
Abstract
Adeno-associated viral (AAV) vectors can target the liver, making them an attractive platform for gene therapy approaches that require the correction of hepatocytes. Crigler–Najjar syndrome is an autosomal recessive disorder of bilirubin metabolism that occurs when the liver's uridine diphosphate glucuronosyl transferase 1A1 (UGT1A1) enzyme activity is partially or completely absent. This syndrome is characterized by elevated bilirubin levels in the blood. An AAV8 vector was developed expressing a codon-optimized human version of UGT1A1 from a liver-specific promoter. High doses of the vector rescued neonatal lethality in newborn UGT1 knockout (KO) mice, which serve as a model of Crigler–Najjar syndrome, and significantly increased survival from 5 to 270 days. Newborn UGT1 KO mice treated with AAV had serum total bilirubin levels that were 5.7 times higher than the levels seen in heterozygous and wild-type mice, likely due to dilution of vector genome copies (GC) in the liver resulting from a proliferation of hepatocytes during growth of the animal. The elevation in serum total bilirubin levels in adult UGT1 KO mice depended on the AAV8 vector dose. At doses <1011 GC/mouse, total bilirubin levels returned to those seen in phototherapy-rescued UGT1 KO mice. Mice injected with vector at 1011 or 3 × 1011 GC/mouse had sustained reduced total bilirubin levels throughout the duration of the study. When an AAV8 vector was re-administered in mice with elevated total bilirubin levels, serum total bilirubin levels decreased to wild-type levels (0.1–0.3 mg/dL) in mice that received a vector dose of 3 × 1012 GC/kg. Therefore, a low-level and likely transient decrease in serum total bilirubin during the first days of life is necessary for rescuing the lethal phenotype present in the neonatal UGT1 KO mouse. Furthermore, it was possible to ablate the elevated total bilirubin levels in adult mice by re-administering an AAV8 vector.
Introduction
C
Although several drugs can slightly reduce jaundice, current medical management consists of phototherapy for at least 12 h/day. 3 However, phototherapy rapidly becomes less effective following puberty, thus increasing the risk for kernicterus and creating the need for liver transplantation to control the disease. An alternative approach of gene replacement therapy would be more effective because Crigler–Najjar syndrome is caused by the deficiency of a single gene product, UGT1A1. Compared to a liver transplant, it would be less risky and more effective to deliver a gene therapy vector expressing UGT1A1 systemically so that the liver could continuously synthesize UGT1A1.
Ugt1–/– or UGT1 knockout (KO) mouse models have been developed that resemble certain key aspects of Crigler–Najjar syndrome. 4,5 These mouse strains have either a neomycin cassette inserted into exon 4 of the Ugt1 gene locus or a point mutation identical to that of the Gunn rat, which is a classical rat model of Crigler–Najjar. 4 –7 Consequently, homozygous UGT1 KO mice display postnatal lethality, hyperbilirubinemia, and jaundice. UGT1 KO mice exhibit very high serum levels of unconjugated bilirubin (7.82 mg/dL vs. 0.13 mg/dL in wild-type or heterozygous mice, representing an approximate 60-fold increase), appear jaundiced within 8 h of birth, and die 7 days after birth. 4 Phototherapy, the current standard of care, has been shown to prolong survival. 5,8 –10
Given that expression of the UGT1A1 transgene could protect UGT1 KO mice from lethal hyperbilirubinemia in the immediate postnatal period, administering gene therapy in the critical neonatal period may rescue the newborn phenotype. Researchers have previously administered gene therapy on postnatal day 4 (P4) via intraperitoneal injection. 8,10 However, in a clinical setting, the more likely route of administration is an intravenous (i.v.) injection of the vector because this is the most efficient way to target the human liver, the clinical site of this syndrome. Injections i.v. have been used for other liver diseases with neonatal phenotypes. 11,12 In this study, an adeno-associated viral (AAV) serotype 8 vector was developed expressing a codon-optimized human version of UGT1A1 from a liver-specific promoter. The vector was administered within 24 h of birth via i.v. injection. In order to evaluate the duration of gene expression, the effects of gene therapy were followed for 1 year post vector administration. A gene therapy vector was also re-administered with the same capsid to evaluate the efficacy of this approach in mice that exhibited increased total bilirubin levels over the course of the study.
