
Abstract
Due to its minimal size and lack of bacterial backbone sequences, minicircle (MC) DNA presents a promising alternative to plasmid DNA (pDNA) for non-viral gene delivery in terms of biosafety and improved gene transfer. Here, luciferase pDNA (pCMV-luc) and analogous MC DNA (MC07.CMV-luc) were formulated into polyplexes with c-Met targeted, PEG-shielded sequence-defined oligoaminoamides, or linear PEI (linPEI) as standard transfection agent. Distinct physicochemical and biological characteristics were observed for polyplexes formed with either pDNA or MC DNA as vectors. The carriers were found to dominate the shape of polyplexes, whereas the DNA type was decisive for the nanoparticle size. c-Met-targeted, tyrosine trimer-containing polyplexes were optimized into compacted rod structures with a size of 65–100 nm for pDNA and 35–40 nm for MC. Notably, these MC polyplexes display a lack of cell cycle dependence of transfection and a ∼200-fold enhanced gene transfer efficiency in c-Met-positive DU145 prostate carcinoma cultures over their tyrosine-free pDNA analogues.
Introduction
A
Gene transfer can be facilitated by physical and biochemical DNA delivery techniques. For example, nanosized polyplexes are formed by complexation of negatively charged nucleic acid with positively charged cationic oligomers. 5–6,9 Recently, precise analogs of linear polyethylenimine (linPEI), a frequently used transfection carrier, 26 –28 were generated by solid-phase supported syntheses. 29,30 By this sequence-defined assembly process, libraries of oligoaminoamides can be generated presenting different functional modifications (nucleic acid compaction and nanoparticle stabilization, surface shielding, receptor binding, endosomal escape) in defined topologies, which might be most suitable for the specific cargo and application. 31 –36
Apart from enhancing the gene transfer process by improving carrier molecules, DNA vectors can also be optimized. 37 –40 Standard plasmids contain a eukaryotic expression cassette (the gene of interest and regulatory sequences) and a backbone, including bacterial sequences. Antibiotic resistance-encoding genes and the origin of replication are necessary to maintain and amplify pDNA vectors in the bacterial hosts. However, bacterial sequences, including CpG dinucleotide motifs, reduce the efficacy, biocompatibility, and safety of pDNA vectors. Regulatory agencies recommend avoidance of antibiotic resistance-encoding genes in production of plasmids for therapeutic use. 41 Minicircle (MC) DNA contains only the therapeutic gene of interest and regulatory sequences. Bacterial backbone sequence and antibiotic-resistant genes are excluded. This may result in reduced risk of spreading antibiotic-resistant genes, enhanced immunocompatibility, improved transfer efficiency, and prolonged transgene expression compared to those of standard plasmids. 42 –47 In fact, MC DNA has already displayed greatly improved transgene expression in several in vitro 25,48,49 and in vivo studies. 44 –47,50 –62 Different nucleic acid cargos, due to their different size or nature, are known to differ in their formulation requirements. 63,64 Therefore, the present study compared physicochemical and transfection characteristics of polyplexes formed with pDNA (pCMV-luc) or MC DNA (MC07.CMV-luc) and linPEI, as well as untargeted and targeted oligoaminoamides.
Materials and Methods
Materials
Linear polyethylenimine 22 kDa (linPEI) was prepared as previously reported. 65 2-Chlorotrityl chloride resin was obtained from Iris Biotech (Marktredwitz, Germany). Benzotriazole-1-yl-oxy-tris-pyrrolidino-phosphonium hexafluorophosphate (PyBOP), 2-(1H-benzotriazol-1-yl)-1,1,3,3-tetramethyluronium-hexafluorophosphat (HBTU) and syringe reactors were purchased from Multisyntech GmbH (Witten, Germany). All amino acids and peptide-grade dimethylformamide (DMF), as well as N-methylpyrrolidon (NMP), N,N-diisopropylethylamine (DIPEA), and trifluoroacetic acid (TFA), were used from Iris Biotech. 1-Hydroxy-benzotriazole (HOBt), triisopropylsilane (TIS), Triton-X, and 1,2-ethanedithiol (EDT) were purchased from Sigma–Aldrich (Munich, Germany). n-Hexane and tert-butyl methyl ether (MTBE) were obtained from Brenntag (Mühlheim an der Ruhr, Germany). Dichloromethane was obtained from Bernd Kraft (Duisburg, Germany). Fmoc-N-amido-dPEG24-acid® was purchased from Quanta Biodesign (Powell, OH). All solvents and other reagents were acquired from Sigma–Aldrich, Iris Biotech, Merck (Darmstadt, Germany), or AppliChem (Darmstadt, Germany). Cell culture media, antibiotics, and fetal calf serum (FCS) were purchased from Invitrogen (Karlsruhe, Germany), HEPES from Biomol GmbH (Hamburg, Germany), and glucose from Merck. All flasks and multi-well plates were manufactured by TPP (Trasadingen, Switzerland). Luciferase cell culture lysis buffer and D-luciferin sodium salt were obtained from Promega (Mannheim, Germany).
Vectors
pCMV-luc (PF461) and MC07.CMV-luc were produced and purified by PlasmidFactory GmbH (Bielefeld, Germany).
Oligoaminoamide synthesis
Oligoaminoamides for use as DNA carriers were synthesized by solid-phase-supported synthesis using Fmoc-chemistry, protected L-amino acids, artificial amino acids, and other building blocks.
29,30,34,66
Syntheses were performed (semi-)manually or by the fully automated Syro Wave peptide synthesizer (Biotage, Uppsala, Sweden), equipped with a microwave cavity. The exact sequences of all oligomers (C- to N-terminus) can be found in Supplementary Table S1 (Supplementary Data are available online at
Targeting peptide synthesis was carried out using the automated Syro Wave peptide synthesizer equipped with a microwave cavity. Automated coupling was carried out with 4 eq of HBTU, 8 eq of DIPEA, 4 eq of HOBt, and 4 eq of Fmoc protected amino acid per free amine on the solid support. The solution was supplemented with 1% Triton X-100 (v/v). Amino acids and HOBt were dissolved in NMP, supplemented with Triton-X. HBTU was dissolved in DMF, and DIPEA was diluted with NMP. Double couplings have been performed at 50 °C for 12 min prior to five repetitive washing steps with DMF. Fmoc removal was conducted by standard deprotection methods with 40% piperidine in DMF (v/v) supplemented with 1% Triton-X for 5 × 10 min.
