after administration, but they were scarcely distributed to the bone. The homing efficiency of DPSCs is higher when administrated in the early stage of a mouse OVX model. Micro-computed tomography indicated that DPSCs-Null or DPSCs-HGF transplantation significantly reduces OVX-induced bone loss in the trabecular bone of the distal femur metaphysis, and DPSCs-HGF show a stronger capacity to reduce bone loss. The data suggest that systemic infusion of DPSCs-HGF is a potential therapeutic approach for OVX-induced bone loss, which might be mediated by paracrine mechanisms.
Introduction
O
It is clear that pluripotent stem cells have an abundant potential in regenerative medicine, and mesenchymal stem cells (MSCs) are numerically the most favored cell type presently undergoing clinical trials. 14 MSCs can be easily obtained from various sources, such as bone marrow, adipose tissue (AD), the umbilical cord (UC), the placenta, the synovium, dental pulp, and so on. MSCs could be induced to differentiate into osteoblasts under the well-recognized conditions of osteogenic induction and could secret various bioactivators to regulate bone formation. 15 –17 Additionally, MSCs possess immunosuppressive properties and are deemed safe for clinical applications approved by the Food and Drug Administration. 18 These properties make MSCs ideal candidates in the search for a cure for osteoporosis. It has been proved that local and systemic injections of autologous or xenogenic MSCs have a beneficial effect in both rodent and goat osteoporosis models. 19 –22 A previous study revealed that dental pulp–derived MSCs (DPSCs) possess low immunogenic potential and have a strong osteogenic differentiation capacity. In a swine periodontitis model, the therapeutic effects of DPSC, periodontal ligament stem cells (PDLSC), UC-MSC, and AD-MSC on periodontal regeneration were compared. The radiological and quantitative assessment data show that DPSCs are the most effective stem cells for bone regeneration. Although DPSCs and BM-MSC both had stronger osteogenic differentiation capacity, DPSCs may have a better prospect for osteoporosis because of their stronger proliferation ability and because they are easy to obtain.
Hepatocyte growth factor (HGF) is a multifunctional cytokine that promotes cell survival and regeneration of tissues, suppresses and improves chronic inflammation and fibrosis, stimulates osteoblast proliferation, and participates in bone remodeling. 23,24 A previous study also showed that HGF gene transfer could significantly enhance the regeneration of periodontal bone. 25 The combined application of HGF gene therapy and DPSCs could improve periodontal tissue regeneration in a swine model.
The present study cultured human DPSCs (hDPSCs) and modified them by introducing an adenovirus vector carrying the HGF gene (Ad-HGF) or empty control (Ad-Null). Then, gene-modified cells (DPSCs-HGF or DPSCs-Null) were administrated into ovariectomy (OVX)-induced bone-loss mice through the tail vein. Using this approach, the roles of HGF and DPSCs in bone regeneration were evaluated. The results showed that DPSCs modified with HGF could significantly postpone trabecular bone loss in the early stage of OVX osteoporosis. Thus, a novel strategy is provided that transplantation of DPSCs-HGF can potentially prevent the development of osteoporosis.
Materials and Methods
Isolation and culture of DPSCs
Normal human impacted third molars were collected from adults (19–29 years old) at the Dental Clinic of Beijing Stomatological Hospital under approved guidelines set by the Research Ethics Committee of Capital Medical University, China (BSH [2015] D-15). All patients provided written informed consent to participate. Tooth surfaces were cleaned and cut around the cementoenamel junction with sterilized dental fissure burst to reveal the pulp chamber. The pulp tissue was gently separated from the crown and root, cut into 1 mm3 pieces, and then digested in collagenase and dispase (Sigma–Aldrich, St. Louis, MO) for 40 min at 37°C. Tissue pieces were then cultured in a cell culture flask (Corning, Corning, NY) with alpha-modified Eagle's medium (α-MEM; Gibco®; Thermo Fisher Scientific, Grand Island, NY) supplemented with 15% fetal bovine serum (FBS; Thermo Fisher Scientific) at 37°C in 5% carbon dioxide. Cells were harvested on day 14. All cells in this study were used after three or four passages. MSC characterizations, including surface molecule expression profiles and multilineage differentiation, were performed as described previously
26
(data shown in Supplementary Figs. S1 and S2; Supplementary Data are available online at
Adenovirus vectors
The adenovirus vectors (AdVs) used in the study were Ad-HGF, a replication-defective adenovirus expressing human HGF; Ad-Luc, a replication-defective adenovirus expressing luciferase; and Ad-Null, a replication-defective adenovirus not carrying exogenous genes. Viruses were created as reported previously. 27
DPSCs were infected at a multiplicity of infection (MOI) of 150 with Ad-HGF or Ad-Null, and cells were collected 2 days post infection for use as osteoporosis therapy. Ad-Luc-infected DPSCs (DPSCs-Luc) were used to detect the distribution and engraftment of DPSCs of OVX mice in vivo.