Materials and Methods
AAV vector production
All AAV vectors were produced by the Penn Vector Core at the University of Pennsylvania, as previously described. 13 Briefly, plasmids expressing codon-optimized versions of human UGT1A1 (hUGT1A1co) from the thyroxine binding globulin (TBG) promoter in the presence or absence of the woodchuck hepatitis post-transcriptional regulatory element (WPRE) were packaged within the AAV8 viral capsid.
Mice
Breeding pairs of heterozygous Ugt1+/– mice were obtained from The Jackson Laboratory (Bar Harbor, ME), 4 and a colony was maintained at the University of Pennsylvania under specific pathogen-free conditions. All animal procedures and protocols were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania. UGT1 KO mice and heterozygous and wild-type littermates received an i.v. injection with 109–3 × 1011 genome copies (GC)/mouse of AAV8.TBG.hUGT1A1co.WPRE.BGH within 24 h of birth via the facial temporal vein. Four mice rescued by newborn injection of 3 × 109 GC/mouse were re-administered 3 × 1011 or 3 × 1012 GC/kg of AAV8.TBG.hUGT1A1co.BGH by i.v. injection on day 56 of life via the tail vein. Phototherapy-rescued UGT1 KO mice were exposed to phototherapy immediately after birth (blue fluorescent light, λ = 450 nm; 10–30 μW/cm2/nm) for 12 h/day for up to 21 days after birth.
Serum analyses
Blood was collected in serum separator tubes and allowed to clot. Serum was isolated and analyzed for total bilirubin levels by Antech Diagnostics (Irvine, CA).
Immunohistochemistry
Mice were necropsied 270 days post vector administration, and the liver was harvested. Liver samples were fixed in formalin for a minimum of 24 h and paraffin embedded. Sections were deparaffinized through an ethanol and xylene series, boiled for 6 min in 10 mM of citrate buffer (pH 6.0) for antigen retrieval, and sequentially treated with 2% H2O2 (15 min), avidin/biotin blocking reagents (15 min each; Vector Laboratories, Burlingame, CA), and blocking buffer (1% donkey serum in phosphate-buffered saline with 0.2% Triton for 10 min). Sections were incubated with 5 μg/mL of the primary antibody against human UGT1A1 (R&D Systems, Minneapolis, MN) for 1 h and then a biotinylated secondary antibody (Jackson ImmunoResearch, West Grove, PA) diluted in blocking buffer for 45 min. A Vectastain Elite ABC kit (Vector Laboratories) was used according to the manufacturer's instructions with 3,3′-diaminobenzidine as the substrate to stain bound antibodies as a brown precipitate.
Quantification of UGT1A1-positive liver area (equivalent to percent positive hepatocytes) was performed by thresholding each image to select the UGT1A1-positive area using ImageJ software (Rasband W.S., National Institutes of Health;
Neutralizing antibody assay
Neutralizing antibodies (NAbs) to the AAV8 capsid were determined, as previously described. 14
Statistical analysis
Comparison of survival curves was performed using a log-rank (Mantel–Cox) test. For serum total bilirubin data, cohort average and standard deviation (SD) was calculated and reported. Comparisons between total bilirubin levels in newborn AAV-treated UGT1 KO, heterozygous, and wild-type mice were performed using linear mixed-effect modeling. Comparisons between total bilirubin levels at day 28 in newborn AAV- and phototherapy-treated UGT1 KO mice were performed using a Wilcoxon rank-sum test. A one-way analysis of variance with Dunnett's multiple comparisons test was performed across vector doses to determine any vector-related effects compared to age-matched UGT1 KO mice rescued to adulthood by phototherapy. Dose groups were compared to each other using a two-sample Wilcoxon rank-sum test. A p-value of <0.05 was considered significant.