During manual peptide synthesis, coupling steps were carried out with 4 eq of Pybop, 8 eq of DIPEA, 4 eq of HOBt, and 4 eq of Fmoc protected amino acid per free amine on the solid support. Amino acids were dissolved together with DIPEA in DCM. Pybop and HOBt were dissolved in DMF. Solutions were combined and coupling was conducted for 60 min at steady shaking with an overhead shaker. Then the solutions were discarded, and three washing steps each, first with DMF followed by DCM, were carried out to remove remaining activated amino acid. A Kaiser test 67 was performed to verify successful coupling. In case of a positive result, the coupling step was repeated. Fmoc removal was conducted by standard deprotection methods with 20% piperidine in DMF (v/v) for 5 × 10 min. Afterwards, the resin was washed with DMF followed by DCM, and another Kaiser test was performed. In this case, a positive result is expected to continue with the attachment of the next amino acid. After the final coupling step, the resins were dried and the products cleaved off the resin for 90 min using a mixture of TFA/EDT/H2O/TIS 94:2.5:2.5:1 (v/v) at a ratio of 10 mL/g of resin. The cleaved oligomers were purified by size exclusion chromatography (SEC) performed with 10 mM of hydrochloric acid/acetonitrile 7:3 as solvent. An ÄKTA purifier system (GE Healthcare Biosciences, Uppsala, Sweden) equipped with a Sephadex G-10 column and a P-900 solvent pump module, a UV-900 spectrophotometrical detector, a pH/C-900 conductivity module, and a Frac-950 automated fractionator was used. The product fractions were collected and combined prior to lyophilization. Analytical data of proton nuclear magnetic resonance ( 1 H-NMR) and reverse-phase high-performance liquid chromatography (RP-HPLC) can be found in Supplementary Figs. S1–S4.
Synthesis of three-arm oligomers
Oligomer 689 was prepared as described previously. 34,66 The novel tyrosine-rich three-arm oligomer 849 was synthesized as follows in detail. The resin, preloaded with Fmoc-L-Cys(Trt)-OH, was deprotected with piperidine, and Fmoc-L-Tyr(tbu)-OH was attached three times. Then Fmoc-L-His(Trt)-OH and Fmoc-Stp-OH were coupled manually, alternately three times. The first arm was terminated with a fourth Fmoc-L-His(Trt)-OH. Then Fmoc-L-Lys(Fmoc)-OH was introduced for symmetrical branching. The two remaining arms were synthesized in parallel as follows: Fmoc-L-His(Trt)-OH and Fmoc-Stp-OH (repeated three times in alternate order) were attached to the solid phase. A fourth Fmoc-L-His(Trt)-OH and three times Fmoc-L-Tyr(tbu)-OH were coupled prior to a terminating Boc-L-Cys(Trt)-OH.
Synthesis of cmb-PEG oligomers
The hepatocyte growth factor (HGF) receptor/c-Met targeting peptid for oligomers 442 and 852 was prepared as described previously using the Syro Wave synthesizer equipped with a microwave cavity. 68 Synthesis of the cationic backbone of oligomer 442 was conducted following recent reports. 34,66 The novel tyrosine-trimer containing cmb-PEG oligomer 852 was synthesized as follows, starting with a resin preloaded with Fmoc-L-Lys(Dde)-OH. After deprotection of the α-amine with piperidine, the c-Met binding peptide (cmb) amino acid sequence (KSLSRHDHIHHH) was assembled step-by-step using the microwave cavity of the Syro Wave synthesizer. The peptide sequence was terminated by Boc modification with a 12-fold excess of di-tert-butyl dicarbonate and 24-fold excess of DIPEA dissolved in NMP for 1 h. After a negative Kaiser test, the side chain protecting Dde group of the initial L-lysine was removed by 20 cycles of incubation with 4% hydrazine hydroxide in DMF (v/v) for 3 min each. Then the polymeric backbone was attached by manual synthesis. First, Fmoc-dPEG24-OH followed by Fmoc-L-His(Trt)-OH and a Fmoc-L-Lysine(Fmoc)-OH was attached to the freed ɛ- amine. The two cationic arms were built in parallel. Therefore, Fmoc-L-His(Trt)-OH and Fmoc-Stp-OH were attached alternating four times. Then another Fmoc-L-His(Trt)-OH and three sequential Fmoc-L-Tyr(tbu)-OH were attached. The synthesis was terminated by coupling of Boc-L-Cys(Trt)-OH.
Polyplex formation
Indicated amounts of pCMV-luc or MC07.CMV-luc and the calculated amounts of oligomer or oligomers combination at indicated N/P (nitrogen/phosphate) ratios were diluted in separate tubes of equal volumes of HBG (20 mM of HEPES, 5% glucose, pH 7.4) each. Only protonatable nitrogens of the Stp units and N-terminal amines of cysteine residues were considered in the N/P calculations. In the case of oligomers combination, cmb-targeted oligomer was mixed with three-arm oligomer at an optimized 70:30 cationic ratio to reach the total N/P ratio of 6 or 12 prior to polyplex formation with the DNA. The oligomer solution was added to the nucleic acid solution, mixed by pipetting, and incubated for 30 min at room temperature under exposure to air oxidation in a closed Eppendorf reaction tube.
Particle size and zeta potential
Particle size and zeta potential of formulations were measured in a folded capillary cell (DTS 1070) by dynamic laser-light scattering using a Zetasizer Nano ZS with backscatter detection (Malvern Instruments, Malvern, United Kingdom). Experiments were performed with 2 μg of pCMV-luc or MC07.CMV-luc and oligomer or oligomers combination at N/P ratio 12 in a total volume of 200 μL of HBG. For size measurements, the equilibration time was 0 min, the temperature was 25°C, and an automatic attenuator was used. The refractive index of the solvent was 1.330, and the viscosity was 0.8872 mPa s; the refractive index of polystyrene latex (1.590) was fixed. Results were plotted as Z-Average and standard deviation (SD) out of three runs, with 12 sub-runs each. For zeta potential measurements, the samples were diluted to 800 μL in 20 mM of HEPES buffer (pH 7.4). Zeta potentials were calculated by the Smoluchowski equation and are displayed as an average (mV) of three runs with up to 15 sub-runs each.
Transmission electron microscopy
For transmission electron microscopy (TEM), samples were prepared with 200 ng of pCMV-luc or MC07.CMV-luc and oligomer or oligomers combination at N/P ratio 12 in Millipore water. The formvar/carbon-coated 300 mesh copper grids (Ted Pella, Inc., Redding, CA) were activated by mild plasma cleaning. Then, the copper grids were incubated with 20 μL of the polyplex solution for 2.5 min. Excess liquid was blotted off using filter paper. Prior to staining, the copper grids were washed with 5 μL of staining solution for 5 s. Afterwards, the copper grids were incubated with 5 μL of a 2% aqueous uranylformate solution for 20 s. Excess liquid was again blotted off using filter paper, followed by air-drying for 30 min. Samples were then characterized using a JEM-1011 (Jeol, Freising, Germany) operating at 80 kV.
