
Abstract
Adenovirus-based vectors comprise the most frequently used vector type in clinical studies to date. Both intense lab research and insights from the clinical trials reveal the importance of a comprehensive understanding of vector–host interactions. Especially for systemic intravenous adenovirus vector delivery, it is paramount to develop safe and efficacious vectors. Very early vector–host interactions that take place in blood long before the first cell is being transduced are phenomena triggered by the surface, shape, and size of the adenovirus vector particles. Not surprisingly, a multitude of different technologies ranging from genetics to chemistry has been developed to alter the adenovirus vector surface. In this review, we discuss the most important technologies and evaluate them for their suitability to overcome hurdles imposed by early vector–host interactions.
Introduction
A
Adenovirus Biology and Capsid Structure
Adenoviruses, members of the family Adenoviridae, are nonenveloped DNA viruses. So far more than 70 types of human-infectious adenoviruses have been identified. Based on genome structure, hemaglutination properties, and sequencing results, these types are divided into seven species (human adenovirus A–G), 6,7 with Ad species B, C, and D comprising the types being most commonly used in gene therapy. The human Ad is a 38-kb double-stranded DNA virus encapsulated by an icosahedral nucleocapsid. 8 This capsid consists of nine proteins of which hexon, penton base, and fiber are referred to as major capsid proteins (Fig. 1). With 720 monomers, hexon is not only the largest, but also the most abundant capsid protein, forming 20 capsid facets each consisting of 12 hexon homotrimers. 9,10 Homopentamers of penton base protein are located on each icosahedral edge (vertex) forming part of the penton and representing the base for the vertex's spike that consists of trimers of glycolysated fiber protein. 10,11 Ad entry is composed of two major steps. Step one involves attachment of the fiber knob to its primary receptor. For Ad types from species A, C, E, and F, this primary receptor is the coxsackie and adenovirus receptor (CAR), while Ad types from species B and D were shown to interact with receptors distinct from CAR. This interaction brings virion and cell surface in spatial proximity to each other facilitating the interaction of cellular integrins with the Arg-Gly-Asp (RGD) tripeptide in penton base. In a second step, this interaction induces cellular responses that change the cytoskeleton, internalize the virion via clathrin-coated vesicles, and transport to the endosome. 12 In the endosome, the Ad virion partially disassembles and is released to the cytoplasm, where it ultimately travels to the nucleus for viral replication.
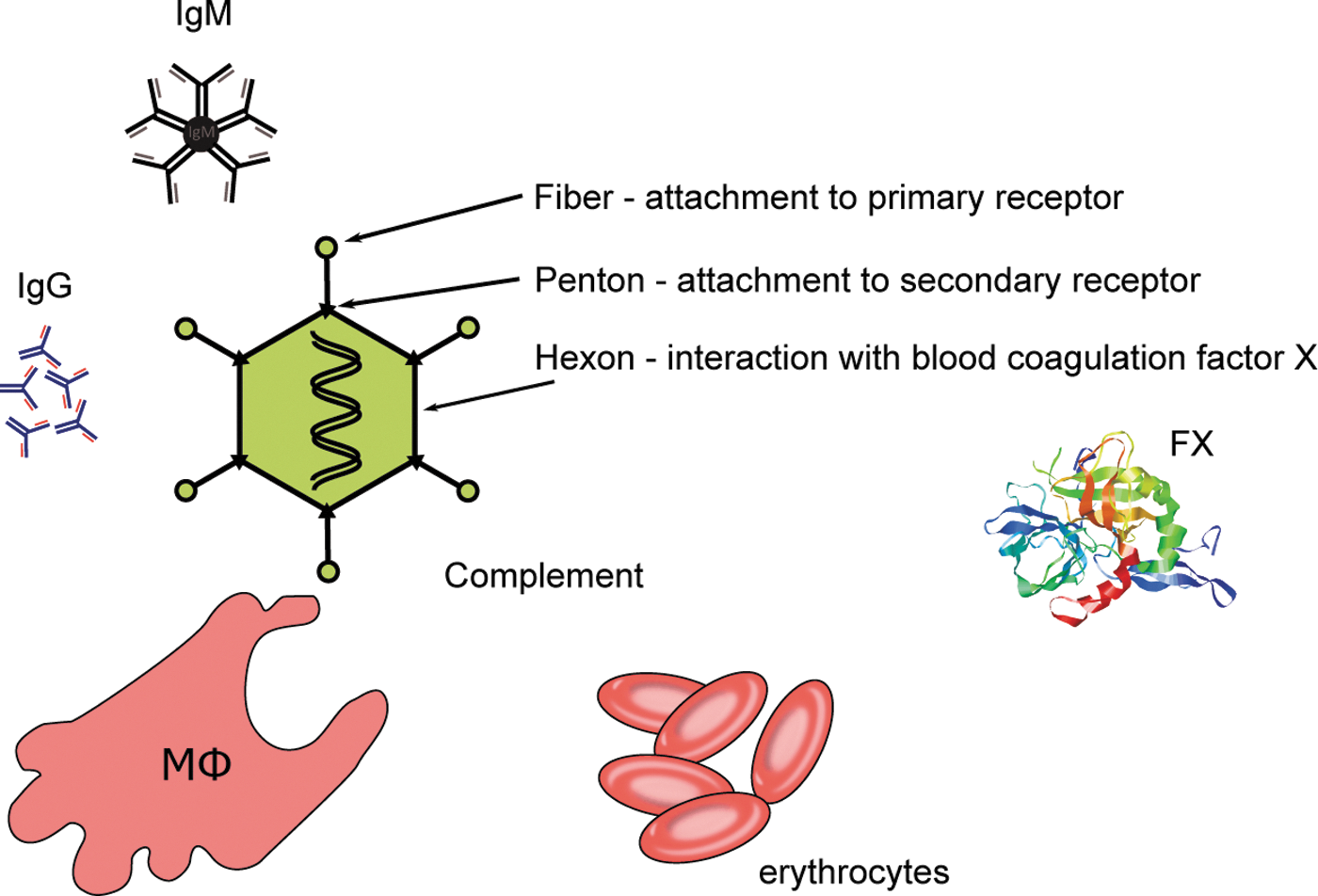
Adenovirus capsids and barriers for delivery. Fiber (36 copies per particle), Penton (60 copies per particle), and Hexon (720 copies per particle) are the most important capsid proteins involved in adenovirus vector sequestration and mistargeting by various barriers. FX, blood coagulation factor X; IgG, immunoglobulin G; IgM, immunoglobulin M; MΦ, macrophages. (Not drawn to scale.)