Results
Rescue of neonatal lethality in the UGT1 KO mouse model of Crigler–Najjar
The UGT1 KO mouse model of Crigler–Najjar exhibits severe neonatal lethality. 4,5 Therefore, litters generated by mating heterozygous Ugt1+/– mice were observed on a daily basis until weaning at day 21 post birth. Pups that were either found dead or euthanized for clinical reasons, including lethargy and dehydration, were genotyped, and survival curves were generated (Fig. 1A). UGT1 KO mice did not survive past the first week of life, whereas heterozygous and wild-type littermates displayed normal survival. Others have previously shown that phototherapy, the current standard of care for human patients, can also be used to treat the UGT1 KO mouse. 8 –10 UGT1 KO mice were exposed to phototherapy (blue fluorescent light, λ = 450 nm; 10–30 μW/cm2/nm) immediately after birth for 12 h/day for up to 21 days after birth. Upon weaning, UGT1 KO mice were also removed from phototherapy. These animals did not require maintenance on phototherapy after weaning, and the majority of mice demonstrated normal survival (Fig. 1B). However, out of 21 weaned mice, three UGT1 KO mice either died or had to be euthanized due to clinical signs at 34, 82, and 159 days of life.
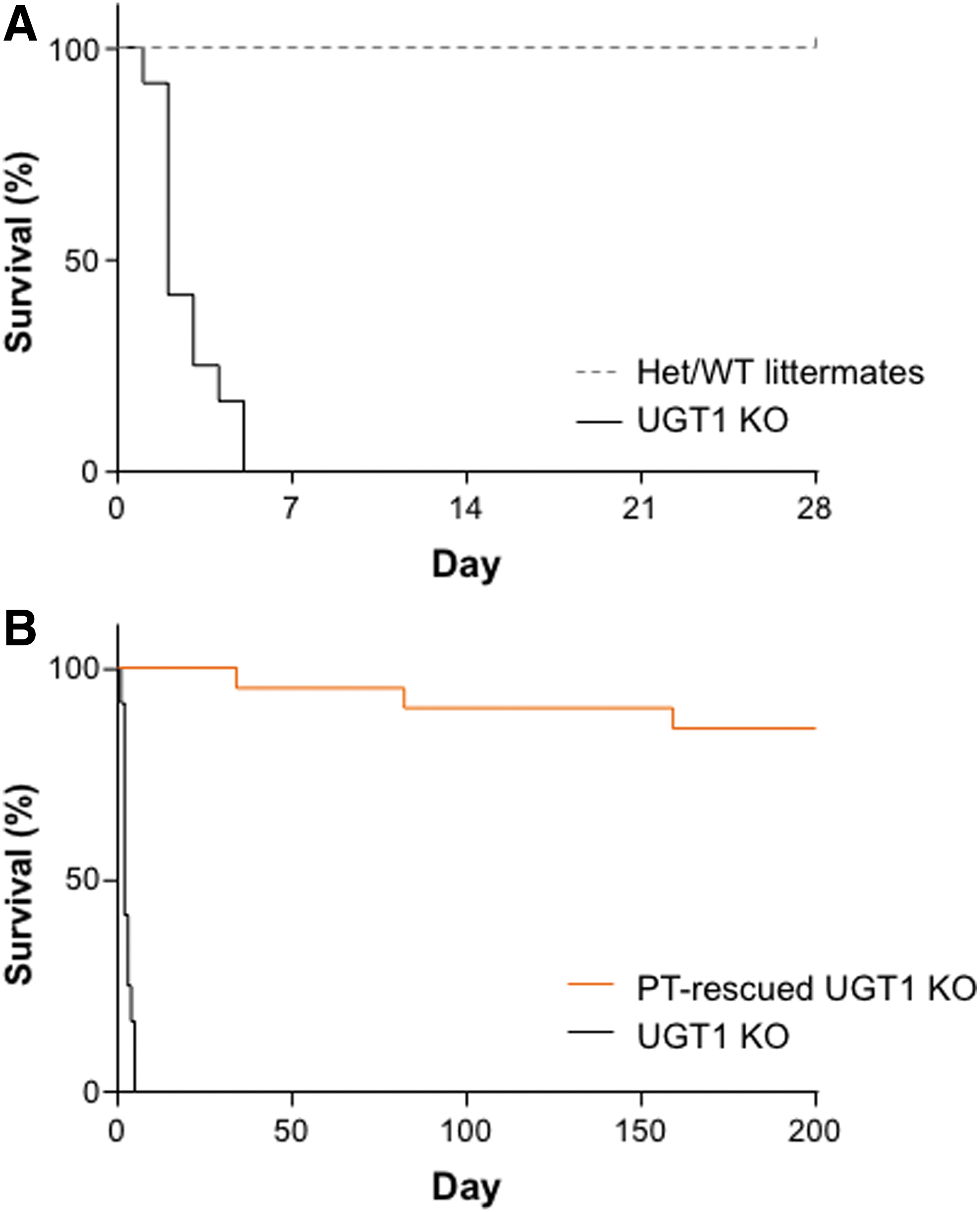
Neonatal lethality occurs within the first week of life in UGT1 knockout (KO) mice.