Ethidium bromide compaction assay
Polyplexes containing 2 μg of pCMV-luc or MC07.CMV-luc were formed at N/P ratio 12 in a total volume of 200 μL of HBG. In parallel to these polyplexes, the following were prepared: HBG buffer (blank value), linPEI polyplexes, and 2 μg of pCMV-luc or MC07.CMV-luc in 200 μL of HBG buffer, which was considered as maximum ethidium bromide (EtBr) fluorescence intensity (100% value). After a 30 min incubation time, 700 μL of EtBr solution (c = 0.5 μg/mL) was added to each sample. The fluorescence intensity of EtBr was measured after an additional 3 min incubation at the excitation wavelength ex = 510 nm and emission wavelength em = 590 nm using a Cary Eclipse spectrophotometer (Varian, Germany). The fluorescence intensity of EtBr was calculated in relation to the 100% value. To investigate polyplex stability against polyanionic stress, 250 IU of heparin (Ratiopharm, Ulm, Germany) was added to the each sample after EtBr addition, and the fluorescence intensity of EtBr was measured again.
Cell culture
Human prostate cancer cell line (DU145) was cultured in RPMI-1640 medium. Cell culture media was supplemented with 10% fetal bovine serum (FBS), 4 mM of stable glutamine, 100 IU/mL of penicillin, and 100 μg/mL of streptomycin. DU145 cell line was cultured at 37°C and 5% CO2 in an incubator with a relative humidity of 95%.
Gene transfer
DU145 cells (10,000 cells/100 μL) were seeded 24 h prior to nucleic acid delivery in 96-well plates. Transfection efficiency of oligomers was evaluated using 200 ng of pCMV-luc or MC07.CMV-luc per well. Additionally, 320 ng of pCMV-luc per well were used in the case of equimolar experiment. Polyplexes were formed at different N/P ratios in a total volume of 20 μL of HBG. linPEI (N/P 9) was used as a positive control, and HBG buffer was used as a negative control. Before treatment, the cell culture medium was exchanged with 80 μL of fresh medium containing 10% FBS. Polyplex solution was added to each well and incubated on cells at 37°C for a determined period of time (0.75 or 24 h). In the case of 0.75 h incubation time, medium was replaced 0.75 h after transfection by fresh medium, and cells were further incubated for 23 h. In the case of 24 h incubation time, cells were incubated with polyplex solution for 24 h after initial transfection. All experiments were performed in quintuplicate. For all experiments 24 h after initial transfection, cells were treated with 100 μL of luciferase cell culture 5 × lysis buffer. Luciferase activity in the cell lysate was measured by using a Centro LB 960 plate reader luminometer (Berthold Technologies, Bad Wildbad, Germany) and LAR buffer supplemented with 1 mM of luciferin solution. Transfection efficiency was evaluated as relative light units (RLU) per well (10,000 cells).
Cell cycle dependent transfections
DU145 cells were seeded 24 h prior to treatment with thymidine into 96-well plates with a density of 7.5 × 103 cells in 100 μL of growth media. A double-thymidine block was applied, as described in the Supplementary Methods, to synchronize cells in specific phases of cell cycle. At the end of the second block, the cells were washed twice with phosphate-buffered saline (PBS) and released in 80 μL of thymidine free growth medium (second release). Polyplexes were prepared as described containing 200 ng of pCMV-luc or MC07.CMV-luc in 20 μL of HBG. Polyplex solutions were added to each well at different time points (0, 6, 8, or 12 h) after second release. Transfection medium was replaced after 45 min at 37 °C, and cells were further incubated at 37°C for 24 h. All experiments were performed in quintuplicate. Afterwards, luciferase activity in the cell lysate was determined as described above, and metabolic activity of transfected cells as described below.
Metabolic activity of transfected cells
The cells were transfected as described above. At 24 h after initial transfection, 10 μL of MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) was added to each well, reaching a final concentration of 0.5 mg/mL. Medium with unreacted dye was removed after an incubation time of 2 h at 37°C. The 96-well plates were stored at −80 °C for at least 1 h, and afterwards the purple formazan product was dissolved in 100 μL dimethyl sulfoxide per well. The absorbance was determined by using a microplate reader (Tecan Spectrafluor Plus, Tecan, Switzerland) at 530 nm with background correction at 630 nm. The relative cell viability (%) related to the buffer-treated control cells was calculated as ([A] test/[A] control) × 100%.
Statistical analysis
Results are presented as arithmetic mean ± SD and the number of replicates. Statistical significances were analyzed using Student's two-tailed t-test. Significance levels were indicated with the following symbols: ns, p > 0.05; *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; and ****p ≤ 0.0001.
Results and Discussion
Vector constructs and polyplex design
pCMV-luc or MC07.CMV-luc were used for polyplex formation (see Fig. 1A). pCMV-luc is a double-stranded plasmid DNA of 6233 bp encoding the firefly luciferase reporter gene under control of the cytomegalovirus promoter, while MC07.CMV-luc is smaller MC DNA (3,881 bp) encoding the same gene under the same promoter.