Successful application of Ad vectors as transfer vehicles requires efficient transduction of the respective targets cells without affecting non target cells. The utilization of tissue-specific promoters that are active in target cells only (transcriptional targeting, reviewed by Sadeghi and Hitt 13 ), represents one approach. While potential toxic side effects of transgene expression can be reduced or even eliminated, this approach is still prone to side effects that may result from mislocalization of vector particles. However, irrespective of replication competence and infectivity, viral capsid proteins themselves have toxic side effects provoking an innate immune response with systemic release of cytokine interleukin-6. 14 Using transcriptional targeting as a single strategy, the efficient transduction of the target cell or tissue is not sufficiently addressed. One major obstacle is that the native tropism of the virus rarely meets the therapeutic need. Hence, efficient transduction of the correct cells often requires ablation of the native tropism and/or modification of the vector to infect cells that are naturally not a target (transductional targeting). Based on the knowledge about native Ad entry mechanisms, non-CAR-expressing cells (e.g., many cancer cells) are predicted to be refractory to Ad transduction. In this scenario, high-CAR-expressing non target cells are efficiently transduced while low-CAR-expressing target cells would show poor transduction. However, in vivo the biodistribution appeared to be independent of CAR expression profiles. 15 Upon intravenous Ad delivery in the mouse the majority of transduction occurred in the liver, followed by spleen, heart, lung, and kidney; this transduction profile, however, does not correlate with CAR expression levels. 16 Likewise, ablating CAR and integrin binding in Ad capsids had only a small effect on virus uptake in the liver. 17,18 A major role of Ad fiber in virion interactions with blood coagulation factors was reported. Modification of Ad5 fiber preventing its interaction with FIX and C4BP resulted in a distinct decrease in liver transduction. 4 In other studies, a specific interaction between FX and hexon hypervariable regions (HVR) 3, 5, and 7 was defined. This interaction could be inhibited by either pharmacological factors or mutated forms of hexon. 19,20
Capsid Modifications
As mentioned above, most hurdles that are currently faced in Ad vector development arise from interactions with viral capsid proteins upon intravenous delivery, and numerous vector–host interactions influencing Ad tropism have been described in the past years. These new insights pave the way for promising modifications of the Ad capsid that are summarized in Table 1. In the following we will focus on genetic and chemical modifications of the Ad capsid. Figure 2 provides a schematic overview of the different technologies.
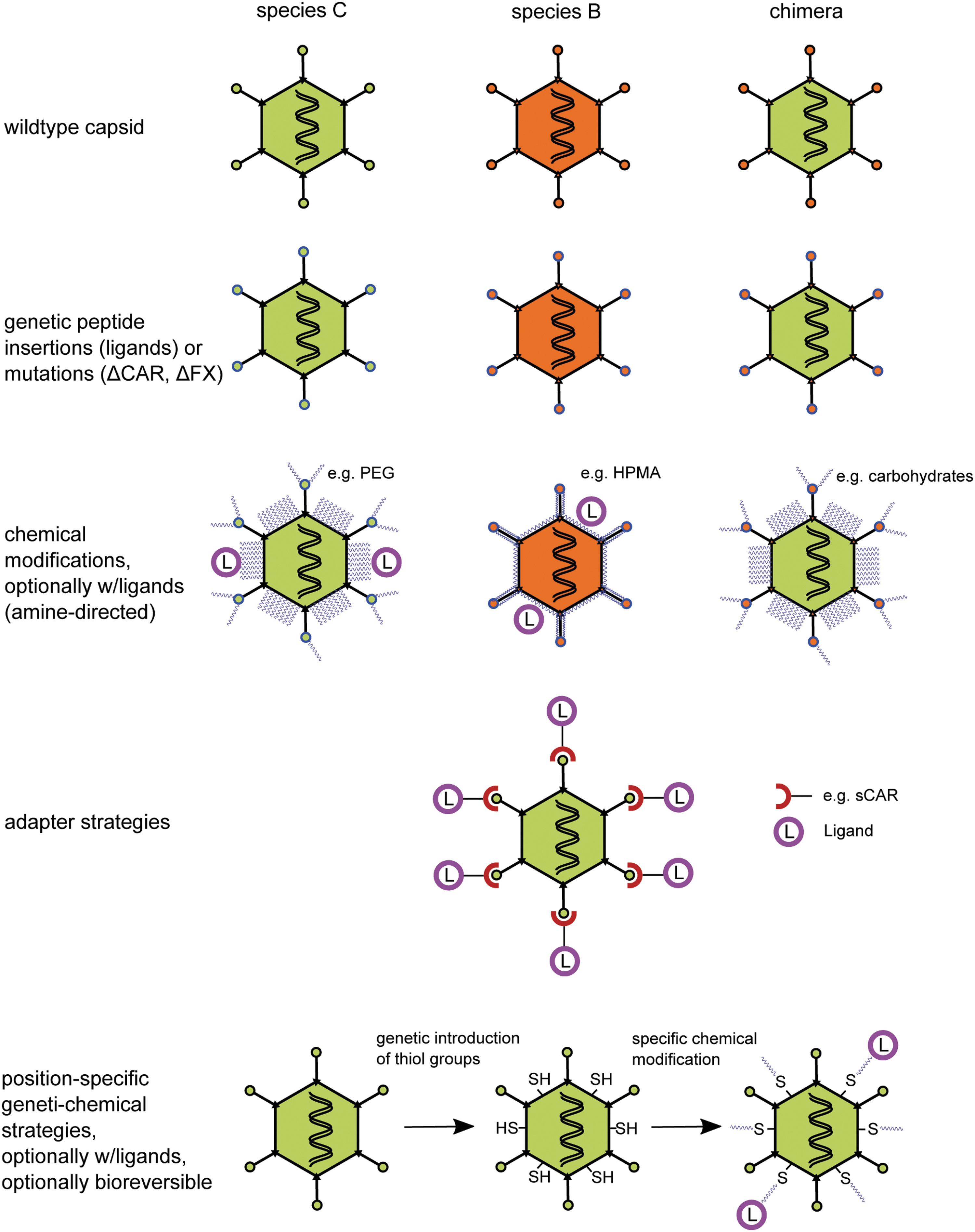