Animals
The OVX procedure was performed on 9-week-old female C57BL/6J mice (N = 47), as described previously. 28 Age- and sex-matched C57BL/6J mice receiving a sham operation served as the controls (n = 12). DPSCs-Null or DPSCs-HGF (1 × 106) in 200 μL of saline were injected into OVX mice via the tail vein on day 1 post OVX (n = 14). The OVX-only group received 200 μL of saline without any cells (n = 14). At 4 weeks and 6 weeks post OVX, 27 mice and 20 mice, respectively, were sacrificed for further examination. All animal experiments were performed under protocols approved by the Institutional Animal Care and Use Committee of the Beijing Institute of Radiation Medicine (reference number: AMMS-13-2015-006; Beijing, China).
Western blot
All samples were lysed in RIPA lysis buffer containing proteinase inhibitor. They were separated by sodium dodecyl sulfate polyacrylamide gel and transferred to Immobilon-P membranes (Millipore, Billerica, MA). The membranes then were blocked with 5% skimmed milk in Tris-buffered saline (150 mM of NaCl and 20 nM of Tris-HCl, pH 7.2) for 1 h at room temperature and then incubated with anti-human or mouse alkaline phosphate (ALP), anti-human or mouse runt related transcription factor 2 (Runx2), and anti-human or mouse osteocalcin (OC; Abcam, Cambridge, United Kingdom) or anti-human or mouse glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Cell Signaling Technology, Danvers, MA) antibody overnight at 4°C. They were treated with horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse antibody (ZSGB-BIO, Beijing, China) for 1 h at room temperature. The bound antibodies were visualized using SuperSignal West Pico (Thermo Fisher Scientific).
Enzyme-linked immunosorbent assay
HGF was measured with an enzyme-linked immunosorbent assay kit (R&D Systems, Inc., Minneapolis, MN) according to the manufacturer's instructions.
Bioluminescence imaging system
The animals received an intraperitoneal injection of 150 mg of luciferin/kg body weight (PerkinElmer, Waltham, MA) 10 min before imaging, and anesthesia was induced with inhaled isoflurane. Bioluminescence images were taken with the IVIS® Spectrum in vivo imaging system (PerkinElmer). The images were rebuilt, and all images were analyzed with the IVIS® imaging software (PerkinElmer).
Cytokine analysis
Mice serum was collected 3 days before cell infusion and 7, 14, 21, 28, 35, and 43 days after cell infusion. Osteoprotegerin (OPG) and receptor activator of nuclear factor kappa B ligand (RANKL) were measured using a commercially available MILLIPLEX kit (Mouse RANKL Single Plex Magnetic Bead Kit and Mouse Bone Magnetic Bead Panel). The data were collected by MAGPI and analyzed with Milliplex Analyst (Millipore) software. Interleukin (IL)-1 and tumor necrosis factor (TNF)-α in some mice serum were measured by the MILLIPLEX kit (Mouse Bone Magnetic Bead Panel; data shown in Supplementary Fig. S3).
Micro-computed tomography analysis
A high-resolution micro-computed tomography (μCT) imaging system (Inveon MM CT; Siemens, Munich, Germany) was used to assess bone structure, set to a 9.08 μm effective pixel size in its mouse model. The trabecular bone was assessed in a 0.5 mm region 0.5 mm below the distal growth plate of the femur, and the cortical bone was assessed in a 1 mm region 5 mm below the distal growth plate of the femur. The scanning, analysis, and 3D rebuild software were Inveon Acquisition Workplace (Siemens), Inveon Research Workplace (Siemens), and COBRA (Exxim, Pleasanton, CA), respectively.
Isolation and culture of mouse BM-MSCs
Mouse BM-MSCs were all collected in the traditional way. Briefly, standard culture medium, made up of α-MEM supplemented with penicillin (100 IU/mL), streptomycin (100 μg/mL; Sigma–Aldrich), and 15% FBS was used to wash the left femurs repeatedly. The suspension was then cultured in six-well plates (Corning) at 37°C in 5% carbon dioxide. After 7 days, non-adherent cells were washed with phosphate-buffered saline, and the culture medium was changed every 4 days.
Quantitative real-time polymerase chain reaction
The total RNA of mouse BM-MSCs from the four groups and human MSCs from a different source were extracted using Trizol Reagent (Invitrogen, Carlsbad, CA), and cDNA was synthesized using RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific). Quantitative polymerase chain reaction (qPCR) was carried out in an ABI 7500 fast system by using SYBR-Green I as double-strand DNA-specific binding dye. PCRs were performed in quadruplicate for each, and mRNA levels were measured using a standard curve. The cDNA of alkaline phosphate (ALP), runt related transcription factor 2 (Runx2), and osteocalcin (OC) and housekeeping gene actin cDNA were quantified in separate tubes. The sequences are shown in detail in Table 1.
The primer sequence for qRT-PCR
qRT-PCR, quantitative reverse transcription polymerase chain reaction.
Statistics
Data were analyzed using one-way analysis of variance (ANOVA) with GraphPad Prism 5 (GraphPad Software, La Jolla, CA). The expression of ALP, Runx2, and OC genes in MSCs from different sources were compared using one-way ANOVA by Dunnett's multiple comparison test. The difference between DPSC-Null and DPSC-HGF groups was analyzed using Student's t-test. Dunnett's multiple comparison test was also used to compare statistical significance of sham, DPSC-Null and DPSC-HGF groups to the OVX group as a control. The data are expressed as mean ± standard error of the mean. In all of the analyses, p < 0.05 was considered to be statistically significant.