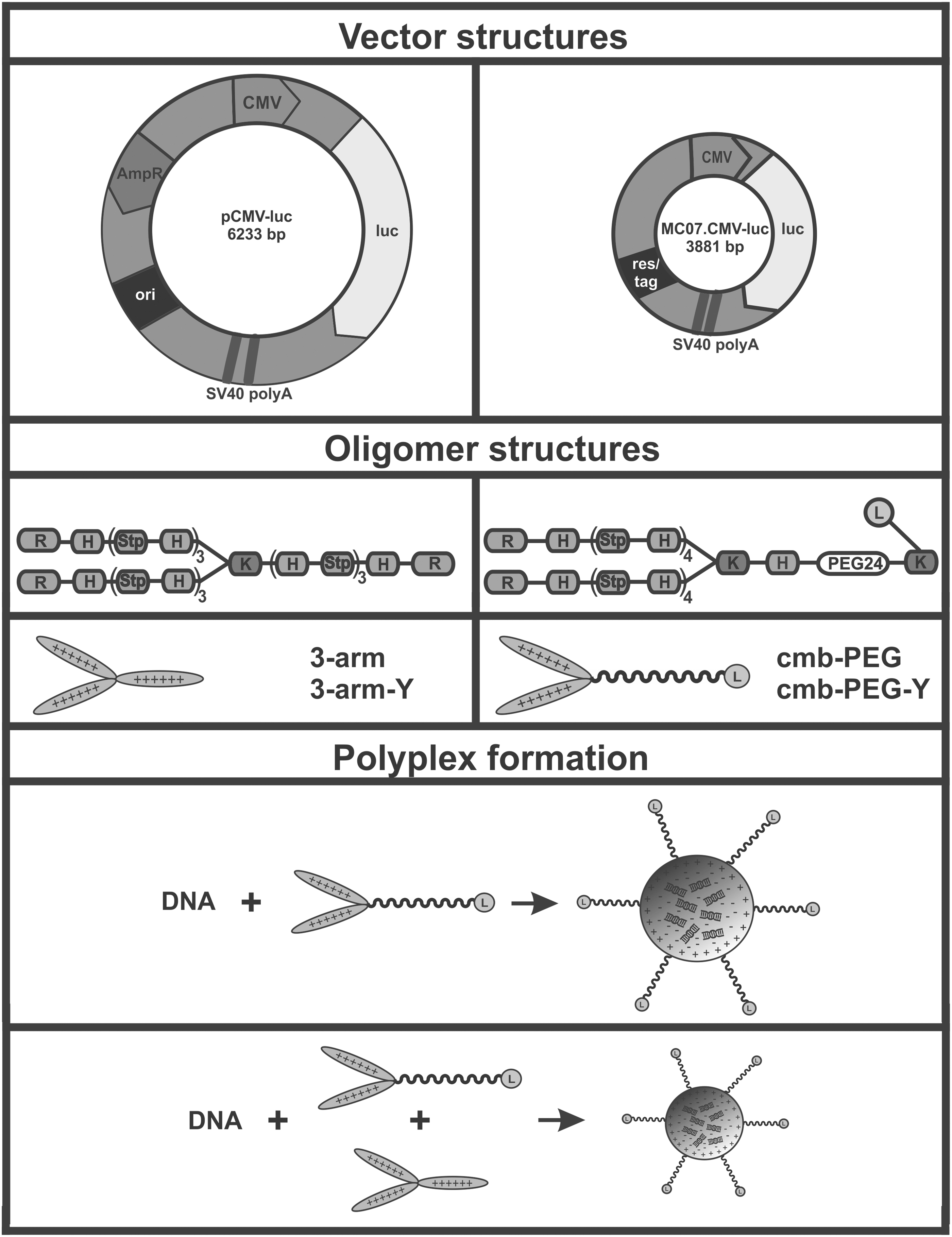
Schematic structures of
For formulation of DNA, linPEI was used as established and effective standard polymeric carrier. 26 –28 Four sequence-defined cationic oligoaminoamides were generated (Fig. 1B and Table S1) by solid-phase assisted synthesis using Fmoc-based peptide synthesis technology. Repeats of the artificial amino acid succinoyl-tetraethylene-pentamine (Stp) alternating with natural histidines served as the central cores for nucleic acid complexation and endosomal buffering function. Terminal cysteines provide disulfide-mediated stabilization of DNA polyplexes. Lysine residues were integrated as branching units, providing non-targeted carriers with three-arm topology (3-arm, 3-arm-Y; see Fig. 1B, left). Alternatively, one cationic arm was replaced by polyethylene glycol PEG24 and the c-Met targeting peptide cmb 69 –71 (cmb-PEG, cmb-PEG-Y; see Fig. 1B, right). PEG24 (containing a precise chain length of 24 oxyethylene units) was chosen to shield polyplexes against unspecific interactions, 34,68 and the cmb-ligand to trigger receptor-mediated endocytosis, as recently demonstrated. 34,70,72 Oligomers three-arm (689) and cmb-PEG (442) were previously described. 34 The additional incorporation of tyrosine trimers (Y3) had been found advantageous for aromatic and hydrophobic polyplex stabilization. 32,36 Therefore, in the current study, the Y3-containing analogous carriers, three-arm-Y (849) and cmb-PEG-Y (852), were also synthesized. Analytical data for these novel tyrosine trimer containing oligomers (characterized by 1 H-NMR and RP-HPLC) can be found in Supplementary Figs. S5–S8.
Efficient DNA compaction (Fig. 1C) is an important requirement for gene transfer. Recent work from the authors' lab revealed suboptimal pDNA packaging by cmb-PEG; apparently, the PEG shielding domain interfered with DNA condensation (Fig. 1C, top). 34,68 The PEG-free three-arm oligomer and also the cmb-PEG/three-arm (7:3 mol/mol) carrier combination (Fig. 1C, bottom) effectively compacted DNA, resulting in effective pDNA gene transfer in vitro and in vivo. 34 For this reason, combination polyplexes were also analyzed in the current comparison of pDNA and MC DNA polyplexes.
Physicochemical characterization of polyplexes
Nucleic acid compaction with synthetic gene carriers into nanosized particles is an important requirement for successful gene delivery. This study compared pCMV-luc and MC07.CMV-luc polyplex formation using a series of different characterizations. Particle sizes and zeta potential of polyplexes formed at N/P ratio 12 were determined by dynamic light scattering (Table 1).
Particle size (Z-average) and zeta potential of pCMV-luc and MC07.CMV-luc polyplexes formed in HBG buffer determined with DLS
Variations refer to the standard deviation of three measurements of the same sample. Combination polyplexes refer to 70 mol% of cmb-targeted oligomer and 30 mol% of three-arm oligomer.
PDI, polydispersity index.
The majority of carriers formed nanoparticles of <200 nm, within the size limit for cellular uptake via receptor-mediated endocytosis. 11 Zeta potential measurements display the expected positive surface charges of +∼20 mV for the nonshielded linPEI and Y3 oligomer, and surface charges of +∼14 mV for cmb-PEG decorated polyplexes, consistent with published work. 34 Comparing complexation with pDNA or MC, well-compacting cationic polymers such as linPEI or Y3 oligomers formed polyplexes with similar diameters, irrespective of the different size of the cargos; linPEI polyplexes had a hydrodynamic diameter of ∼90 nm. This is not surprising and is in agreement with previous work testing various nucleic acid cargos. 5 For plain polycations, the different cargo DNA size is compensated by aggregating different cargo copy numbers into one polyplex. The situation is different for carriers that avoid nano-aggregation and form monomolecular DNA polyplexes. 73,74
In the current work, in contrast to linPEI, the tyrosine-free oligoaminoamides form polyplexes where sizes significantly differ for pDNA and MC. Especially for cmb-PEG with lower compaction activity, hydrodynamic diameters of MC polyplexes were only ∼55% of the pDNA polyplexes. These findings are consistent with a previously established decorated rod model. 74,75 Mixing of cmb-PEG with PEG-free three-arm results in combination polyplexes with far less difference between pDNA and MC (170 nm vs. 140 nm diameter).