Schematic overview of capsid modifications technologies to improve delivery. To improve delivery by adenoviral vectors, multiple capsid modifications technologies exist. Chimeric adenovirus (Ad) particles can be generated by swapping entire capsid proteins or domains (top panel, wildtype capsid). Small genetic modifications can be employed to introduce peptidic ligands and to ablate coxsackie and adenovirus receptor (CAR)-binding or blood coagulation factor X (FX) binding. Such modifications have also been performed for types other than Ad5 and chimeric vectors (second panel, genetic peptide insertions). Amine-directed chemical modifications can be used to shield the vector capsids by synthetic polymers like polyethylene glycol (PEG) or poly-N-hydroxypropylamid (HPMA). In addition, ligands (L) can be coupled to the polymer shield. Chemical modifications are feasible with types other than Ad5 and chimeric vectors (third panel, chemical modifications). In contrast with chemical approaches, adapter strategies rely on noncovalent attachment of ligands via bifunctional adapters. Shown is sCAR, the soluble extracellular domain of the Ad receptor CAR that can be fused to ligands (fourth panel, adapter strategies). Geneti-chemical modifications combine small genetic modifications with position-specific, thiol-directed chemical modifications. The advantage is a precise shield instead of a dense cloaking. This way the infectivity of the particles can easily be maintained and ligands can be attached at advantageous positions (lower panel, position-specific genti-chemical strategies).
Overview of possible adenovirus capsid modification strategies
The table summarizes advantages and disadvantages of different capsid modification strategies, outlining specific obstacles that can be overcome.
Ad, adenovirus.
Chimeric capsids and directed evolution
Modifying vector tropism via genetic alteration of the Ad capsid represents the most direct approach. While the native entry biology of the commonly used Ad5 is based on a fiber–CAR interaction, primary attachment of other Ad species is CAR-independent. Accordingly, several attempts to modify Ad tropism focused on generation of chimeric Ad vectors. In this approach, parts of or the complete fiber are genetically substituted with its counterpart of other species resulting in altered tropisms, for example, enhanced transduction of malignant glioma. 21,22 The new types are preferentially generated from species B and D. Species B viruses have been shown to interact with receptors such as CD46, 23 CD80, and CD86, 24 or desmoglein 2 (types Ad3, 7, 11, 14), 25 while viruses of species D interact with CD46 and the glycoprotein component α(2-3)-linked sialic acid. 26 Interestingly, chimeric Ad vectors with fibers from species B Ad7 and Ad35 displayed an altered intracellular trafficking pathway. 27,28 These chimera resided longer in the endosome. While some virions eventually travelled to the perinuclear lysosome, others were recycled back to the cell surface 28 . In vivo, a chimeric, CAR and integrin-ablated Ad5 derived from Ad35 or Ad40 showed decreased liver transduction. 29,30 Additionally, chimeric Ad vectors were not only constructed for transductional targeting purposes, but also to evade anti-Ad5 immunity.