It was hypothesized that gene therapy provided within the neonatal period could protect UGT1 KO mice from lethal hyperbilirubinemia in the immediate postnatal period. Mice generated from heterozygous Ugt1+/– mating received an i.v. injection of 1011 GC/mouse of AAV8.TBG.hUGT1A1co.WPRE.BGH within 24 h of birth via the facial temporal vein. Mice were observed daily until weaning at day 21 post birth, and blood was collected weekly starting on day 28 for determination of serum total bilirubin levels. Compared to untreated UGT1 KO mice, those treated with the AAV8 vector demonstrated significantly increased survival from day 5 to day 270 when the study terminated (Fig. 2A; p < 0.0001). From clinical observations, rescued UGT1 KO mice appeared phenotypically identical to their heterozygous and wild-type littermates, albeit with elevated total bilirubin levels. Serum total bilirubin levels in the AAV-treated UGT1 KO mice were elevated 5.7-fold compared to heterozygous and wild-type animals (Fig. 2B and C; p < 0.01). The rise in total bilirubin levels from day 28 to a plateau at around day 100 is likely due to dilution of vector GC in the liver as a result of the proliferation of hepatocytes during growth. This has been observed for other liver diseases where newborn vector administration has been performed. 11
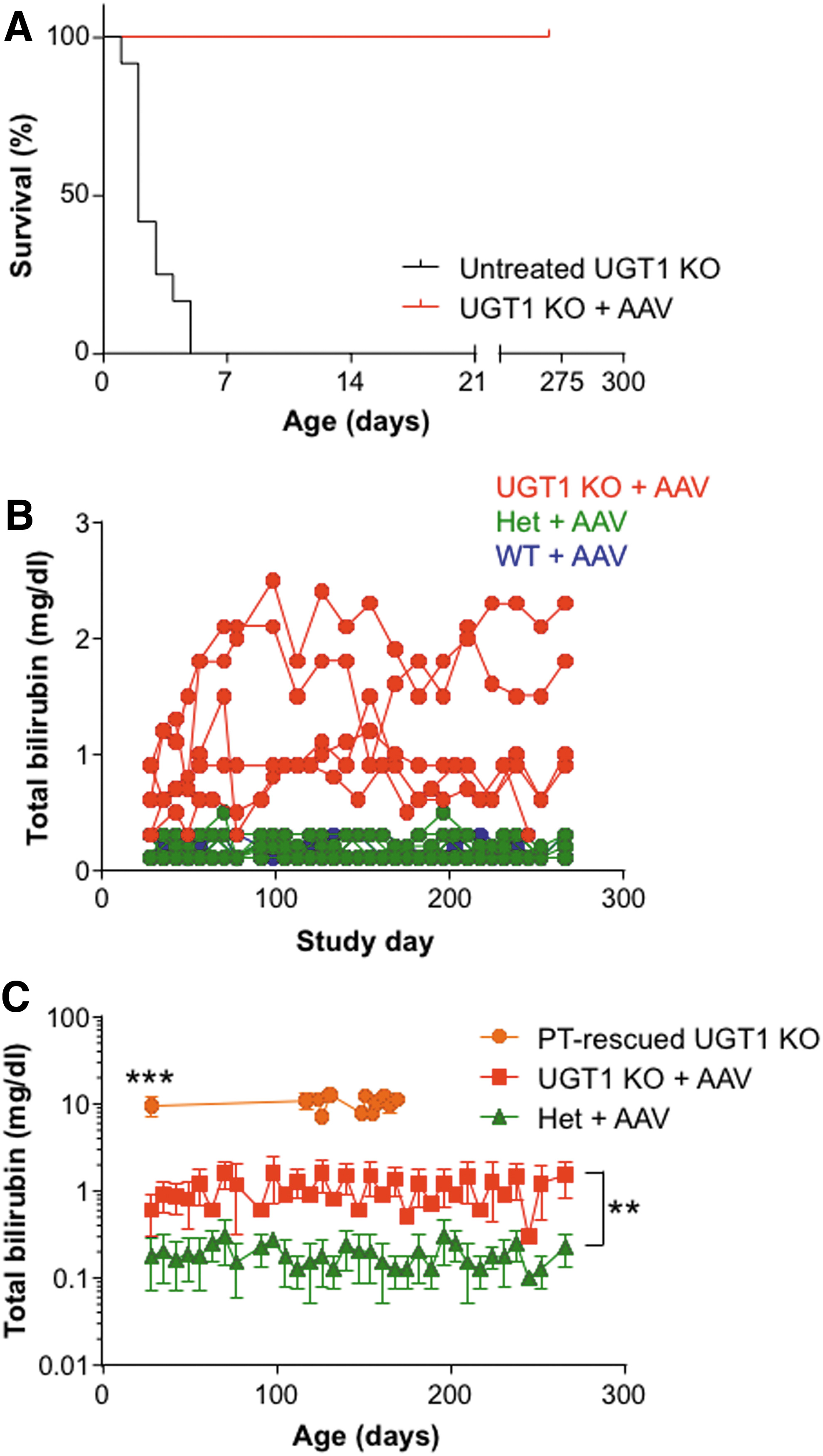
UGT1 KO mice have increased survival and decreased total bilirubin levels following intravenous (i.v.) injection of adeno-associated viral (AAV) serotype 8 vector within 24 h of birth.
Comparison of AAV8 treatment to phototherapy for newborn rescue of the UGT1 KO mouse
