
Abstract
Adeno-associated virus serotype 9 (AAV9) is an efficient vector for gene transfer to the myocardium. However, the use of ubiquitous promoters, such as the cytomegalovirus (CMV) promoter, can result in expression of the transgene in organs other than the heart. This study tested if the efficiency and specificity of cardiac transcription from a chicken cardiac troponin T (TnT) promoter could be further increased by incorporating a cardiomyocyte-specific transcriptional cis-regulatory motif from human calsequestrin 2 (CS-CRM4) into the expression cassette (Enh.TnT). The efficiency of luciferase expression from the TnT and Enh.TnT constructs was compared to expression of luciferase under the control of the CMV promoter in both adult and neonatal mice. Overall, expression levels of luciferase in the heart were similar in mice injected with AAV9.TnT.Luc, AAV9.Enh.TnT.Luc and AAV9.CMV.Luc. In contrast, expression levels of luciferase activity in nontarget organs, including the liver and muscle, was lower in mice injected with the AAV9.TnT.Luc compared to AAV9.CMV.Luc and was negligible with AAV9.Enh.TnT. In neonates, in organs other than the heart, luciferase expression levels were too low to be quantified for all constructs. Taken together, the data show that the AAV9 Enh.TnT constructs drives high levels of expression of the transgene in the myocardium, with insignificant expression in other organs. These properties reduce the risks associated with the AAV9-mediated expression of the therapeutic protein of interest in nontarget organs. The excellent cardiac specificity should allow for the use of higher vector doses than are currently used, which might be essential to achieve the levels of transgene expression necessary for therapeutic benefits. Taken together, the findings suggest that the Enh.TnT transcription unit is a potentially attractive tool for clinical cardiac gene therapy in adults.
Introduction
M
Cardiomyocyte “specific” promoters have been described before (e.g., Liu et al., 4 Müller et al., 5 Pleger et al., 6,7 Prasad et al., 8 and Raake et al. 9 ), but so far, all reported promoters (or promoter–enhancer combinations) displayed significant transgene expression in organs other than the heart and/or expressed the transgene at significantly reduced levels when compared to strong ubiquitous promoters such as the cytomegalovirus (CMV) promoter or the Rous sarcoma virus long terminal repeat (RSV) promoter.
We previously performed a genome-wide computational analysis to identify cis-regulatory motifs (CRMs) that enable efficient cardiac transduction. 10 In that study, a particularly promising sequence was identified from the human calsequestrin gene (CS-CRM4), which contains several transcription factor binding sites, such as HNF3α, MEF2, SRF, NF1, RSRFC4, HNF3β, and HFH1. 10 When coupled to the α-myosin heavy-chain (αMHC) promoter, cardiac expression was increased 100-fold compared to the αMHC promoter alone, reaching expression levels comparable to expression from a CMV promoter construct. 10 Unfortunately, the transgene was also expressed in significant amounts in skeletal muscle, and in failing human hearts, myosin heavy-chain expression is biased toward the expression of βMHC not αMHC. 10 Coupling of the same enhancer element to a synthetic promoter (Spc-5-1211) led to a further increase in cardiac expression, but comparable increases in expression were also observed in skeletal muscle. Among the most promising cardiac promoters described today is a minimal chicken cardiac troponin T promoter (TnT), which results in more cardiac specific expression when compared to expression driven by the CMV promoter, although nontarget organ expression was not completely eliminated. 8 The results of the two studies 8,10 prompted the evaluation of whether the addition of the cis-regulatory motif from the human calsequestrin gene to the TnT promoter could further enhance cardiac expression levels and/or reduce transgene expression in nontarget organs.
Here, we demonstrate that the addition of the calsequestrin enhancer sequence to the TnT promoter further reduces undesirable ectopic expression in the liver, one of the two major nontarget organs. Moreover, we show that expression using this novel transcription cassette (Enh.TnT) results in comparable or higher transgene (luciferase) levels in both adult mice and neonates when compared to transduction by AAV9 vectors using the CMV or the TnT promoter. These results suggest that the Enh.TnT transcriptional regulatory element, once it is validated in nonhuman primates, might be useful for clinical cardiac gene therapy.