The high immunogenicity and high titers of Ad5-neutralizing antibodies in the human population indeed represents an important hurdle that needs to be overcome. Since the neutralizing antibodies are primarily directed against the HVRs of hexon, replacing HVRs of Ad5 with those of a less prevalent type (e.g., Ad48) 31 was one successful strategy. Replacing all seven HVRs of Ad5 with their counterparts from Ad48 evaded the majority of preexisting Ad5 immunity in vivo. 32 Although some studies postulate that the majority of anti-Ad5 antibodies is directed against HVR1 and HVR5, 33,34 partial HVR-chimeric Ad5 vectors with only a subset of HVR substituted only insufficiently evaded preexisting Ad5-specific immunity in mice. 35 These data indicate that swapping all HVRs of Ad5 is necessary to evade neutralising antibodies. Such a vector has been evaluated recently as a human immunodeficiency virus (HIV) vaccine vector in phase 1 clinical trial. 36 However, Ad5:Ad48 chimeric vectors intravenously injected in mice triggered a robust inflammatory response, 37 with serum cytokine and chemokine responses elicited by Ad5:Ad48 being higher than those elicited by Ad5. 38
In an approach termed “directed evolution,” Kuhn et al. (2008) passaged a pool of seven Ad types (from species B–F) on cancer cell lines to invite recombination. 39 Subsequently, potent Ad variants or types showing early signs of cytopathic effects were selected. In this process a novel chimeric Ad3/Ad11p oncolytic virus, ColoAd1, was generated. ColoAd1 displayed enhanced potency (9- to 100-fold over Ad5) on various colon tumor cell lines and in an intravenous liver tumor model. Relative to the parent Ad11p, ColoAd1 displayed a chimeric region in E2B and deletions in E3 and E4. 39 Interestingly, recombination did not occur in late genes encoding for viral capsid proteins. Since the seroprevalence of Ad11p is rather low, 40 ColoAd1 represents an attractive approach to systemically treat colon cancer and is currently undergoing a series of early phase clinical trials.
Capsid modifications with genetically inserted peptides or protein domains
A different approach uses short peptides inserted in the HI-loop, an exposed loop connecting β-sheets H and I, or C-terminus of fiber knob domain for Ad vector targeting. In principle, the C-terminus is the ideal location for peptide insertion. Linking an integrin-binding RGD-motif or heparin sulphate binding motif (pK7) in the C-terminus resulted in increased transduction of fibroblast and smooth muscle cells expressing alpha(v)integrin and cells lacking high levels of CAR expression, respectively. 41 However, since the interaction of fiber with CAR is not inhibited by insertions of short peptides, Ad vectors with expanded tropism are created. When combining both motifs in Ad fiber, RGD motif in HI-loop and pK7-motif in C-terminus, increased transduction efficiencies compared with nontargeted and singly modified Ad vectors in both CAR-positive and CAR-negative cells was shown. Moreover, incorporation of both motifs in Ad fiber knob resulted in a loss of its natural CAR-affinity. 42 In further studies it was shown that fiber knob can accept up to 89 amino acids without a loss or impairment of function. Specifically, the HI-loop was shown to accept an RGD-containing sequence of penton base of 83 amino acids in length, 43 while the C-terminus was fused to a 89 amino acid linker and biotin acceptor peptide (BAP), 44 demonstrating the applicability of adapter strategies at fiber knob. Besides tropism-expanding peptides, cell-specific peptides were also incorporated: A vascular-endothelial cell targeting peptide was shown to enhance Ad infectivity for a variety of cancer cell lines. 45 Insertion of an Affibody, an antibody mimetic peptide, resulted in HER2/neu and Taq-polymerase-specific targeting. 46 However, besides fiber also other Ad capsid proteins gained attention due to their ability to display peptides. Protein IX, a polypeptide known to stabilize the capsid, is capable of displaying large polypeptides and proteins like EGFP with marginal effects on virion thermostability and bioactivity, 47 demonstrating that large proteins can also be linked to Ad capsid proteins. Another site for peptide insertion is the HVR5 surface loop of hexon. However, this site has been shown to accept only small insertions like RGD or His6, 48 –50 with the exception of a 71 amino acid BAP. 51 In a combinatory approach, Hesse et al. (2007) demonstrated that insertion of a 9 amino acid RGD peptide into one of the three loops of Ad41 short fiber knob (EG, HI, and IJ) was feasible without loss of fiber trimerization. 52 These successfully modified Ad41s fibers were then used to generate chimeric Ad5 vectors that exhibited increased transduction efficiencies in several cell lines. 52 However, despite some success incorporation of peptides is limited by a small number of motifs and therefore not suitable for specific targeting for all tissues. Moreover, there is no guarantee that any given peptide will be tolerated. Peptide insertion, although only 5–15 amino acids in length, may interfere with folding and/or multimerization of the modified capsid protein preventing vector rescue. It is also conceivable that peptides, although tolerated, lose their specificity in the structural context of the modified protein domain.