Similar to human patients with Crigler–Najjar, UGT1 KO mice also respond to phototherapy. Adult UGT1 KO mice that had been rescued with phototherapy were bled, and serum total bilirubin levels were determined (Fig. 2C). The average serum total bilirubin levels in UGT1 KO mice rescued with phototherapy was 9.1 mg/dL (n = 52). In comparison to phototherapy-rescued UGT1 KO mice, serum total bilirubin levels in AAV8-treated UGT1 KO mice were reduced 15.2-fold at day 28 (p < 0.001). Therefore, either phototherapy or gene therapy can provide sufficient relief of hyperbilirubinemia in the immediate postnatal period to allow survival to adulthood.
Sporadic and clonal hepatocyte transduction was sufficient for maintenance of adult UGT1 KO mice
AAV8-treated UGT1 KO mice were euthanized at day 270 post vector administration, and the liver was harvested for immunohistochemistry (IHC) staining of the human UGT1A1 protein (Fig. 3). The liver from an untreated UGT1 KO mouse was included as a control (Fig. 3A). Diffuse background staining was seen from endogenous mouse UGT1A1 expression in both AAV-treated heterozygous (Fig. 3B) and wild-type (Fig. 3C) littermates with dark staining of single cells by IHC, evidence of hepatocytes highly expressing hUGT1A1. In UGT1 KO mice, these areas were abundant throughout the liver, as seen at 10 × and 4 × magnification (Fig. 3D and E, respectively), leading to sufficient correction of hyperbilirubinemia. Images were quantified to determine the percentage of transduced hepatocytes, which ranged from 1% to 3% of the liver (1.9 ± 0.6%, mean [M] ± SD).