Results
hDPSCs show a stronger osteogenic differentiation capacity in vitro
To confirm the capacity of osteogenic differentiation of hDPSCs, the RNA of DPSCs, UC-MSCs, BM-MSCs, and AD-MSCs was extracted to detect the expression of the osteogenic-related genes ALP, Runx2, and OC. The results showed that the DPSCs and BM-MSCs had a stronger osteogenic differentiation capacity (Fig. 1A–C).
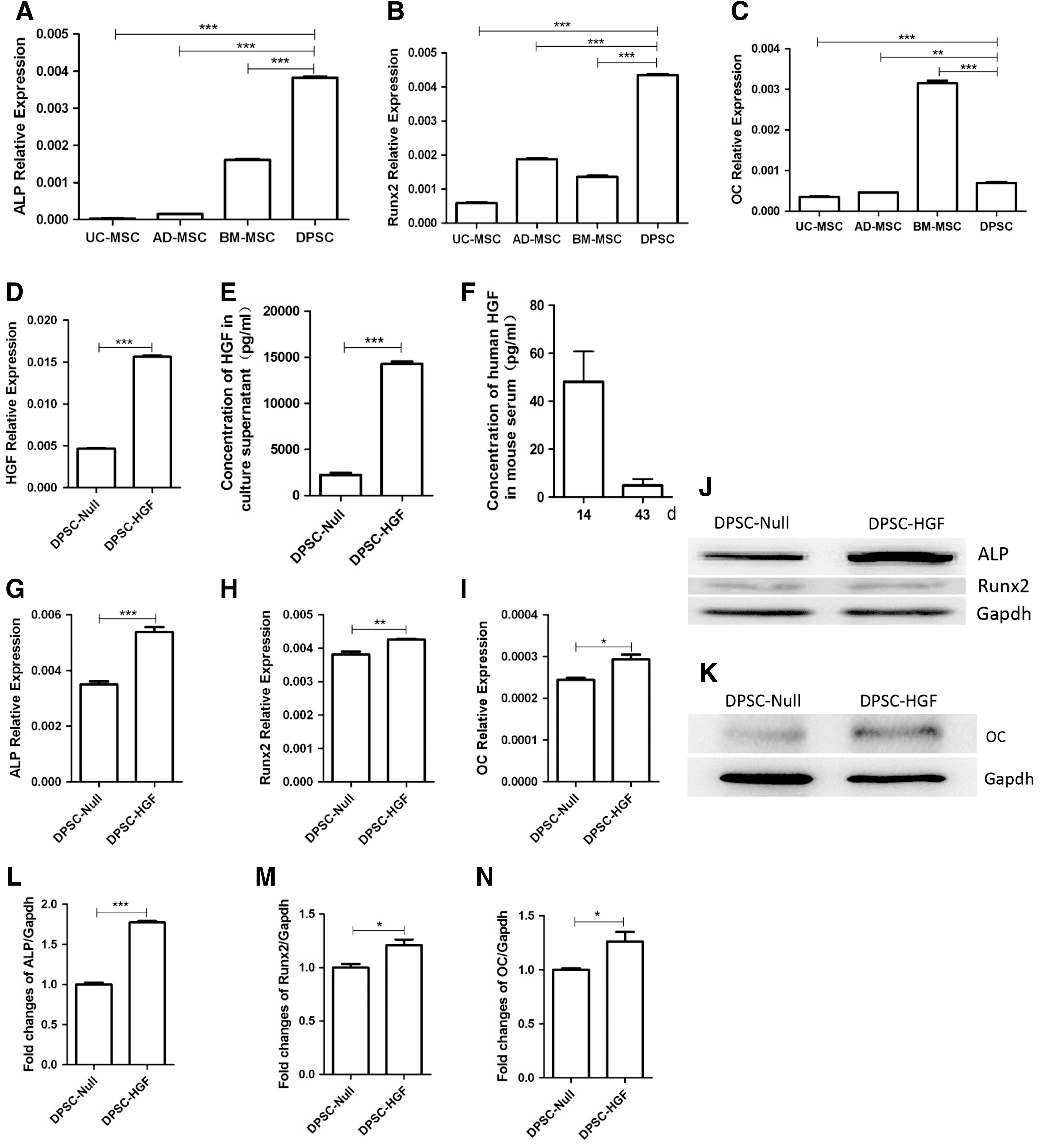
The dental pulp–derived mesenchymal stem cells (DPSCs) and hepatocyte growth factor (HGF)-modified DPSCs (DPSCs-HGF) show a stronger osteogenic differentiation capacity in vitro.
HGF gene modification promotes the expression of osteogenic genes of DPSCs
Before transplanting MSCs into the osteoporotic mice, the capacity of osteogenic differentiation of the DPSCs-HGF was evaluated in vitro. It was found that DPSCs-HGF highly expressed the HGF gene (Fig. 1D). The concentration of HGF in cell culture supernatants was 2.24 ± 0.24 ng/mL in DPSC-Null and 14.27 ± 0.28 ng/mL in DPSC-HGF. HGF gene modification could increase the secretion of HGF of DPSCs (Fig. 1E). HGF transfection results in an increased concentration of human HGF in a limited level in the serum of the DPSCs-HGF group (Fig. 1F). The concentration of human HGF was 48.07 ± 12.62 ng/mL and 4.83 ± 2.66 ng/mL on days 14 and 43, respectively, after cell infusion. The results of qPCR and Western blotting showed that HGF gene modification could enhance the expression of the osteogenic differentiation related gene (Fig. 1G–N) of DPSCs.