Adapter strategies
Another strategy for transductional targeting is based on the expression of single-component fusion proteins that serve as adapters. Fusing a single-chain antibody (scFv) against fiber knob to a scFv directed against epidermal growth factor (EGF) increased Ad gene transfer to EGF receptor–expressing cells. 53 Likewise, based on a truncated and soluble portion of CAR (sCAR) fused to a target molecule, a novel adapter system was developed. The first sCAR adapter, fused to EGF, transduced several EGF-overexpressing cancer cell lines with higher efficiency when compared with its untargeted counterpart. 54 Here the construction of trimerized sCAR fused to targeting ligands was shown to further improve the interaction of sCAR with fiber knob. 55 Other strategies fused sCAR to a scFv against polySia, a homopolymer that is frequently expressed on tumors of neuroendocrine origin, thus enabling effective targeting of oncolytic Ad to polySia expressing tumors. 56 The observation that some proteins like Tat (transactivator of transcription) of hHIV 57 or VP22 of herpes simplex virus 58 are taken up by mammalian cells via receptor-independent pathways led to the identification of protein transduction domains (PTD). PTDs are 10–30 amino acids in length and have been reported to deliver therapeutic proteins, 59 antisense oligonucleotides, 60 and plasmid DNA 61 into mammalian cells. Fusing PTDs to sCAR strongly increased affinity to and infection of CAR-negative nonpermissive cells. Moreover, when co-infected with conditionally replicating Ad vectors, uptake rate and lysis of permissive tumor cells was also increased. 62 Although adapter-based strategies provided vast evidence for retargeting of native Ad tropism, the preparations of these adapter molecules can be highly variable. The noncovalent nature of the adapter molecule–Ad capsid binding may limit its use for in vivo purposes and clinical translation as these interactions may be partially disrupted upon delivery. Further, endogenous CAR receptors or naturally occurring antibodies can compete for adapter binding thus circumventing vector targeting.
Chemical and genti-chemical capsid modifications
While the above described modification strategies are limited due to the noncovalent binding-nature and/or size of inserted or attached molecules, a modification approach originally used to modify therapeutic proteins has been adapted for Ad vectors. Polyethylene glycol (PEG) has been shown to reduce antigenicity and immunogenicity of therapeutic protein compounds. 63,64 Therefore, the covalent coupling of polymers to Ad capsid proteins opened new perspectives for transductional targeting and shielding strategies. These chemical modifications are performed after production and purification of the virus avoiding specific production cells. Covering Ad vector particles with polymers like PEG and poly-N-(2-hydroxypropyl)methacrylamid (pHPMA) shields the vectors from undesired vector–host-interactions. Due to the hydrophilic nature of covalently attached polymers, vector particles in solution are surrounded by a stable water shell that reduces amongst others enzymatic degradation and immune cell recognition. Commonly, polymer shielding targets ɛ-amine groups from lysine side groups that are randomly distributed on the capsid surface. PEGylation has been performed for the first time in 1999. In their work, O'Riordan et al. demonstrated that PEGylated Ad vectors evaded neutralization by purified anti-hexon antibodies in vitro and in pre-immunized mice in vivo. 65 Besides vector shielding, polymers can also be used to covalently attach ligands for targeting. Using polymer-incorporated ligands like FGF resulted in 6-fold higher transduction of skeletal muscle in mice. 66 While nonreactive ends of monovalent PEG molecules protrude from the capsid surface, multivalent pHPMA molecules are linked to a multiplicity of reactive sites coupling the polymer to the particle surface. Polymer shielding using pHPMA led to promising results in terms of evasion from neutralizing antibodies, 67 binding to blood components, 68 and prolonged blood circulation in vivo. 69 Further, when analyzing transgene expression in the liver after systemic delivery of pHPMA-decorated Ad vectors, an up to 10,000-fold decrease in transgene expression for pHPMAylated vectors was demonstrated. These data indicate that polymer modifications can be used for detargeting from the liver. 69 Virus particles with pHPMA coupled to the capsid surface via degradable disulfide bonds showed higher infectivity when compared with virions with covalently coupled pHPMA, 68 a phenomenon probably due to its multivalent nature. However, the beneficial effects of polymer shielding required large moieties which impaired virus bioactivity. 68 Further, amine-directed shielding occurs randomly throughout the whole vector surface and does not allow for shielding of specific capsomers. Hence, polymer-modified vector particles show high heterogeneity even within one preparation. Further, extensive capsid modifications are needed to achieve above described effects and in combination with genetic insertion of targeting ligands the subsequent shielding also would modify these. To overcome these limitations, Kreppel et al. 70 introduced a geneti-chemical concept for vector re- and detargeting. Cysteines were genetically introduced in the virus capsid at solvent-exposed positions like fiber HI-loop, 70 protein IX, 71 and hexon. 72,73 Although not naturally occurring, cysteine-bearing Ad vectors can be produced at high titers in normal producer cells. Importantly, insertion of cysteines in not only certain capsomers, but also in different positions within one capsomer, allows for highly specific modifications with thiol group-reactive moieties. This geneti-chemical approach has been shown to overcome numerous obstacles in Ad vector design. The combination of amine-based PEGylation for detargeting and thiol-based coupling of transferrin to the fiber knob HI-loop has been proven to successfully retarget modified Ad vectors to CAR-deficient cells. 70 Since hexon is involved in most undesired interactions (neutralizing antibodies, blood coagulation factor FX), thiol-based modification strategies were also applied to hexon. Coupling small PEG moieties to HVR5 of hexon prevented Ad vector particles to transduce SKOV-3 cells in the presence of FX, whereas large PEG moieties increased hepatocyte transduction. 72,74 Ad vector particles carrying mutations in the fiber knob to inhibit CAR binding and in HVR7 inhibiting binding of FX, and bearing inserted cysteines in HVR1 for position-specific PEGylation, were shown to evade antibody- and complement-mediated neutralization as well as scavenger receptor-mediated uptake, without loss of infectivity. Interestingly, despite lack of the natural FX shield, PEGylation again improved transduction of hepatocytes as a function of PEG size 73 . However, it was shown that covalent shielding does have an impact on intracellular trafficking processes. Prill et al. compared irreversible versus bioresponsive shields based on pHPMA and demonstrated that neither the mode of shielding nor co-polymer charge had an impact on cell entry, but rather affected particle trafficking to the nucleus. Employing a bioresponsive shield with positively charged pHPMA co-polymers allowed for particle trafficking to the nucleus maintaining the high transduction efficiencies of Ad vectors in vitro and in vivo. 75
In summary, these data indicate that, even under the assumption all vector–host interactions were known and considered, excessive capsid modifications are necessary to overcome all hurdles associated with systemic vector delivery. The individual modification strategies possess their specific limitations. While genetic strategies are limited in the number of positions that can be modified without impairing production titers, the insufficient characterization of other human Ad types is limiting for the development of novel chimeric Ad viruses. The chemical approach at least overcomes limitations due to coupled molecule sizes and attachment stability; nevertheless, excessive polymer shielding impairs viral bioactivity and requires bioreversibility for efficient disassembly in the cytoplasm. Therefore, for successful utilization of Ad vectors in gene therapeutic approaches a rational and application-specific combination of genetic and chemical modification strategies is inevitably.