Differences in size and morphologies of the more stable polyplexes (with linPEI and tyrosine-oligomers) were analyzed in more detail by transmission electron microscopy (Fig. 2). Considering the methodological differences, TEM data are well consistent with DLS. In general, nominal sizes of hydrodynamic diameters by DLS (∼90–140 nm) are larger than TEM diameters (∼30–100 nm) due to the emphasis of DLS on larger particle fractions (light scatter intensity ∼r 6 ) on the one hand, and the dehydrated stage of TEM samples on the other hand. TEM confirmed stabile particles formation for all carriers, with nanoparticle shapes differing between the carriers but not the DNA used in polyplex formation. In contrast to DLS, TEM provided evidence for the presence of smaller ultrastructures for MC than for pDNA polyplexes. For example, linPEI formed spherical structures with both DNA vectors, with sizes of 50–90 nm for pDNA and smaller 25–70 nm for MC. The addition of a three-arm structure to the cmb-targeted structure led to a smaller and more compact particle in the case of polyplex formation with both plasmids. In accordance with a recent observations, 34 the PEG-free three-arm-Y structure formed compact spherical structures of around 30–50 nm. In contrast, oligomer cmb-PEG-Y formed less compact rods (80–100 nm for pDNA/50–60 nm for MC) or toroids (40–50 nm for pDNA/30 nm for MC) demonstrating a DNA cargo size-dependence. As expected, the oligomer combination resulted in better compacted rod structures, with a clear size difference between pDNA and MC formed polyplexes (65–100 nm vs. 35–40 nm). Notably, the majority of nanoparticles were <50 nm (at least in one dimension), which would be compatible with the <50 nm nuclear envelope pore limit to enable transport of polyplexes into the nucleus. 22

Transmission electron microscopy (TEM) images of pCMV-luc and MC07.CMV-luc polyplexes formed at N/P 12 in deionized water and stained with uranylformate. Combination polyplexes refer to 70 mol% of cmb-PEG-Y and 30% of three-arm-Y. Scale bar: 200 nm.
The EtBr exclusion assay (Fig. 3) provides an alternative mode for evaluation of DNA binding ability of carriers. The decreased fluorescence correlates with an increased compaction of polyplexes (Fig. 3A). Compaction was best for linPEI polyplexes, irrespective of the type of DNA (<5% EtBr fluorescence). Interestingly, polyplexes formed with oligoaminoamides displayed better compaction for MC compared to pDNA. Consistent with the other presented work, MC DNA combination polyplexes showed even better compaction (only 6–8% EtBr fluorescence). Figure 3B presents the EtBr fluorescence after the addition of 250 IU of heparin sulfate to mimic physiological anionic dissociative stress, resulting in partial release of DNA from polyplexes. 76 Under these conditions, both pDNA and MC polyplexes of linPEI are dissociated (>90% EtBr fluorescence). Oligoaminoamide polyplexes, which are stabilized by bioreversible disulfide cross-links, are more resistant to the applied heparin stress. Again, polyplexes formed with MC DNA display higher stability than pDNA polyplexes, and MC combination polyplexes the highest stability (∼25% EtBr fluorescence).
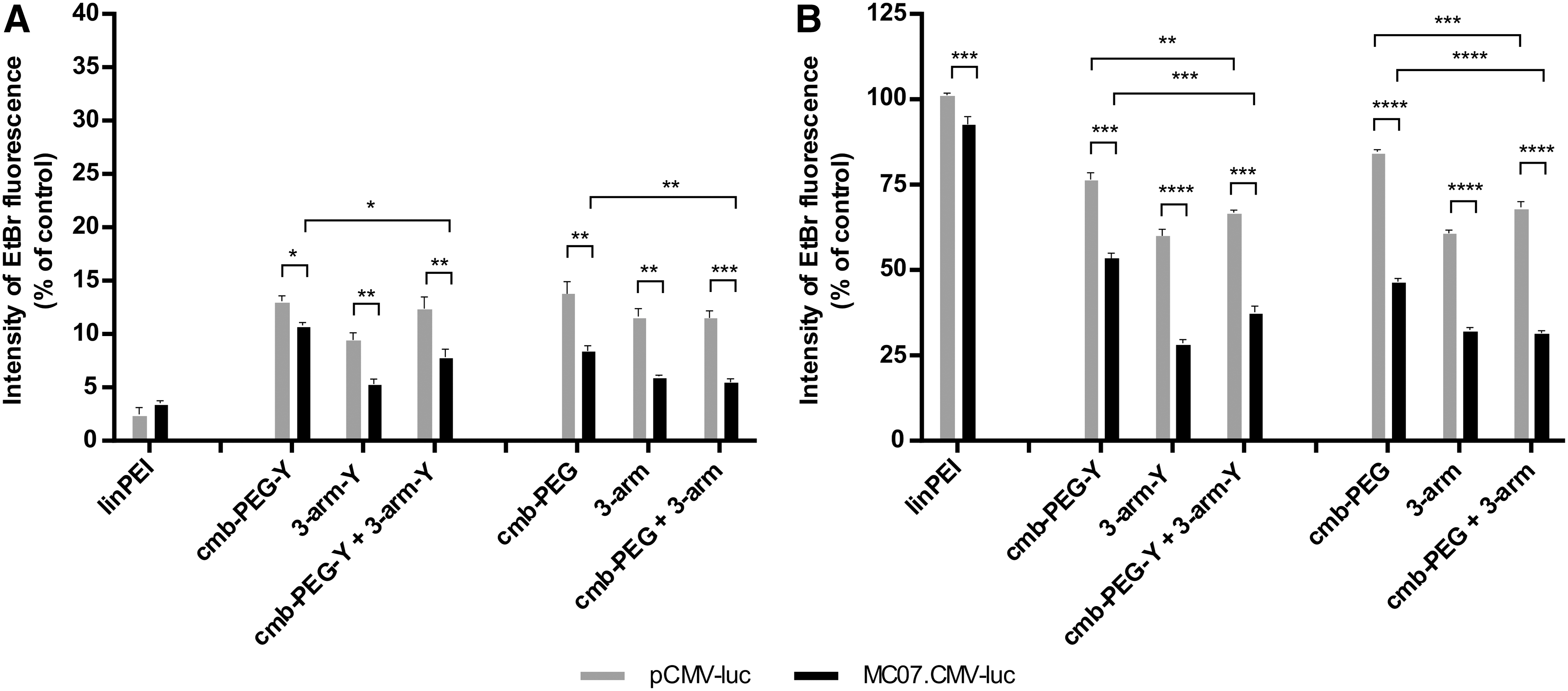
Ethidium bromide exclusion assay to determine the ability of DNA compaction of the oligomers, inversely correlating with the remaining fluorescence of DNA intercalating ethidium bromide (EtBr). Results are presented in % against free pCMV-luc or MC07.CMV-luc. Polyplexes at N/P 12 without
Transfection efficiency of polyplexes formed with pCMV-luc or MC07.CMV-luc
To compare transfection efficiency of polyplexes formed with either pCMV-luc or MC07.CMV-luc, luciferase gene transfer in the prostate carcinoma cell line DU145 was performed. This cancer cell line expresses the cell surface receptor HGFR/c-Met (see Supplementary Fig. S5). First, cells were incubated with polyplexes for a defined period in complete serum-supplemented medium, and luciferase gene expression (Fig. 4, Supplementary Fig. S6) as well as metabolic activity of cells (Supplementary Fig. S6 and S7) was recorded after 24 h. Metabolic activity assays confirmed that the transfection were performed under nontoxic conditions. As shown in Fig. 4A, polyplexes formed with MC07.CMV-luc and linPEI mediated better transfection than polyplexes formed with pCMV-luc. The advantage was particularly pronounced (fivefold higher transfection) in short-term (0.75 h) incubations with polyplexes, but still visible (2.3-fold higher transfection) in 24 h long-term incubations. For pCMV-luc polyplexes, this prolonged incubation was necessary to reach the same transgene expression levels as obtained with MC07.CMV-luc polyplexes after short incubation. Also, for three-arm-Y polyplexes, upon 24 h of incubation, an advantageous gene transfer by MC compared to pDNA was observed (Fig. 4B). For receptor-mediated transfections, cells were incubated with cmb-PEG or cmb-PEG-Y polyplexes for only 0.75 h. Previous work demonstrated enhanced, HGF receptor-specific uptake at these short incubation times. 34 Additionally, the corresponding combination polyplexes (targeted oligomers plus three-arm oligomers) were applied (Fig. 4C). Also, in these settings, polyplexes formed with MC07.CMV-luc showed 6- to 13-fold higher transfection efficiency than polyplexes formed with pCMV-luc. An advantageous effect of the novel c-Met targeted oligomers with integrated tyrosine trimers analogously was also noted, as previously observed for folate receptor targeted pDNA polyplexes. 32, 36 Altogether, the beneficial effect of tyrosine trimer integration, formation of combination polyplexes, and use of MC added up to a ∼200-fold enhanced gene expression.