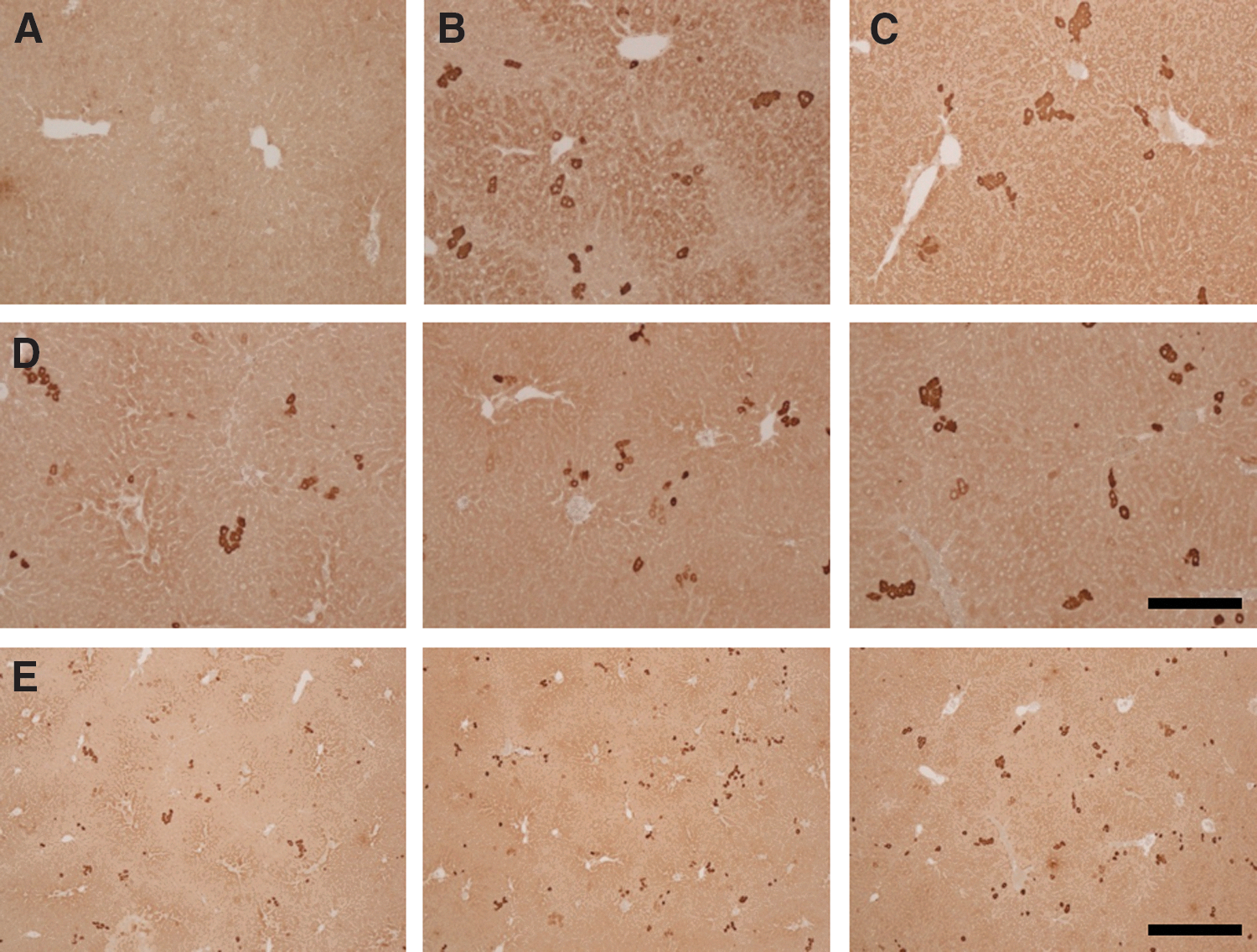
UGT1A1 IHC in the liver from AAV8 vector-injected UGT1 KO, heterozygous, and wild-type mice. Mice were injected i.v. with 1011 GC/mouse of AAV8.TBG.hUGT1A1co.WPRE.BGH within 24 h of birth via the facial temporal vein. All mice were genotyped at weaning. Mice were necropsied at 270 days post vector administration, and immunohistochemistry staining was performed on liver sections for detection of UGT1A1 in
Serum total bilirubin levels in adult UGT1 KO mice were AAV8 vector dose dependent
To evaluate the minimum AAV8 vector dose required for UGT1 KO mouse survival, a dose-ranging study was performed. Thirty UGT1 KO mice were rescued from neonatal lethality by administering each with a dose of AAV8 vector (109–3 × 1011 GC/mouse) via the facial temporal vein within 24 h of birth. Compared to adult phototherapy-rescued UGT1 KO mice (orange bar in Fig. 4A; p < 0.01), serum total bilirubin levels at day 28 post vector administration were significantly reduced in mice administered with >109 GC/mouse AAV8 vector. There was a clear dose–response effect at day 28 post vector administration, with decreased serum total bilirubin levels in mice administered with increasing AAV8 vector doses (Fig. 4A). At the lowest dose (109 GC/mouse), one out of six injected mice survived to adulthood, with a 37% reduction in serum total bilirubin compared to phototherapy-rescued UGT1 KO mice. Total bilirubin levels of UGT1 KO mice administered with <1011 GC/mouse AAV8 vector returned to those seen in phototherapy-rescued UGT1 KO mice during the 1-year follow up (Fig. 4B). However, mice injected with 1011 or 3 × 1011 GC/mouse of vector maintained reduced total bilirubin levels throughout the study. This dose-ranging study provides additional evidence that a low-level and likely transient decrease in serum total bilirubin during the first days of life is required for rescue of the neonatal lethal phenotype of the UGT1 KO mouse.