Distribution and survival of DPSCs in mice after transplantation via tail vein
Before evaluating the effect of MSCs in treating osteoporosis, the distribution and survival time were confirmed in vivo. To evaluate the best opportunity to transplant the cells to OVX mice, the DPSCs transduced with luciferase gene were traced, which can be detected in vivo. Then, DPSCs-Luc were transplanted into OVX-mice on day 1 or 30 after OVX. Six hours after the infusion, a bioluminogenic imaging system was used to reveal the distribution of DPSCs-Luc in mice. It showed that the majority of cells were distributed to the lungs after transplantation. Four days later, most cells then migrated to the liver (Fig. 2A). At day 28 after infusion, a strong signal was also detected in the liver, showing that DPSCs-Luc still existed. To detect the specific sites where gene-modified MSCs located further, the luciferases signals were detected in dissected organs of sacrificed mice. The results revealed that only the liver showed a strong signal (Fig. 2C and D), which suggests that the DPSCs may scarcely locate in the bone, and the effects of DPSCs for the prevention of osteoporosis may mainly depend on paracrine mechanisms rather than direct engraftment.
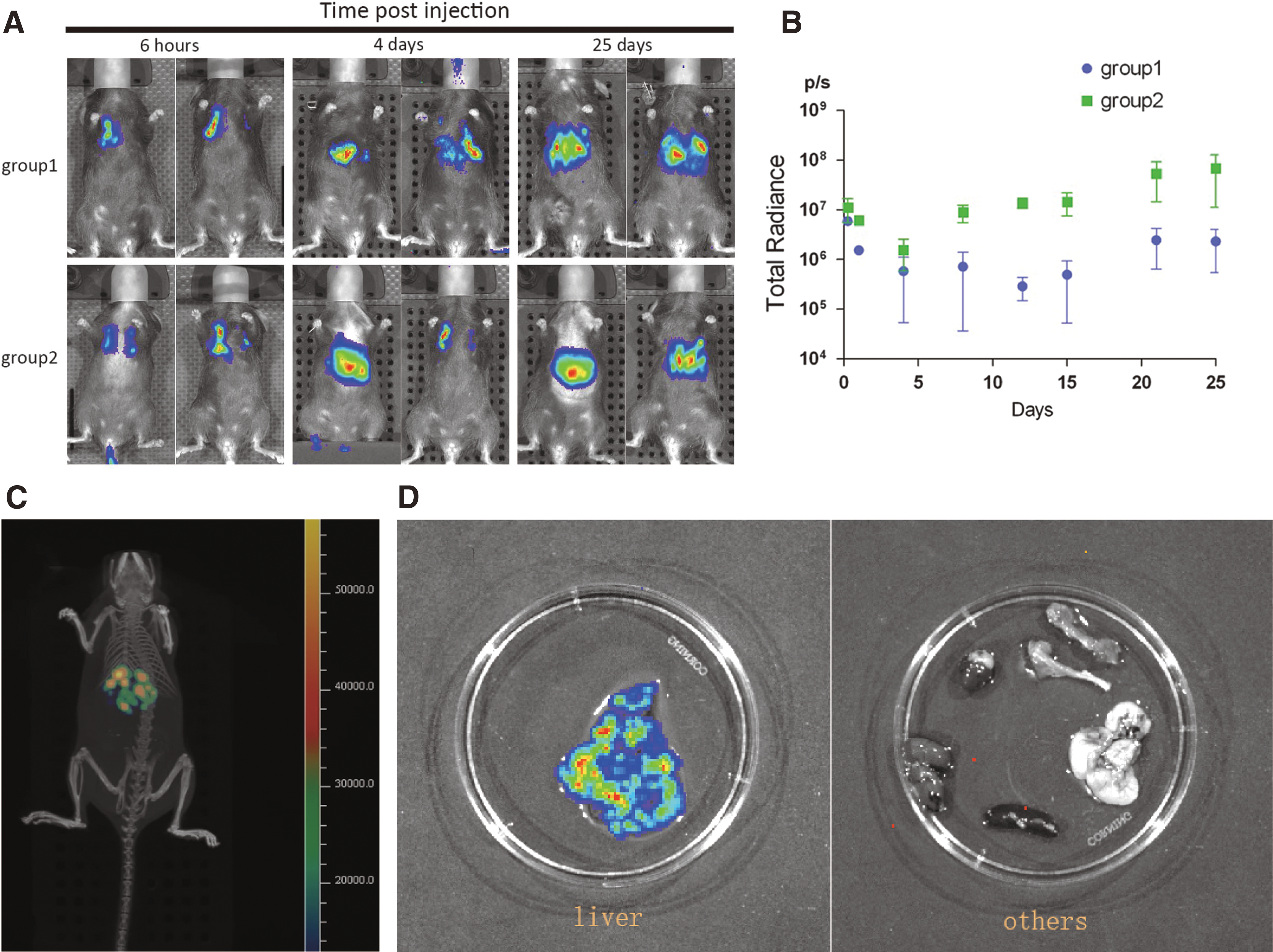
DPSCs infused through the tail vein can survive >1 month in vivo.