Methods
Cells and plasmids
pACMV-Luc (hereafter CMV; Fig. 1) and pAcTnT-Luc (hereafter TnT; Fig. 1) were a kind gift from Brent French. 8 pTR-enhancer-TnT-Luciferase (Enh.TnT in Fig. 1) was created as follows. The enhancer (AGTAGAAAAACAGCCAAGCTAGGGAGGCTGGGAGGCCAAGCCCCAGATACCTTACATAGCTCTGCTCAGCCTCTGTCTCATTAGGAACTCCATTTTTAGGATGCAGTTGTTTCAGGCTAAAAATAAATCATGCAATGAATAAAAAAGTTAGATACGACACTGTAGAGGGATTCGCTGATACAGTCTGTCCGA) was removed from pAAV-Enh4-MHC-BGI-hrGFP10 by KpnI digest, blunted with Klenow fragment of DNA polymerase 1, and inserted between the inverted terminal repeat (ITR) and TnT promoter of pAcTnT-Luc at the SnaBI site. Sequence analysis confirmed the correct orientation.
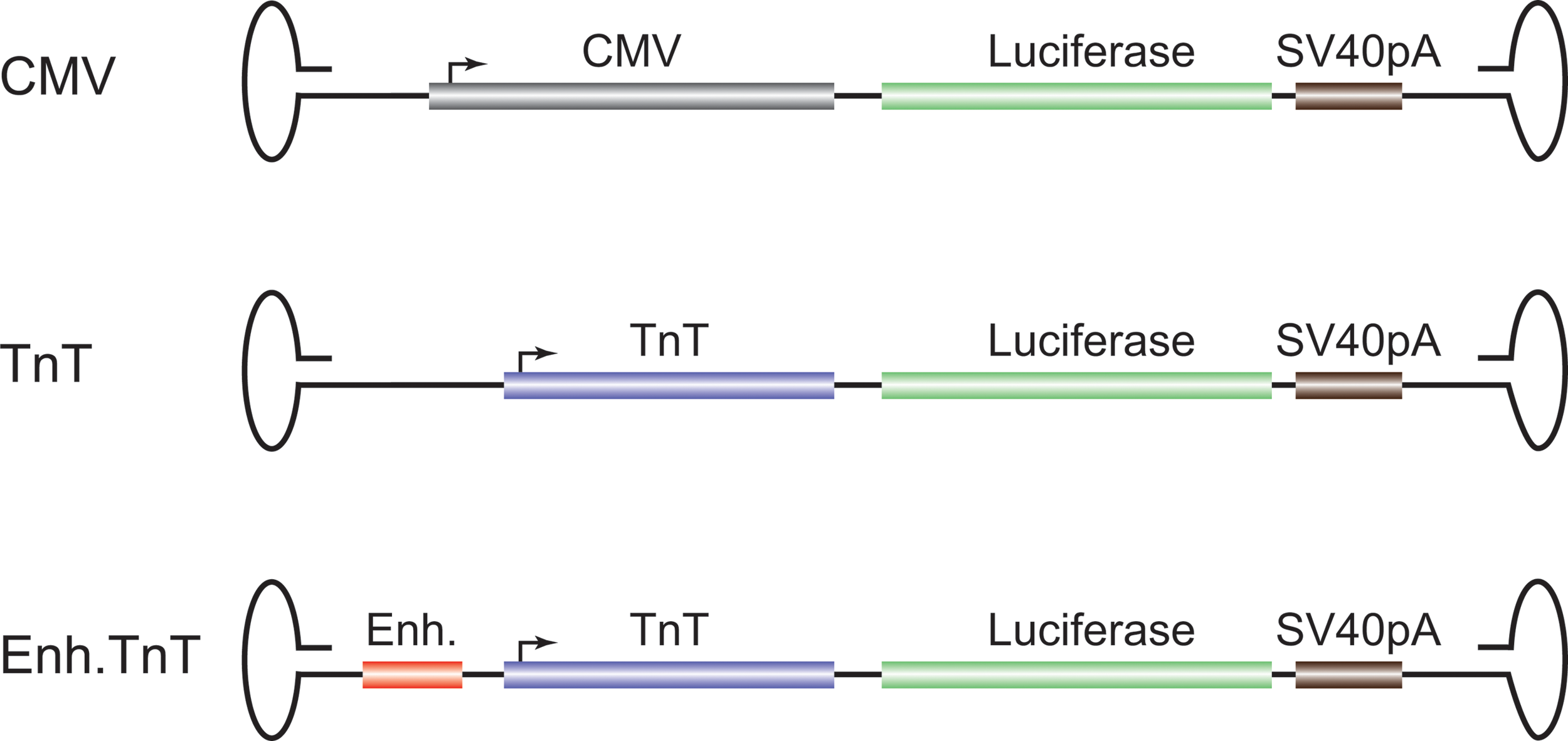
Genome organization of adeno-associated virus (AAV) cis-plasmids. The plasmids used to produce recombinant AAV serotype 9 (rAAV9) vectors contained a firefly luciferase expression cassette under the control of either: a cytomegalovirus (CMV) promoter, a chicken troponin T promoter (TnT, or a TnT with the addition of a calsequestrin enhancer. All constructs contained an SV40 poly A signal and were flanked by AAV2 inverted terminal repeats (ITRs). The sizes of the indicated elements, the complete expression cassettes, and the whole viral genomes are: CMV, 588 bp; TnT, 407 bp; Enh, 192 bp; luciferase, 1,653 bp; SV40pA, 241 bp; CMV expression cassette, 3,030 bp; TnT expression cassette, 2,849 bp; Enh.TnT expression cassette, 3,041 bp; CMV viral genome, 3,547 bp; TnT viral genome, 3,547 bp; Enh.TnT viral genome, 3,739 bp.
HEK 293T cells were grown in Dulbecco's modified Eagle's medium (DMEM; Gibco, Waltham, MA) supplemented with 10% fetal bovine serum (FBS; Crystalgen, Commack, NY) at 37°C in 5% CO 2 . The helper plasmid pDG9 was grown in DH5α cells in lysogeny broth containing 100 μg/mL of ampicillin at 37°C. In contrast, cis-plasmids encoding the luciferase expression cassettes flanked by ITRs were grown in SURE2 cells (Agilent, Santa Clara, CA) in lysogeny broth containing 100 μg/mL of ampicillin at 30°C. Maxiprep DNA for transfection was prepared using a Zymopure Plasmid Maxi Prep Kit (Zymo Research, Irvine, CA) according to the manufacturer's instructions.
Preparation of recombinant vectors