Dose-dependent serum total bilirubin levels in UGT1 KO mice following i.v. injection of AAV8 vector within 24 h of birth. UGT1 KO mice were injected i.v. with one of a range of doses of AAV8.TBG.hUGT1A1co.WPRE.BGH (109–3 × 1011 GC/mouse) within 24 h of birth via the facial temporal vein. Total bilirubin levels were determined in serum at
Re-administration with AAV8 vector further reduces serum total bilirubin levels in adult UGT1 KO mice
As the effect of low-dose gene therapy on serum total bilirubin levels in UGT1 KO mice slowly reversed to the hyperbilirubinemia levels seen in phototherapy-rescued UGT1 KO mice, AAV8 vector was re-administered. UGT1 KO mice received an i.v. injection with 3 × 109 GC/mouse of AAV8.TBG.hUGT1A1co.WPRE.BGH via the facial temporal vein within 24 h of birth. Mice were bled weekly from day 28 (Fig. 5A). To determine the feasibility of re-administration, NAbs to the AAV8 capsid were investigated on serum samples collected at day 35. Three out of the four mice were negative for AAV8 NAbs (<1:5), and one had a titer of 1:5. Low-level or negligible NAb titers following newborn i.v. vector administration has been reported previously. 11 Re-administration was carried out with 3 × 1011 GC/kg or 3 × 1012 GC/kg of AAV8.TBG.hUGT1A1co.BGH vector to all four mice on day 56 via the tail vein (Fig. 5B). Serum total bilirubin levels decreased to wild-type levels (0.1–0.3 mg/dL) in mice administered with 3 × 1012 GC/kg of the vector. However, administration with a vector dose one log lower did not reduce total bilirubin levels. Therefore, re-administration with a high dose of AAV8 vector was effective, but only at a high vector dose.

Re-administration of AAV8 vector causes a further decrease in serum total bilirubin levels in UGT1 KO mice rescued by AAV8 vector treatment within 24 h of birth.
Discussion
The ability to evaluate gene therapy applications for genetic diseases in animal models is crucial to the development of effective therapies. The UGT1 KO mouse not only serves as a mouse model for Crigler–Najjar syndrome, but also enables the evaluation of gene therapy approaches in the presence and absence of the current standard of care—phototherapy. In comparison, the traditional animal model for Crigler–Najjar syndrome, the Gunn rat, does not require phototherapy for survival to adulthood. Furthermore, previous studies suggest that certain AAV vector capsids transduce the liver less efficiently in rats compared to mice. 6,7,15,16 Therefore, the decision was made to evaluate a gene therapy approach in the UGT1 KO mouse.
It was found that i.v. neonatal gene therapy with AAV8.TBG.hUGT1A1co.WPRE.BGH protected UGT1 KO mice from lethal hyperbilirubinemia in the immediate postnatal period, and significantly increased survival from 5 days in untreated UGT1 KO mice to 270 days. Based on clinical observations, rescued UGT1 KO mice appeared phenotypically identical to their heterozygous and wild-type littermates. However, the serum total bilirubin levels in the AAV8-administered UGT1 KO mice were elevated 5.7-fold compared to heterozygous and wild-type animals, but were reduced 15.2-fold compared to UGT1 KO mice rescued to adulthood by phototherapy. Previous studies have shown similar results with intraperitoneal vector injections in P4 mice. 8,10 However, here the rise in serum total bilirubin appeared to lessen over time, and the use of i.v. administration of the vector in this study is more clinically relevant.