It was also noticed that after 6 h of cell infusion, all mice showed the same level of fluorescence intensity (total radiance of about 6 × 106 p/s). Cell infusion on day 1 after OVX (group 2) showed a much stronger signal (Fig. 2A and B) compared to cell infusion at 1 month after OVX (group 1). After 25 days of cell infusion, the total radiance of group 1 was about 4 × 106 p/s, but for group 2 it was about 5 × 107 p/s. This suggests that cells infused within 1 day after OVX can survive more easily in the mouse, and cell transplantation should be performed as early as possible.
One-time infusion of DPSCs-HGF prevented OVX-induced early bone loss
To determine whether transplantation of DPSCs-Null or DPSCs-HGF ameliorates the osteoporotic phenotype, these cells were infused into OVX mice and their effects were analyzed at 4 or 6 weeks after treatment. According to previous reports, the distal metaphysis of the femur is the area most sensitive to estrogen deficiency. 29 The right femurs were separated for μCT analysis. The results indicated that OVX induced significant bone loss in the trabecular bone of the distal femur metaphysis when compared to the sham group (Fig. 3B), as shown by decreased bone volume/total volume (BV/TV; Fig. 3C), trabecular thickness (Tb.Th; Fig. 3D), trabecular number (Tb.N; Fig. 3E), and trabecular bone mineral density (Tb.BMD; Fig. 3F), along with increased trabecular space (Tb.Sp; Fig. 3G), bone surface area/bone volume (BSA/BV; Fig. 3H), and trabecular pattern factor (Tb.PF; Fig. 3I) in OVX mice. Micro-CT analysis also showed that after 4 weeks of DPSCs-HGF transplantation, there was a significant increase in BV/TV (∼23%; Fig. 3C), Tb.Th (∼13%; Fig. 3D), Tb.N (∼18%; Fig. 3E), and TB.BMD (∼22%; Fig. 3F), along with decreased Tb.sp (∼22%; Fig. 3G), BSA/BV (∼15%; Fig. 3H), and Tb.PF (∼23%; Fig. 3I) compared to the OVX group. However, although transplantation could alleviate osteoporosis in the DPSCs-Null group, it did not reach statistical significance (Fig. 3C–I). Interestingly, with the observation time elongation, at 6 weeks after cell infusion, there was a significant difference in the DPSC-Null group compared to the OVX group, although the therapeutic effect was still better in the DPSCs-HGF group than in the DPSCs-Null group, but there was no statistical significance (Fig. 3K–Q) between the two groups. These results suggest that HGF might promote the DPSCs to prevent partial bone loss after OVX in a short time compared to DPSCs alone.
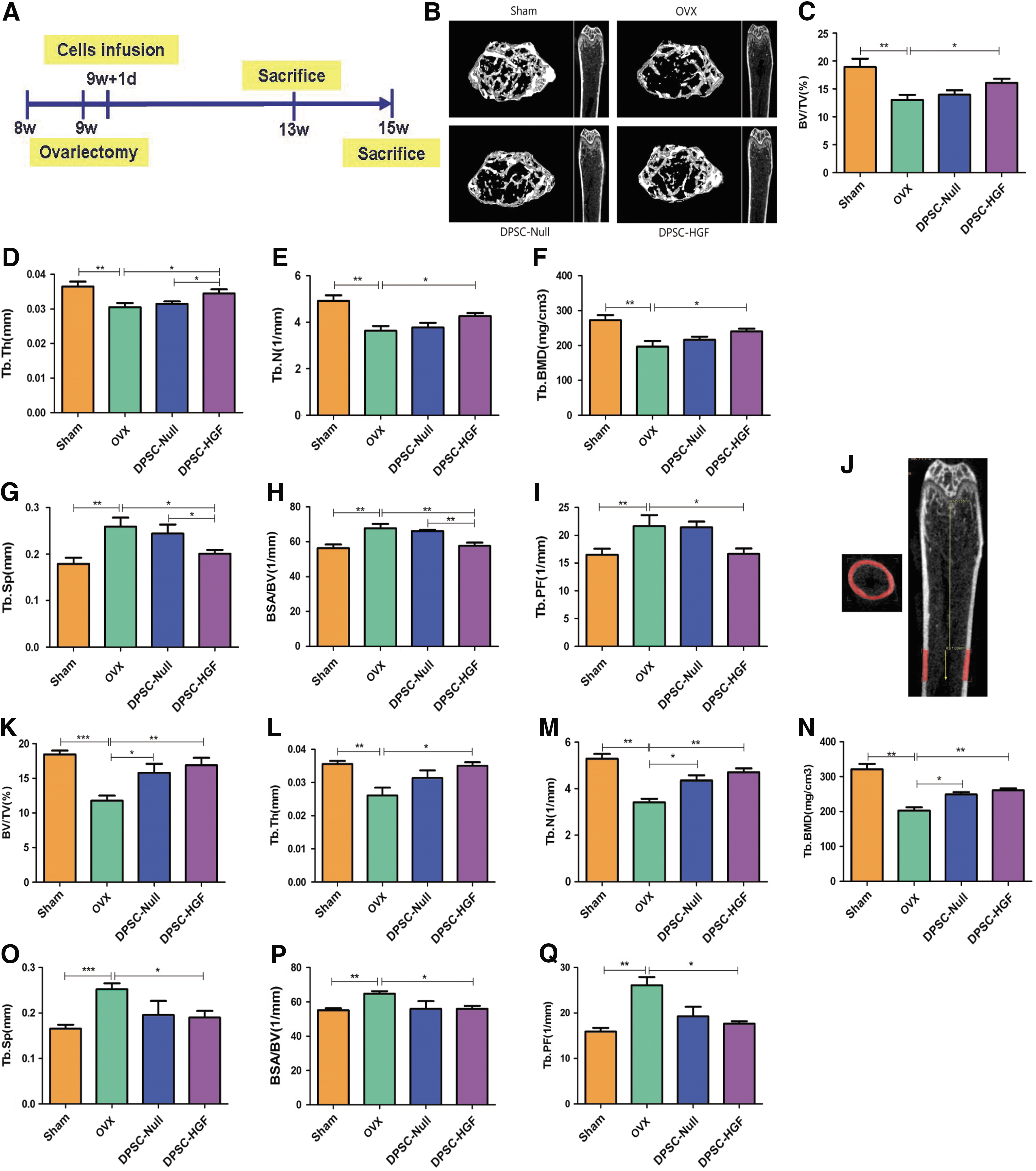
One-time infusion of DPSCs-HGF prevented OVX-induced early bone loss.