Recombinant AAVs were prepared as described, 12,13 with minor modifications. Briefly, the day before transfection triple flasks (total surface area 500 cm2) were each seeded with one confluent 15 cm dish of HEK 293T cells and allowed to grow overnight. Once 70% confluence was reached, transfection was performed by adding the following to 20 mL of room temperature DMEM: 50 μg of cis-plasmid, 150 μg of pDG9 (briefly vortex), and 250 μL of PEI-max (Polysciences, Warrington, PA) solution (pH 4.5). The solution was vortexed for 15–20 s and incubated at room temperature for 15 min. The transfection mix was then added to 90 mL of pre-warmed DMEM containing 2% FBS and mixed by swirling. The medium from the triple flasks was removed and replaced with the transfection mix. After 3 days of growth, the cells were harvested by vigorous tapping of the flask, transferred to a sterile container, and pelleted by centrifugation. The supernatant was kept for downstream processing. The cell pellet was resuspended in 10 mL of lysis buffer (150 mM of NaCl, 50 mM of Tris-HCl, pH 8.5), and freeze-thawed three times at −80°C and 37°C with brief vortexing after each thaw cycle. Non-encapsidated DNA and contaminating RNA was then removed with the addition of 2 μL (10 IU/μL) of Pierce Universal Nuclease (Thermo Fisher Scientific, Waltham, MA) followed by incubation for 30 min at room temperature. The crude cell lysate was centrifuged to pellet debris, and the supernatant was reserved for iodixanol (Sigma–Aldrich, St. Louis, MO) gradient ultracentrifugation. Virus from the cell culture supernatant was precipitated by the addition of 31.3 g of ammonium sulfate per 100 mL of supernatant followed by incubation on ice for at least 30 min. The precipitate was pelleted via centrifugation, resuspended in lysis buffer, and then recombined with the cell pellet supernatant for iodixanol gradient ultracentrifugation. Samples for ultracentrifugation were prepared in 32.4 mL of polypropylene Optiseal tubes (Beckman Coulter, Brea, CA). Viral lysates were loaded on top of discontinuous iodixanol gradients composed of 4 mL of 60% iodixanol, 4 mL of 40%, 4.9 mL of 25%, and 7.3 mL of 17% (with 1M sodium chloride). The gradients were centrifuged at 350,333 g (avg) for 60 min at 18°C in a Beckman type 70 Ti fixed angle rotor. Fractions (1.25 mL) were collected from the bottom of the tube and kept for virus titration. Peak fractions were dialyzed in lactated Ringer's solution (Baxter international, Deerfield, IL), filtered through a 0.22 μm pore filter (Merck Millipore, Billerica, MA), and stored at −80°C.