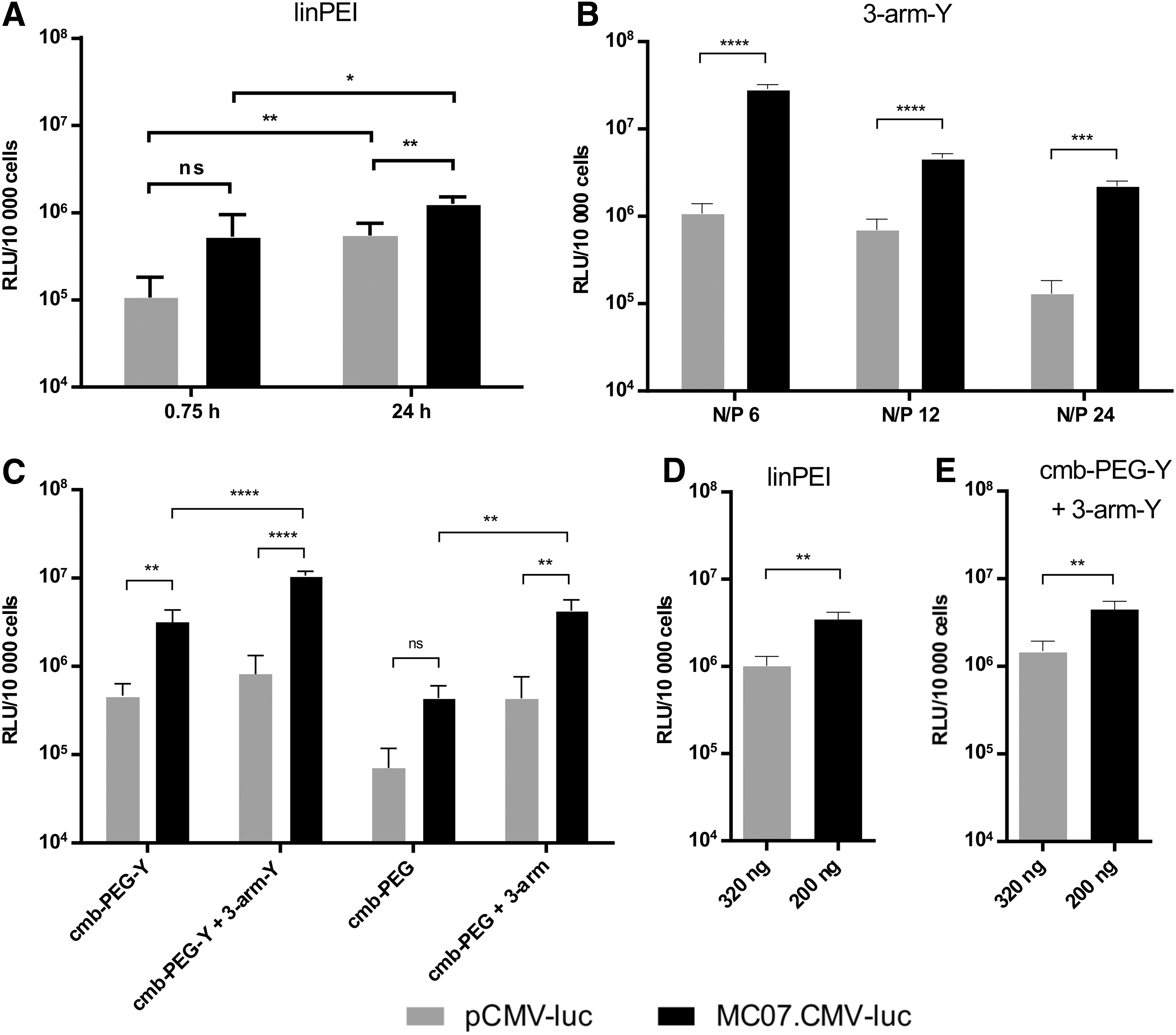
Luciferase reporter gene expression in the DU145 prostate carcinoma cell line at 24 h after transfection with pCMV-luc (gray) or MC07.CMV-luc (black) polyplexes.
In the presented transfections, polyplexes with equal mass quantities of DNA were used. To rule out that the higher copy number of MC07.CMV-luc than the corresponding pCMV-luc vector is responsible for the enhanced transfection, additional polyplex transfections were performed with equimolar quantities of vectors. Under these conditions, linPEI and combination polyplexes formed with MC also showed around 3.5- and 3-fold higher transfection efficiency, respectively, in comparison to equimolar quantities of pDNA polyplexes (Fig. 4D and E).
Overcoming intracellular barriers
The advantage of MC DNA in polyplex transfection, although confirmed in the current and other previous work, is not easy to understand. The clear-cut differences in physicochemical properties, such as smaller and more compacted nanostructures of MC DNA polyplexes, might favorably or unfavorably translate in the complex, multi-step process of non-viral gene delivery (see Fig. 5). 77 For electroporation of free DNA, MC DNA presented a more effective vector uptake. 47 Maucksch et al. 78 also reported that a small pDNA size can be important for cell and nucleus entry upon electroporation; comparing 4.7 kb of pEGFP-monomer with 9.4 kb of pEGFP-dimer pDNA, a 17% versus 13% nuclear delivery was observed.