The cortical bone mass of all groups was nearly the same, as shown by the cortical bone parameters, including total cross-sectional volume (Tt.Vo), cortical bone volume (Ct.Vo), cortical bone fraction (Ct.Vo/Tt.Vo), and cortical BMD (Ct.BMD; data shown in Supplementary Fig. S4), suggesting that 1 month after resection, bone loss in OVX mice takes place in the cancellous bone rather than the cortical bone.
DPSC therapy promotes the expression of bone formation related genes
To assess the impact of DPSCs-Null and DPSCs-HGF transplantation on osteoclasts, the concentrations of RANKL and OPG were examined in serum. RANKL is essential to activate osteoclasts. As shown in Fig. 4A, the concentration of RANKL increased evidently in the OVX group, and the expression levels in cell-treated groups were lower than those in the OVX group (Fig. 4A). The concentration of OPG, a natural antagonist to RANKL-induced osteoclastogenesis and bone resorption, increased significantly 3 days after OVX compared to the sham group. However, 7 days after cell infusion, OPG expression in the cell-treated groups was the same as in the sham group and stayed at a stable level, but OPG in the OVX group gradually decreased to a low level (Fig. 4B).
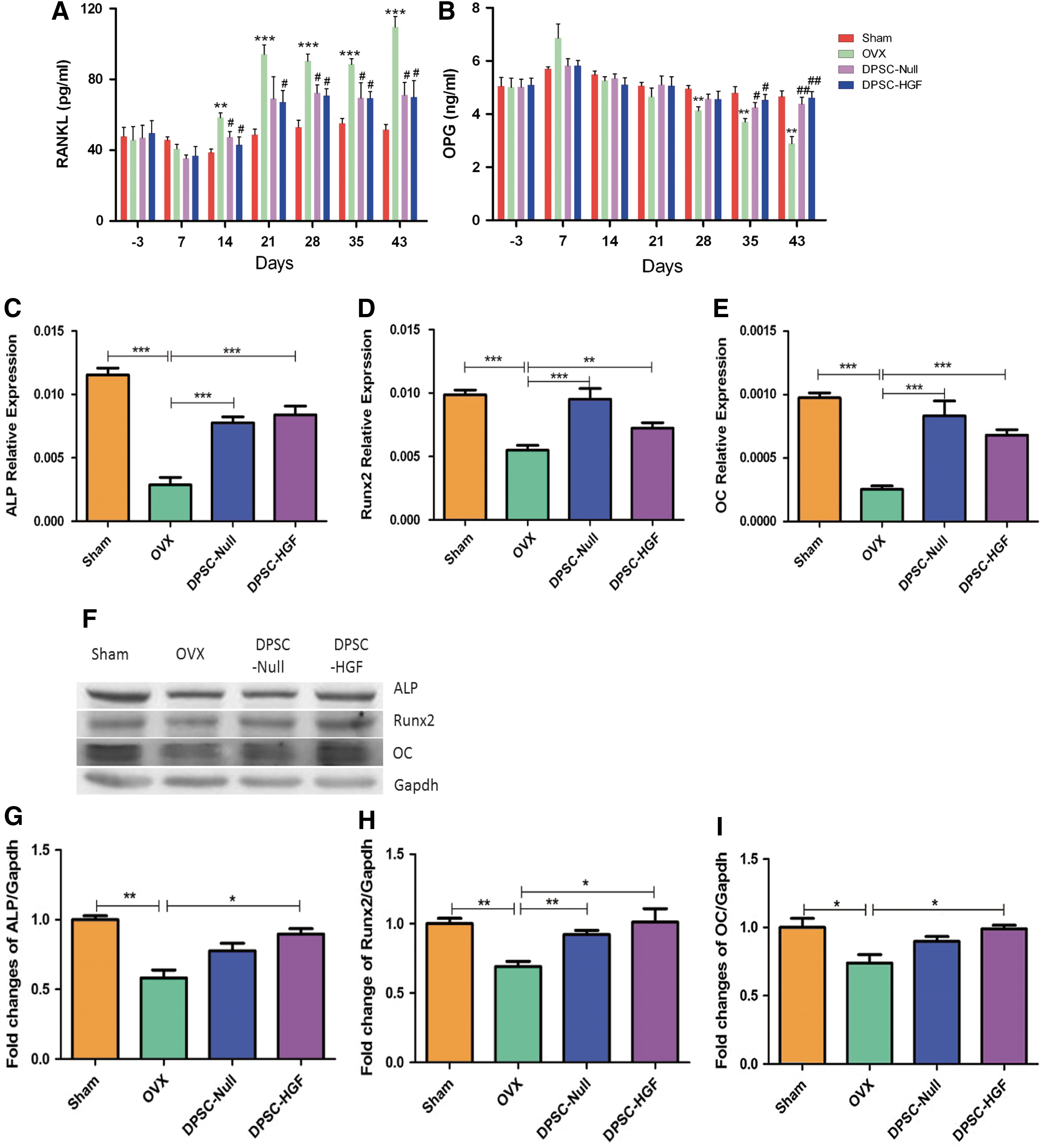
DPSC therapy promotes the expression of bone formation related genes.
To elucidate the mechanisms of DPSCs further, the BM-MSCs were isolated and cultured 6 weeks after OVX, and the expressions of different osteogenic related genes and proteins were detected. The expression of genes related to bone formation in mouse BMMSCs were decreased in the OVX group. The cell-treated groups stayed at the same level as the sham group (Fig. 4C–E), and the protein results were almost the same (Fig. 4F–I).
Discussion
Osteoporosis is a debilitating disease characterized by systemic bone loss in the musculoskeletal system. Clinically, the most frequently used treatment includes hormone replacement therapy, selective estrogen receptor modulators, bisphosphonates, and parathyroid hormone. 30 These agents mainly focus on inhibiting bone resorption and preventing further bone loss. However, they cause serious side effects, such as osteonecrosis of the jaw, atypical femoral fractures, and an increased risk of cancer and heart disease. 30,31
MSC administration is a novel strategy for treating bone- and joint-related diseases. Cartistem®, derived from cord blood and produced by Medipost (Seongnam, South Korea), has been used to treat degenerative arthritis and cartilage injury of the knee joint. MPC, produced by Mesoblast (New York, NY), has been used to repair bone.