Determination of recombinant AAV viral titer
Viral titers were determined by quantitative polymerase chain reaction (qPCR) using primers that bind to the luciferase region: (fwd: 5′-ACACCCGAGGGGGATGATAA-3′; rev: 5′-GTGTTCGTCTTCGTCCCAGT-3′). A standard curve was made by making serial dilutions of luciferase cis-plasmid in nuclease-free water. Dialyzed virus stocks were diluted 1:1,000 in nuclease-free water and qPCR mixtures were made up using the iTaq Universal SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA). The qPCR was carried out according to the manufacturer's instructions in a 7500 Real Time PCR System thermal cycler (Applied Biosystems, Foster City, CA) with the following thermal cycling program: one cycle of 95°C for 5 min followed by 40 cycles of 95°C for 10 s, 58°C for 10 s, and 72°C for 45 s.
Animal procedures
All procedures involving the handling of animals were approved by the Institutional Animal Care and Use Committee (IACUC) of the Icahn School of Medicine at Mount Sinai or by the IACUC of the University of Texas Health Sciences Center at Houston and adhered with the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health. All animals were provided with nesting material, had ad libitum access to food and water, and were housed in a facility with a 12 h/12 h light/dark cycle. For experiments with adult mice, 4- to 5-week-old male and female BALB/c or C57BL/6 mice were purchased from Charles River Laboratories (Wilmington, MA). Adult mice were temporarily restrained and injected into the tail vein with either 1e11 vector genomes (vg) or 2e10 vg diluted in Lactated Ringer's solution in a total volume of 60 μL using a 27G insulin syringe. They were then sacrificed either 1 or 3 months later using CO2, and the brain, heart, liver, lung, kidneys, and quadricep muscles were immediately harvested and stored at −80°C.
Neonatal mice were injected subcutaneously on postnatal days 2, 3, and 4 with 50 μL of lactated Ringer's solution containing 6.7e9 vg each. The mice were sacrificed at 1, 2, and 4 weeks after injection (from first day of injection). Heart, liver, skeletal muscle, kidney, lung, and brain tissues were collected and frozen.
All mouse organs were crushed into a fine powder using a pestle and mortar that had been pre-chilled on dry ice and were stored at −80°C.