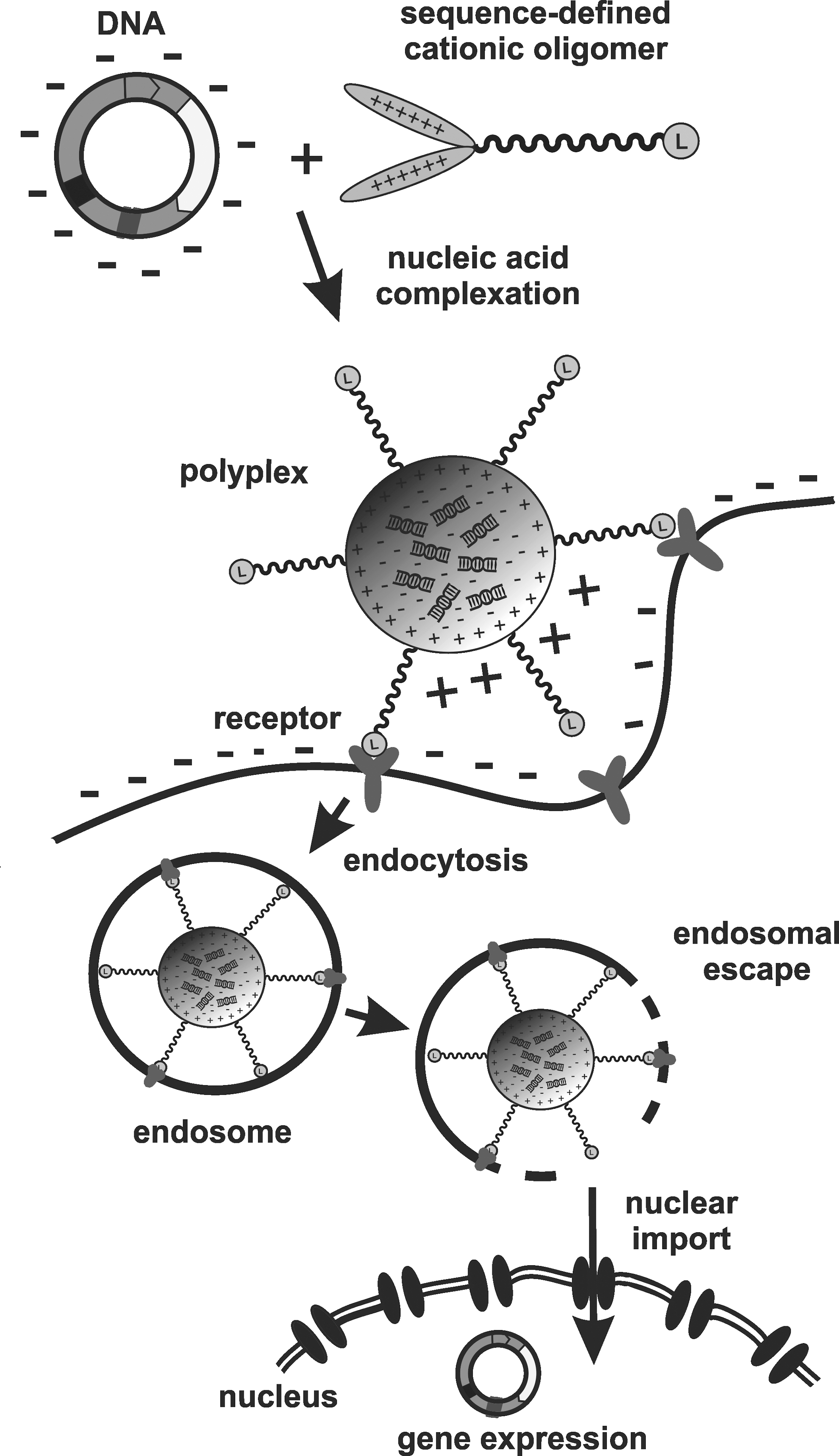
Schematic presentation of the gene delivery process of polyplexes formed from DNA and sequence-defined cationic oligomer.
For polyplexes, which usually package multiple vector molecules, nanoparticle size may have more impact for initial cellular uptake than vector size. On the one hand, larger PEI polyplexes were previously found to possess increased transfection efficiency due to their enhanced cellular contact by sedimentation in cell culture, subsequent uptake, and enhanced endosomal escape activity by the proton sponge effect. 79 On the other hand, smaller nanoparticle sizes may provide favorable characteristics in cellular uptake by mechanisms of receptor-mediated endocytosis, and favorable subsequent intracellular sorting pathways avoiding lysosomal destruction. Intuitively, by their smaller size, they might have advantages in sneaking across intracellular barriers. In particular, the intranuclear delivery of the vector presents a crucial size-dependent process, followed by unpackaging and successful gene expression in the nucleus. 25 Recent work suggests that DNA/PEI with a diameter <50 nm would pass through nuclear envelope pores. 22 Larger pDNA nanoparticles might enter the nucleus during the small window of mitosis, when the nuclear membrane breaks down. Smaller MC nanoparticles might have the advantage to enter the nucleus at any stage of cell cycle.
Therefore, transfections using DU145 cells were performed in different stages of cell cycle (Fig. 6). For synchronization, a double-thymidine block was used. 80,81 The cell cycle stage was monitored by flow cytometry (see Supplementary Fig. S8). Upon second release (0 h), 88% of cells were in G1 phase, at 6 h after release 45% of cells peaking in S phase, and at 8 h 60% of cells in G2/M phase. Cells were transfected at time points 0 h (G1), 6 h (largely S), 8 h (G2/M), and 12 h (after M). Three different formulations, the well-compacting carriers linPEI and the cmb-PEG-Y plus three-arm-Y combination, and the less compacted cmb-PEG-Y, were selected for packaging pDNA or MC DNA. Irrespective of cell cycle stage, the MC formulations mediated up to a 10-fold higher gene expression (Fig. 6). The transfections confirmed the lack of significant cell cycle dependence for linPEI/pDNA polyplexes, which correlates with previous studies. 18 An absence of cell cycle dependence was also observed for linPEI/MC polyplexes. An analogous lack of cell cycle dependence was found for both types (pDNA and MC) of well-compacted cmb-PEG-Y/three-arm-Y combination polyplexes. Thus, from these experiments, a hypothetically improved nuclear entry of smaller MC polyplexes over pDNA polyplexes can be neither verified nor excluded. It rather appears that nuclear import is a low barrier for both types of small compacted polyplexes. Interestingly, for the less compacting carrier cmb-PEG-Y, the pDNA- but not the MC-mediated transfection shows cell cycle dependence at 12 h (after mitosis), which is about 10-fold lower than for transfection before mitosis (0 or 6 h), and also about 10-fold lower than the better compacted pDNA polyplex (Fig. 6, top left). Importantly, MC DNA polyplexes (Fig. 6, top right) display lack of cell cycle dependence for all formulations, suggesting an advantage from their inherent smaller size.