One factor that affects the therapeutic effect of MSCs is their source. This study compared the osteogenic capacities of MSCs derived from different sources, including DPSCs derived from dental pulp, UC-MSCs derived from the umbilical cord, BM-MSCs derived from bone marrow, and AD-MSCs derived from adipocytes. qPCR data revealed that MSCs express osteogenesis-related genes such as ALP, Runx2, and OC. ALP is a recognized marker of osteoblast differentiation and plays an important role in bone calcification. ALP in osteoblasts produces phosphoric acid, which reacts with calcium to produce calcium phosphate and is deposited in the bone. 32 Runx2 is a cell-specific member of the Runt family of transcription factors, and plays a critical role in osteoblastic differentiation. Runx2 expression is indispensable and is sufficient to induce osteoblastic differentiation for subsequent regulation of the expression of a variety of genes characteristic of the osteoblastic phenotype, including Osteocalcin and α1(I) Collagen. 33,34 OC is a marker for terminal osteoblast differentiation. 35 Overexpression of these genes could prevent the progress of osteoporosis. The results (Fig. 1A–C) showed higher expression of these genes in DPSCs and BM-MSCs, which means DPSCs and BM-MSCs possess stronger osteogenic capacity than that of UC-MSCs and AD-MSCs. Thus, DPSCs and BM-MSCs are considered ideal candidates for cell therapy on osteoporosis. Furthermore, DPSCs are more suitable for cell therapy than BM-MSCs because they have stronger proliferation ability and are more easily obtained. It is possible to harvest >6 × 109 DPSCs at P6 from one dental pulp.
Another factor that affects the therapeutic effect of MSCs is injection time. To determine the optional opportunity of MSC transplant, IVIS® Spectrum in vivo imaging system (PerkinElmer) was used. The DPSCs were administrated into OVX mice, and the amount and site of homing cells were detected in vivo at day 1 and day 30 post transplantation. It was observed that transplanted cells may survive >1 month in vivo. It is consistent with previous reports that MSCs can survive long term in vivo. 36,37 Because the cell infusion on day 1 after OVX (group 2) showed a much stronger fluorescent signal compared to the cell infusion at 1 month after OVX (group 1), it was recommended that MSCs therapy should be applied in the early stage of OVX. The DPSCs mainly resided in the liver 4 days after the cells were transfused. So, liver function was evaluated by detecting ALT and AST in serum on days 2, 12, 21, and 30 after cell infusion by using an automatic biochemical analyzer. The concentrations of ALT and AST in the three groups—OVX group, DPSCs-Null group, and DPSCs-HGF group—showed no statistical difference. This indicates that the 1 month survival of DPSCs-Null or DPSCs-HGF in the liver has no effect on liver function (Supplementary Fig. S5).