Luciferase protein assay
Samples for luciferase assay analysis were prepared in luciferase cell culture lysis reagent (Promega, Madison, WI). For adult mice, approximately 25 mg of crushed tissue from each organ was mixed with 500 μL of 1 × lysis buffer, vortexed for 15 s, and then agitated at room temperature for 15 min. The samples were subjected to three cycles of freeze-thawing at −80°C to 37°C with brief vortexing after each thaw cycle and centrifuged at 12,000 g for 3 min to pellet debris. Then, the supernatant was removed and placed in a new tube. The pellet was subjected to a second round of protein extraction using the same method (with the exception of the freeze–thaw cycles), and the second supernatant was combined with the first to make a total sample volume of 1 mL. Lysates of organs from animals injected as neonates were prepared in accordance with the protocol described in Manthorpe et al. 14
Protein content was determined by Pierce BCA protein assay (Thermo Fisher Scientific) according to the manufacturer's instructions. Luciferase activity was determined with the Promega luciferase assay system. Luminescence was measured with a luminescence counter: Microbeta Trilux 1450 LSC and Luminescence Counter (Perkin Elmer, Carlsbad, CA) for adult mice and a TECAN- infinite M200 plate reader (Tecan Trading AG, Männendorf, Switzerland) for neonatal mice. Luciferase activity was normalized to protein content and is expressed throughout as relative light units per mg protein (RLU/mg).
DNA isolation and qPCR
Total DNA from the heart, liver, and muscle tissue was purified using a Qiagen Blood and Tissue DNA extraction kit (Qiagen, Hilden, Germany) according the manufacturer's instructions and eluted in 200 μL of elution buffer. Viral genome copy numbers were determined by qPCR using the same conditions used for virus titering and normalized to the total amount of DNA present.
Statistical analysis
Statistical significance of differences between individual groups was determined by a two-tailed Mann–Whitney U-test using GraphPad Prism v7 (GraphPad Software, La Jolla, CA).
Results
To test the hypothesis that the addition of a previously identified 192 bp long human calsequestrin 2 enhancer motif (CS-CRM4) 10 upstream of a chicken cardiac TnT could enhance the tissue specificity and/or increase expression levels in the myocardium when compared to the CMV and TnT promoters, three AAV constructs were designed (Fig. 1). They contained the CMV promoter, the TnT promoter, or the calsequestrin enhancer TnT promoter combination driving the expression of firefly luciferase. Then, AAV9 vectors were produced to compare the transduction in the heart and other major organs after systemic injection into mice.
In a first step, transduction by the different AAV9 constructs was analyzed in adult male and female BALB/c mice. BALB/c mice were chosen because the intention was to use noninvasive IVIS® imaging to follow the time course of luciferase expression, which is easier in white BALB/c mice than in dark-skinned C57BL/6 mice. However, because the extensive expression of luciferase in the liver in mice injected with AAV9.CMV.Luc prevented the measurement of cardiac expression with this method, a decision was made to concentrate instead on the analysis of luciferase expression in organs harvested from animals sacrificed at two different time points. The different vectors (1e11 vg) were injected into 4- to 5-week-old mice via the tail vein. One month post injection, the mice were sacrificed, and the major organs were harvested. Transgene expression was analyzed by in vitro measurement of luciferase activity. As can be seen from Fig. 2, all three AAV9 vectors caused high levels of luciferase expression in the heart, with the AAV9.Enh.TnT construct showing the strongest expression of luciferase, although this did not reach statistical significance. The most significant nontarget organ expression with AAV9.CMV.Luc was observed in the liver and skeletal muscle (quadriceps; Fig. 2). Luciferase expression driven by AAV9.CMV.Luc in the liver and the muscle reached about 50–60% of the expression levels in the heart. In contrast, expression in the liver of animals injected with the AAV9.TnT.Luc construct was around 40-fold lower (p < 0.0001), and expression in the liver in animals injected with the AAV9.Enh.TnT vector was >400-fold lower than in the heart (p < 0.0001 vs. CMV; p < 0.001 TnT vs. Enh.TnT). Expression in the skeletal muscle was >500-fold lower than the TnT and Enh.TnT constructs when compared to levels obtained with AAV9.CMV.Luc (p < 0.0001).