Virotherapy for Cancer Treatment
For systemic delivery of therapeutic Ad vectors, riddance of unwanted vector–host interactions is mandatory. This can be deemed an enhanced understanding of detargeting. At the same time, efficient targeting approaches are required. However, only the combination of highly specific targeting technologies and the addressing of all unwanted vector–host interactions at once will lead to the successful construction of safe and efficient therapeutic Ad vectors.
Oncolytic cancer therapy is a strategy that uses engineered viruses to treat malignancies. Oncolytic viruses (OVs) are modified such as they specifically infect cancer cells and replicate in these cells only. Malignant cells may be eliminated directly by the viral infection and tumor cell lysis, releasing additional virus particles to infect neighboring cells. Also, therapeutic viruses activate and support the immune system to recognize and attack tumor cells and unmask clandestine tumor antigens. A broad range of OV has been delivered by intratumoral injections with a perceptible success in treating reachable solid tumors. 76 However, for the treatment of advanced or metastatic malignancies, or inaccessible tumor entities such as pancreatic cancer, a systemic delivery of OVs is favorable, allowing the simultaneous treatment of primary tumor and disseminated metastases. Nevertheless, the systemic delivery of virus particles is complex and faces specific hurdles that will be discussed in the context of cancer virotherapy in the following.
The systemic delivery of OV by i.v. injection requires careful considerations regarding tumor targeting. Cancer cell–specific promoters (e.g., hTERT) and surface markers that are selectively expressed on tumor cells, such as EGF receptor, HER2/neu, and folate receptor can be used for transductional targeting and conditional replication. For example, genetic insertion of fiber displaying a single-domain antibody directed against human carcinoembryonic antigen (hCEA) in combination with the cancer cell-specific CXC4 promoter resulted in a retargeted and oncolytic conditionally replicating Ad vector (CRAd) showing increased infection of hCEA positive cells. 77 Likewise, an oncolytic CRAd expressing the human CD40 ligand induced activation of dendritic cells leading to an induction of a Th1 response and increased tumor-specific T-cells. 78 However, cancer cells exhibit high rates of mutation and chromosomal rearrangement, giving the tumor a high plasticity to modulate gene expression that allows for adaption to any kind of cytotoxic stress. This also results in a in a high intratumoral heterogeneity: different parts of the tumor may express different genes and therefore present different tumor cell-specific receptors. This limits the utilization of genetic modifications of the Ad capsid. Incorporation of peptides or the generation chimeric viruses may result in partial targeting of only a subset of tumor cells. Another challenging aspect is the presence of a dense stroma composed of different kinds of cells (e.g., fibroblasts, macrophages, endothelial cells), blood vessels, and extracellular matrix. Diffusion of OV is blocked by this stroma and tumor cells that are sparsely embedded within evade antitumor therapeutics. Since Ad viruses are highly immunogenic, a collateral induction of innate and adaptive immune responses against tumor cells has been observed 79 and is currently explored as an approach in oncolytic virotherapy (reviewed in Woller et al. 80 ). However, the efficient delivery of OV to the tumor by the blood stream represents another hurdle. As outlined above, in nonphysiological conditions when injected in the blood stream, Ad vector particles transduce hepatocytes. Interactions with soluble factors and cellular receptors result in fast clearance with a half-life of minutes. Ablating CAR and integrin binding did not affect liver tropism, 18 whereas in combination with fiber shaft exchange a reduced gene transfer into hepatocytes was observed. 29,30 However, compared to conventional Ad5 chimeric Ad vectors were shown to trigger robust immune responses. 38 Further, the presentation of chimeric capsid proteins may eventually trigger the generation of adapted neutralizing antibodies, impeding repeated administration of therapeutic Ad vectors.