The original intention of stem-cell therapy was to induce bone formation via proliferation and differentiation of bone progenitor cells. Some researchers injected MSCs directly into the bone-marrow cavity of femurs in OVX models to achieve a high engraftment efficiency of transplanted MSCs. 19,38,39 However, local infusion of stem cells only improves the limited area where they are implanted and is not suitable for the treatment of osteoporosis, which is a systemic disease. Scientists turned to tail-vein injection, predicting the cells to reach the appropriate physiological areas that required repaired. Excitingly, 2 months post injection, the MSCs homed to bone marrow, and μCT showed that bone volume of the OVX mice had been restored to the normal level. 40 The data from the present study indicate that DPSCs scarcely home to the bone after the OVX procedure. OVX-induced osteoporosis is different from other diseases, such as injured and inflammatory disease. Maybe in the early stage of the OVX procedure, there was not an efficient chemotactic factor to attract a large amount of MSCs to home to the bone.
Human stem cells hardly spontaneously engraft and differentiate to functional osteoblasts at relevant areas in mice. Some investigators have used genetically modified MSCs, such as CXCR-4, Cbfa-1, RANK-Fc, and so on, to solve these problems. 41,42 Modification with these genes promotes the homing and osteogenic differentiation of MSCs. The present study introduced the HGF gene, which promotes cell survival, suppresses chronic inflammation, stimulates osteoblast proliferation and participates in bone remodeling, to the DPSCs to prevent bone loss. It is known that HGF has a therapeutic effect on OVX mice. However, HGF protein has limited clinical application because it is extremely unstable in the blood circulation, with a half-life of <15 min. As a result, it is almost impossible to sustain a constantly high level of exogenous HGF in the circulation, even with repeated injections of HGF protein at short intervals. A logical way to overcome this problem is to develop a gene transfer strategy allowing the persistent expression of the HGF protein in vivo. The present in vivo results show that DPSCs-HGF transplantation could increase BV/TV (∼23%), Tb.Th (∼13%), Tb.N (∼18%), and BMD (∼22%), along with decreased Tb.sp (∼20%), Tb.PF (∼23%), and BSA/BV (∼15%), significantly when compared to the OVX group and DPSCs-Null group at the early stage (Fig. 3C–I). Though there was no significant statistical difference between the DPSCs-HGF group and the DPSCs-Null group in Tb.Th, Tb.N, and BSA/BV, the tendency showed that the effect of the DPSCs-HGF group was stronger than that of the DPSCs-Null group. With the passing of time, the DPSC-Null group could also play a role in preventing bone loss (Fig. 3K–Q). That means that HGF gene modification could promote a curative effect, especially in the early stage, through increasing the secretion of bioactive factors by DPSC-HGF. These results reveal that HGF gene modified DPSCs could be an effective therapeutic means to treat osteoporosis. The possible mechanism is relies mainly on the increased bioactive factors that are secreted by DPSCs-HGF. The in vitro results of qPCR and Western blotting showed that HGF gene modification could enhance the expression of osteogenic differentiation related protein of DPSCs, including ALP, Runx2, and OC.
This study did not observe the local engraftment of DPSCs, which means the therapeutic effects of DPSCs did not rely mainly on their engraftment and differentiation ability to functional osteoblasts. Enhanced paracrine secretion of cytokines may contribute to their regenerative effect in osteoporosis. In this study, several cytokines, such as RANKL and OPG, were detected by the MILLIPLEX kit. RANKL is an osteoclast differentiation and activation factor, 43 while OPG acts as decoy receptor for RANKL and thereby neutralizes its function in osteoclastogenesis. OPG could inhibit the activation of osteoclasts and promote osteoclast apoptosis in vitro. Bone homeostasis seems to depend on the local RANKL/OPG ratio. 44 The results show that the concentration of RANKL increased evidently in the OVX group, and the concentration level of the cell-treated group was lower than that of the OVX group. Conversely, the concentration of OPG in the cell-treated group stayed at a stable level but gradually decreased in the OVX group. These results were further confirmed by gene expression of isolated and cultured mouse bone marrow–derived MSCs using qPCR and Western blotting. In conclusion, HGF-modified DPSCs could be an effective therapeutic strategy for the treatment of bone loss. The therapeutic effects of DPSCs rely mainly on secreting bioactivators to prevent bone loss at the early stage of OVX osteoporosis.
Conclusions
One-time infusion of hDPSCs-HGF prevents bone loss in the early phase of OVX osteoporosis, and paracrine mechanisms may explain their therapeutic and preventive effects.
Footnotes
Acknowledgments
This work was supported by grants from National Natural Science Foundation of China (No.81573086, 81372924) and Chinese High-Tech “863” Program (2014AA020515).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.