Luciferase activity in the major organs of adult BALB/c mice 1 month post vector injection with 1e11 vector genomes (vg) of recombinant AAV9 (rAAV9). Adult male and female BALB/c mice were injected with 1e11 vg of rAAV9 containing a luciferase expression cassette under the expression of a CMV (male n = 4, female n = 5), a TnT (male n = 4, female n = 4), or an Enh.TnT (male n = 4, female n = 5) promoter. After 1 month, the mice were sacrificed, and the major organs were harvested and frozen at −80°C. The organs were crushed to a fine powder, the tissues were lysed in lysis buffer, and luciferase activity and protein quantities were determined, as described in the Methods. Non-injected mice (female n = 2, male n = 1) served as controls. Luciferase activity was normalized to protein content and expressed as relative light units (RLU)/mg protein. Boxes correspond to upper and lower quartiles, the horizontal line represents the median, and the whiskers mark the minimum and maximum values. Statistical significance of differences between individual groups was determined by a two-tailed Mann–Whitney U-test. *p < 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001. For clarity, statistical differences were not indicated for injected versus control (naïve) groups.
Because C57BL/6 mice are widely used to test AAV gene therapy approaches to treat cardiac disorders, luciferase expression levels were also tested in C57BL/6 mice injected with the three AAV9 vectors. Compared to BALB/C mice, similar expression patterns were obtained in C57BL/6 mice (Supplementary Fig. S1; Supplementary Data are available online at
Next, the effect of vector dose on transduction and whether expression levels were maintained over an extended period of time were analyzed. To this end, additional cohorts of mice were injected via the tail vein with either 2e10 vg or 1e11 vg of AAV9.CMV.Luc, AAV9.TnT.Luc, or AAV9.Enh.TnT.Luc. One group each of mice injected with 2e10 vg and 1e11 vg was sacrificed 3 months post injection, and an additional group injected with 2e10 vg was sacrificed after 1 month. Luciferase activity was then determined from tissue lysates of the heart, liver, and muscle (the data for the 1-month 1e11 group was obtained from the experiments performed for Fig. 2). Unsurprisingly, overall expression levels were lower in the 2e10 vg groups than in the 1e11 vg group. However, the differences varied widely, especially at the 1 month time point. For instance, at 1 month, expression in the heart of mice injected with 1e11 vg of AAV9.Enh.TnT.Luc was 175-fold higher than mice that were injected with 2e10 vg of the same vector. On the other hand, the difference in muscle expression in mice injected with 2e10 versus 1e11 vg of either AAV9.TnT.Luc or AAV9.En.TnT.Luc was less than twofold. Of the four cohorts tested, expression in the heart from the Enh.TnT constructs was highest in all groups, except in the 2e10 vg cohort that was sacrificed 1 month post injection, but after 3 months, the Enh.TnT construct was again the highest expressing construct, although this did not reach statistical significance (Fig. 3). Not unexpectedly, expression in the liver in mice sacrificed at 3 months after administration of AAV9.CMV.Luc was around 220-fold (p < 0.0001) lower when compared to the 1 month group—presumably due to silencing of the CMV promoter. 15 –18 Interestingly, expression in the liver also decreased in mice injected with the TnT construct (∼10-fold; p = 0002). The near background levels of luciferase activity in the liver of mice injected with 1e11 vg of AAV9.Enh.TnT.Luc were maintained at 3 months (Fig. 3). It is worth noting that the silencing was more pronounced in females than in males (p < 0.05 for both AAV9.CMV.Luc and AAV9.TnT.Luc). Strikingly, expression in the heart in mice injected with 1e11 vg of AAV9.CMV.Luc appeared to be much lower (∼30-fold) at 3 months than at 1 month, but this did not reach statistical significance, and was not the case in the 2e10 vg group. Interestingly, the lower expression at 3 months in the 1e11 vg AAV9.CMV.Luc group was due solely to a statistically significant decrease in expression in males (p = 0.016).