A key role in hepatocyte transduction can be drawn to capsid protein hexon. Binding of blood coagulation factor FX to Ad5 hexon HVRs bridges the virus to heparan sulphate proteoglycans on the surface of hepatocytes. The pharmacological or genetical inhibition of FX-hexon interaction resulted in reduced transduction rates of the liver. 19,20 In mice, macrophages and Kupffer cells are the main cells removing Ad vectors from the blood using scavenger receptor A (SR-A). Combining ablation of FX-binding to hexon with ablation of SR-A binding or binding to other scavenger receptors such as SREC-1 81 will be interesting next steps, as a combined pharmacological inhibition of these factors resulted in tumor targeting. 82 However, since neutralizing anti-Ad antibodies are also mainly directed to hexon, particle “coating” by FX protects the virus from host immunoglobulin M and complement-mediated neutralization. In humans, neutralizing antibodies will have a major impact on vector half-life. If tumor targeting using Ad vectors is challenging in Ad-naïve patients, it is even more challenging in patients that have experience Ad vector treatment before or have preexisting neutralizing antibodies. In this scenario, shielding virus particles with complex polymers represents a promising approach. Shielding of Ad6 with low seroprvalence with PEG blunted liver damage and cytokine production after systemic delivery but may also reduce its oncolytic efficacy, 83 The position-specific oupling of PEG using a geneti-chemical approach 70 enables for both particle shielding and genetic capsid modifications for transductional targeting. As demonstrated recently, the combination of genetically ablated CAR and FX binding with PEGylation of HVR1 prevented complement and antibody neutralization as well as particle sequestration via scavenger receptor-mediated mechanisms, 73 Due to polymer attachment, Ad vector particles are slightly increased in size, which may have an interesting side effect on liver transduction. The size of the sinusoidal fenestrae in humans without liver pathology (107 ± 1.5 nm) 84 allows unmodified Ad particles (80–100 nm) to translocate. Studies in rabbits suggest that the sinusoidal fenestrae size may function as a natural barrier for Ad vectors when particle sizes are increased. 5
Being the most abundant blood cell type, binding of Ad5 to human erythrocytes is of high relevance. Human erythrocytes were shown to sequester circulating Ad5 via CAR and complement receptor 1–mediated binding. 85 Due to the lack of integrins on human erythrocytes surface, CAR-mediated binding is reversible and does not prevent extravasation and organ transduction 86 . However, complement receptor 1–mediated binding of Ad5 to erythrocytes is targeted for rapid clearance, presumably involving the classic complement pathway. 85
Despite the encouraging results of oncolytic Ad5-viruses being currently tested in clinical trials (reviewed in Uusi-Kerttula et al. 87 ), rapid elimination of the therapeutic vector from the bloodstream upon systemic delivery severely limits their use to either Ad5-naïve patients and/or intra-tumoral or ex vivo applications. Noteworthy, the ablation of the ability of Ad5 particles to bind FX detargeted the particles from hepatocytes, 20 but at the same time rendered them highly susceptible for neutralization by natural antibodies and complement. 88 This example demonstrates that the vector biology in an organism is much more complex than simple on/off switches. However, since Ad5 is the best-characterized type so far, and given the high immune-prevalence for Ad5 type, further development and combination of the above discussed approaches is absolutely essential for the design of safe and efficient therapeutic vectors. In this context, the genti-chemical approach might offer advantages. The comparably minor modifications typically maintain the potency of the virus and can be applied to both well-characterized Ad types such as Ad5 but also to largely unknown types. Being a rational approach, geneti-chemical capsid modifications are solely based on the knowledge of hurdles that need to be overcome. A wide choice of ligands like antibodies, peptides, aptamers, affilins, lipids, and carbohydrates can be coupled by stable maleimide and bismaleimide groups or in a bioreversible manner forming e.g., disulphide bridges, yielding in defined particles. However, a vector for successful systemic delivery through the blood stream will very likely be generated by a combination of existing technologies.
Vaccination
Traditionally, vaccination against viral pathogens is based on attenuated or inactivated viruses or virus subunits. Progress in molecular virology and immunology has led to the generation of vectored vaccines, expressing an antigen of choice. Due to their aptitude to induce strong innate and adaptive immune responses, to efficiently transduce many cell types including professional antigen presenting cells (APC) and the potential adjuvant function of hexon, Ad vectors have been extensively studied as vaccination vectors.