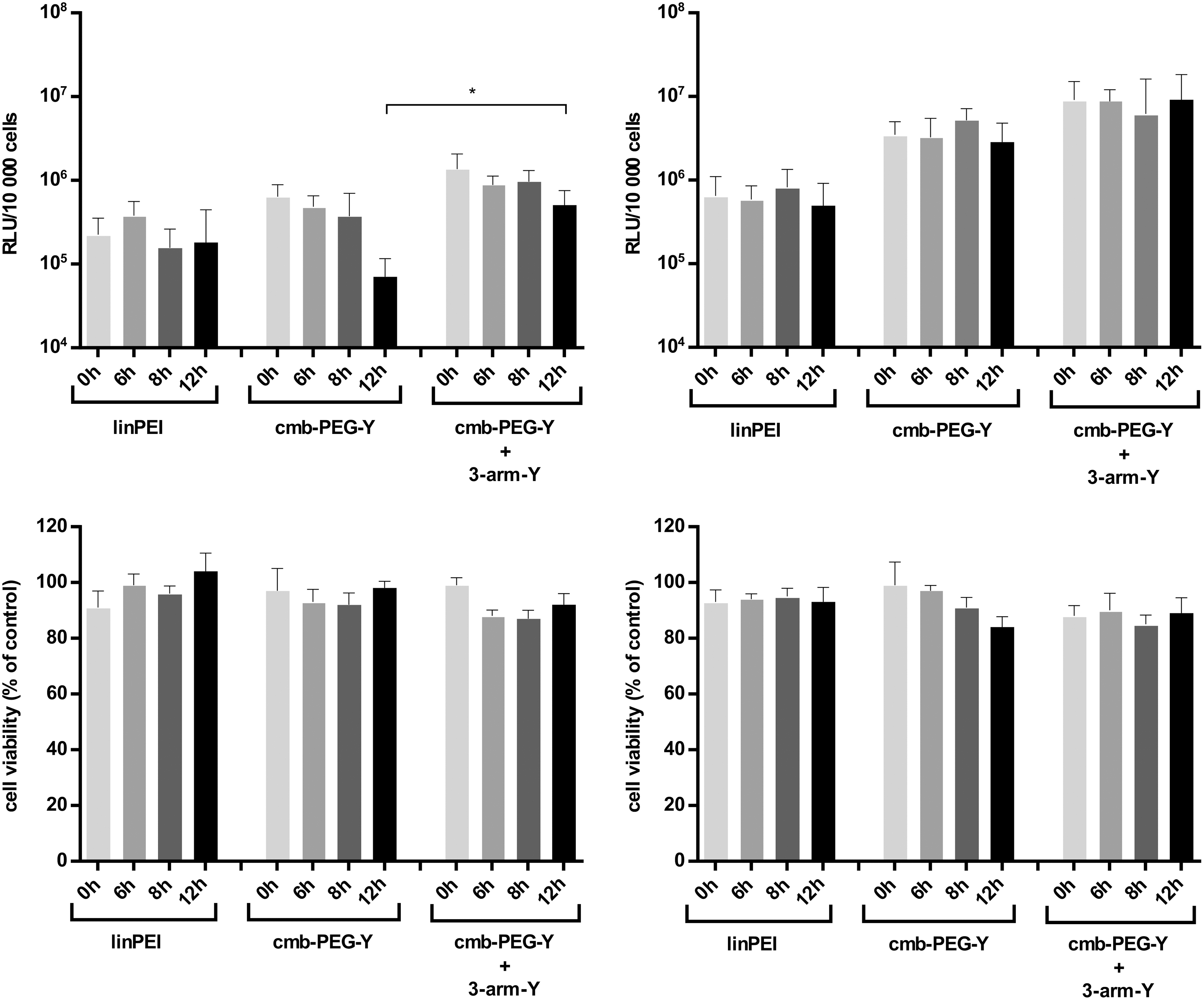
Cell cycle dependence of gene transfer. DU145 cells were transfected in specific phases of cell cycle after release from double thymidine block; primarily G1 phase (0 h), S phase (6 h), G2 phase (8 h), and G1 phase of the second cycle (12 h). Luciferase reporter gene expression (top) and corresponding cell viability by MTT assay (bottom) at 24 h after 0.75 h of incubation with pCMV-luc (left) or MC07.CMV-luc polyplexes (right) at N/P ratio 12.
MC DNA comprises several additional advantages beyond the aspects discussed above. In particular, for in vivo gene transfer, reduced innate immune responses and strongly improved maintenance of transgene expression are noteworthy. 44,47,48 This favorable sustained expression appears to be linked with incorporation of MC DNA into active chromatin. 50 Following in vivo gene transfer of standard pDNA into the liver of mice, the presence of the bacterial backbone resulted in a 10- to 1,000-fold reduction of gene expression over a period of few weeks due to gene silencing at the nuclear transcription stage. Apparently, DNA elements in cis (directly linked with the transgene expression cassette) are responsible for transcriptional blockade; the in vivo excision of the transgene cassette from such elements strongly improved maintenance of gene expression. 82 Interestingly, the size (≥1 kb) of the extragenic DNA had more impact than the CpG content, 83,84 though the increase of A/T content in the backbone or antibiotic resistance genes reduced transcriptional silencing. 85
In conclusion, the work presented here demonstrates better physicochemical characteristics and transfection efficiencies of MC DNA polyplexes compared to standard pDNA with linPEI or sequence-defined oligoaminoamides. Within the latter class of carriers, novel c-Met targeted and tyrosine trimer-stabilized oligomers were designed, which by optimized formulation in combination with MC DNA yielded more than 100-fold enhanced gene transfer efficiency in receptor-positive target cells. These results provide an encouraging future perspective for targeted in vivo gene delivery.
Footnotes
Acknowledgments
We are grateful to Marco Schmeer, PlasmidFactory GmbH (Bielefeld, Germany), for helpful discussions. We also thank Olga Brück for secretarial assistance and Wolfgang Rödl for technical support. This work was supported by the Deutsche Forschungsgemeinschaft (DFG) Cluster of Excellence Nanosystems Initiative Munich (NIM), SFB1032 (project B 04), SFB1066 (project B 05), and the Sino-German Center research grant GZ995 (all to E.W.).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.