IHC for the hUGT1A1 protein showed highly transduced sporadic and clonal hepatocytes where the AAV genome had likely integrated during proliferation of hepatocytes and growth of the mouse. This overexpression of UGT1A1 in 1–3% of hepatocytes in the adult mice was sufficient to produce a 93% reduction in total bilirubin levels. This approach may be better for the correction of hyperbilirubinemia than low-level transduction across all hepatocytes. This hypothesis has been previously suggested for Crigler–Najjar applications, as hepatocyte transplantation studies in Gunn rats and in humans have resulted in low levels of engraftment (most recently 2.5–7.5%) and reductions in bilirubin levels of 30–60%. 17 –19 However, for other gene therapy applications, different AAV vector capsids have been selected for clinical use to allow expression in a high percentage of hepatocytes to increase levels of a circulating transgene product. 20
Random integrations of the AAV genome can occur, especially during growth, 21 leading to concerns over the risk of developing hepatocellular carcinoma (HCC). All of the doses evaluated here by newborn administration are within the range of doses that have been previously evaluated for potential HCC generation. 22 For this proof-of-concept study, the initial vector used also contained WPRE, which has been shown to cause a high frequency of HCC cases in a mouse model of phenylketonuria. 23 Administration of gene therapy vectors to newborn mice may make them more susceptible to HCC generation, but the risk is dose dependent. 22,24 Therefore, this risk may be mitigated by using the lowest possible dose for rescue of the newborn phenotype.
Following dose-ranging studies, it was demonstrated that even lower levels of hUGT1A1 expression were required to allow survival of the UGT1 KO mouse to adulthood. Mice administered with 3 × 109 GC/mouse (and one mouse administered with 109 GC/mouse) were protected against lethal postnatal hyperbilirubinemia and were rescued to adulthood. Assuming an approximate mouse body weight of 1 g at the time of dosing (within the first 24 h after birth), 3 × 109 GC/mouse equates to a dose of ∼3 × 1012 GC/kg. However, serum total bilirubin levels did rise to an average of 5.6 mg/dL by day 56 of life due to dilution of hepatic vector GC during growth, but it was possible to re-administer mice with a similar AAV8 gene therapy vector (using the same capsid) due to low-level or negligible NAb titers following newborn i.v. vector administration. 11 Re-administration of AAV8 at a dose of 3 × 1012 GC/kg resulted in wild-type serum total bilirubin levels in adult UGT1 KO mice. Others have shown the potential to re-administer gene therapy vectors in adult UGT1 KO mice rescued by newborn administration. 10 However, here the effective re-administration dose was 10-fold lower (3 × 1012 GC/kg vs. ∼1.1 × 1013 vg/kg), and serotype switching was not performed for the second vector.
This method of newborn vector administration followed by re-administration into juveniles or adults, where the exponential phase of liver growth has slowed, could translate to the clinic. Newborn vector administration has been previously shown to be highly effective in macaques, indicating translation of this approach from mice to nonhuman primates. 25 Following identification of children with Crigler–Najjar syndrome, there are two opportunities where the first round of the AAV gene therapy could be applied: either at birth or at 7–11 months of age when there is a decline in NAbs due to a drop in maternal antibody levels. 26 Continuous expression of UGT1A1 from a gene therapy vector may be better able to control bilirubin levels than the current standard of care—phototherapy—due to compliance issues. As the liver grows, there may be a decrease in gene therapy efficacy over time. If serum total bilirubin levels reach unacceptable levels, there would be a number of options available: (1) phototherapy, (2) re-administration of gene therapy, or (3) a liver transplant. Based on the NAb titer of the patient, the gene therapy vector used for re-administration could be packaged with either the same capsid or an alternative capsid to which the patient is seronegative. Currently, liver transplantation is the only cure available for Crigler–Najjar syndrome, with most doctors aiming to perform this procedure early in life to prevent brain damage due to kernicterus. If re-administration was not desirable, administration of gene therapy to newborns could be used increase the window of opportunity to find a donor liver.
Footnotes
Acknowledgments
We would like to thank Deirdre McMenamin and Yanqing Zhu for invaluable technical assistance, and Mingyao Li and Yan Che for help with biostatistics. This research was supported by Audentes Therapeutics.
Author Disclosure
The authors declare having potential competing financial interests. J.M.W. is an advisor to, a founder of, holds equity in, and has a sponsored research agreement with REGENXBIO. He has equity in Solid Biosciences. He also has a sponsored research agreement with Ultragenyx. In addition, he is a consultant to several biopharmaceutical companies, and he is an inventor on patents licensed to various biopharmaceutical companies. J.A.G. and J.M.W. are co-inventors on a patent (WO2017/106345A1) covering technology related to this work, which has been licensed to Audentes Therapeutics. For all other authors, no competing financial interests exist.