Capsid modifications for targeting and shielding
A successful vaccination strategy requires the induction of a cytokine profile orchestrating the maturation of B- and cytotoxic T-cells, resulting in a prolonged and boostable immune response. Upon vector-receptor interaction, Ad5 was shown to trigger rapid release of IL-6, Il-12, and TNFα 89 thus activating the differentiation of immature dendritic cells to professional APC. Induction of CD8+ cells is primarily stimulated by de novo synthesized peptides, while CD4+ cells are favorably activated by peptides derived from phagocytosis and lysosomal cleavage. Naturally, Ad virions cause lytic infections resulting in short antigen presentation time of individually infected cells, thereby potentially favoring a CD4+ immune response. Most Ad vectors currently used for vaccination are replication defective or single cycle, 90 resulting in prolonged antigen presentation time for both transgene and capsid proteins. Additionally, the generation of chimeric Ad5 vectors carrying fiber from Ad30 and Ad35 resulted in increased transduction of dendritic cells without affecting transgene-specific B- or T-cell responses, 91 Another study demonstrated that besides improved tropism for dendritic cells, an Ad5/Ad35 chimeric vector also enhanced induction of CD8+ cells in vitro and in vivo. 92 Incorporation of an RGD peptide in Ad5 fiber knob also increased the efficiency in transducing dendritic cells and was shown to induce higher transgene-specific CD4+- and CD8+-specific immune responses. 93 Further, due to the promiscuous nature of Ad vectors, leakage of vector particles from injection sites such as muscle or skin, will contribute to systemic transduction of nonlymphoid tissue, particularly the liver. It was demonstrated that vector dissemination from an intramuscular injection site to the liver resulted in early priming of transgene-specific CD8+ cells. However, hepatocyte-specific silencing mediated by miRNAs significantly enhanced early priming of transgene-specific CD8+ cells, allowing up to 100-fold reduction of vector dose. 94 However, in pre-exposed humans, hexon-specific CD4+ and CD8+ T-cell responses have been demonstrated. 95 Binding of neutralizing Ad-specific antibodies prevents cell transduction and thus transgene (epitope) expression and presentation. Recent efforts to circumvent antibody neutralization focus on Ad types exhibiting low serum prevalence in human population. Promising candidates such as human Ad6, as well as chimpanzee (Ch) Ad types ChAd3 and ChAd63 were studied for their use as vectored vaccines for hepatitis C virus, malaria, and HIV. 96 –98 Another strategy to overcome Ab neutralization may be the utilization of Ad dodecahedrons. In the 1960s, the spontaneous formation of symmetric particles that were significantly smaller than Ad virions was observed in transduced cells. 99,100 In the late 1990s the spontaneous formation of these dodecahedral particles was observed upon expression of either Ad3 penton base alone or combined expression of penton base and fiber. 101,102 Since Ad dodecahedrons contain fiber and integrin-binding RGD motif in penton base they can efficiently enter cells, whereas the lack of hexon greatly reduced the probability of being neutralized by anti-Ad antibodies. Another strategy to overcome antibody-mediated neutralization is chemical shielding of vector particles, as described above. Shielding of Ad vectors with PEG resulted in efficient detargeting upon systemic and intramuscular delivery. Despite being coated, PEGylated Ad vectors were still capable to induce a potent transgene-specific immune response while evading neutralization by preexisting antibodies. 103
Capsid modifications for antigen presentation
An alternative approach to vector-encoded epitope expression is to incorporate the antigen in the viral capsid, either by genetic fusion or using adapter molecules. This strategy has firstly been used in 1994. An eight-amino-acid sequence of the VP1 capsid protein of poliovirus type 3 was genetically incorporated into two regions of Ad2 hexon. Antiserum derived from one of these vectors was shown to specifically recognize the VP1 capsid protein. 48 In similar studies, the immunodominant 14 -amino-acid peptide Epi8 of pseudomonas was incorporated in Ad5 HVR5. Mice vaccinated with this vector showed an increased antibody response consisting of anti-pseudomonas IgG1 and IgG2s subtypes and a remarkable number of pre-immunized mice survived when challenged with pseudomonas. 104 In a rather exceptional approach, a third-generation cocaine hapten termed GNE was conjugated to the capsid of a disrupted Ad5 vector. Vaccination with this vector evoked anti-cocaine antibodies in rats and prevented cocaine-induced hyperactivity and addiction related behavior. 105 In nonhuman primates, repetitive delivery maintained high levels of anti-cocaine titers that were sufficient to prevent cocaine from binding to dopamine transporter. 106 However, the presentation of target epitopes, either vector-encoded or capsid-incorporated, competes with numerous viral proteins for major histocompatibility complex (MHC) molecules and thus the release of stimulatory cytokines, which are needed for activation and proliferation of T- and B-cells. This might have negative consequences for the desired immune response. Therefore, one obstacle is to keep the balance between Ad- and transgene-specific immune responses; exploiting the adjuvant function of Ad capsid proteins while inducing a strong transgene-specific immune response. This might be achieved with a partial, position-specific shielding of hexon to preserve its adjuvant function but decrease the probability of neutralization and unwanted immune responses at the same time.
Besides being less prone to antibody neutralization, Ad dodecahedrons also proved to be suitable to present antigens on their surface. The prolin-containing motifs xPPxY in penton base were exploited to couple protein cargo to Ad dodecahedral particles by fusing WW domains to target proteins. Using this technique full-length proteins up to 150 kDa in size can be efficiently coupled to dodecahedra and transported into cells (reviewed by Kron and Kreppel 107 ). Ad3 dodecahedra coupled with the influenza matrix protein M1 entered myeloid dendritic cells with an efficiency of up to 90% and exhibited an immunostimulatory effect. 108 The potential of Ad dodecahedra to induce humoral and cellular immune responses in vivo was further investigated with the model antigen ovalbumin. Upon subcutaneous delivery of ovalbumin-loaded dodecahedra in mice, ovalbumin-specific T-cells were detected in splenocytes. Moreover, these ovalbumin-loaded dodecahedra could induce ovalbumin-specific antibody response and titers could be boosted upon repeated delivery. 109 In summary, these studies impressively demonstrate the high potential of Ad dodecahedra in vaccination strategies.
Despite the advances in understanding the molecular basis of diseases, molecular virology, and viral immunology, little is known about the mechanisms of Ad APC transduction in vivo and the number of APC that must be transduced to induce a potent immune response. 91,110,111 Therefore, the rational design of Ad-based vaccines requires a thorough knowledge of the infectious agent the vaccination vector is developed for. However, although additional obstacles lie ahead the application of Ad vectors for vaccination looks promising.
Footnotes
Acknowledgments
This work was partly funded by the Center for Biomedical Education and Research (ZBAF) of the Witten/Herdecke University (UW/H).
Author Disclosure
No competing financial interests exist.