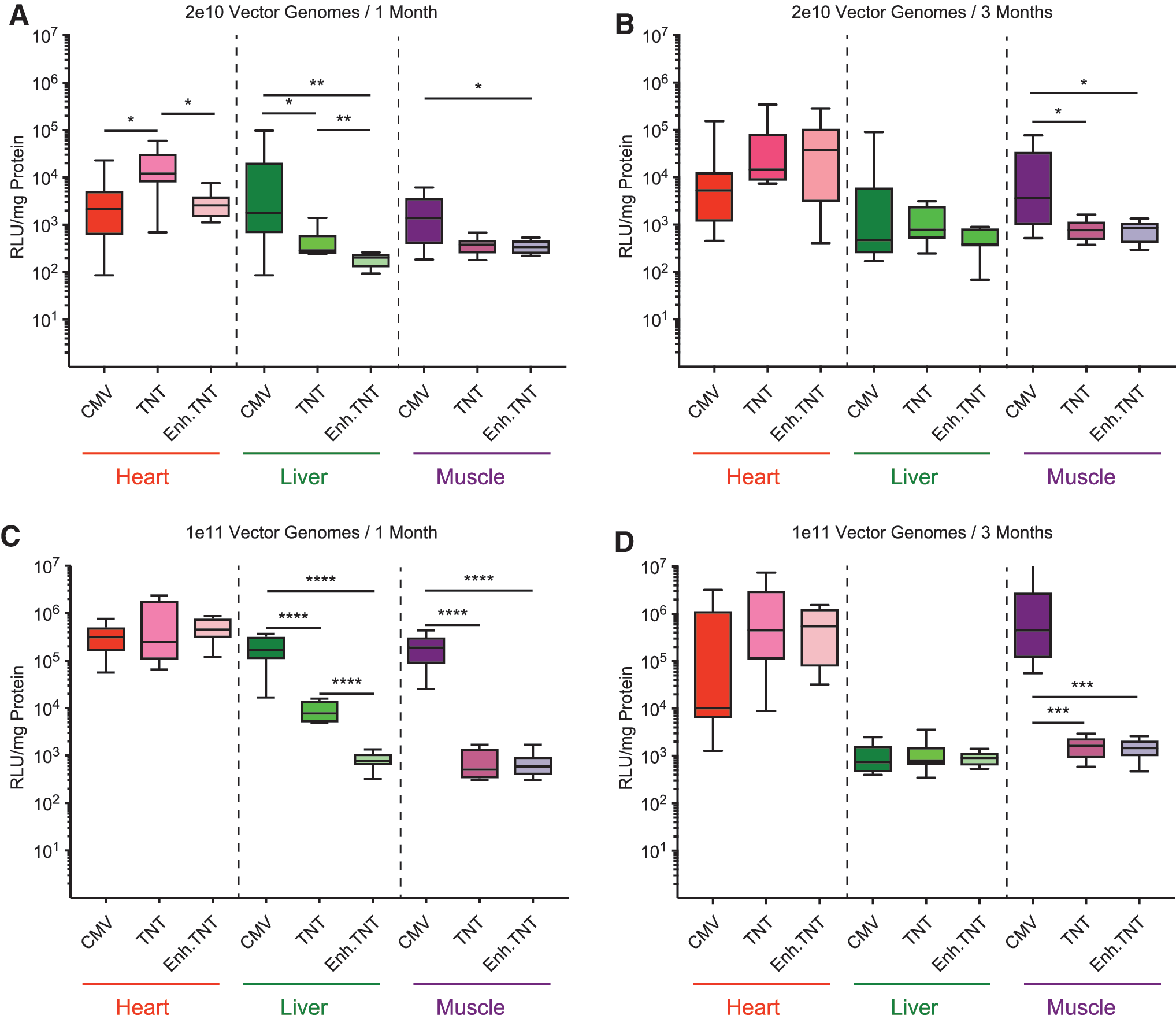
Luciferase activity in the heart, liver, and skeletal muscle of adult BALB/c mice injected with a low or high dose of rAAV9 and sacrificed at 1 or 3 months after vector injection. Adult male and female BALB/c mice were injected with either 2e10 vg
To determine if the differences in luciferase expression observed were due to variations in vg copies in the heart, liver, and muscle, the number of viral genomes was determined by qPCR in mice injected with 1e11 vg and sacrificed at 1 or 3 months. The increased vg numbers of TnT and Enh.TnT in muscle of mice sacrificed after 1 month further demonstrate the strong transcriptional detargeting. In mice sacrificed at 3 months, there were no differences in vg numbers among the different promoter groups. These data confirm that variations in expression observed in mice at 3 months were due to differences in transcriptional activity among the three different constructs (Fig. 4).
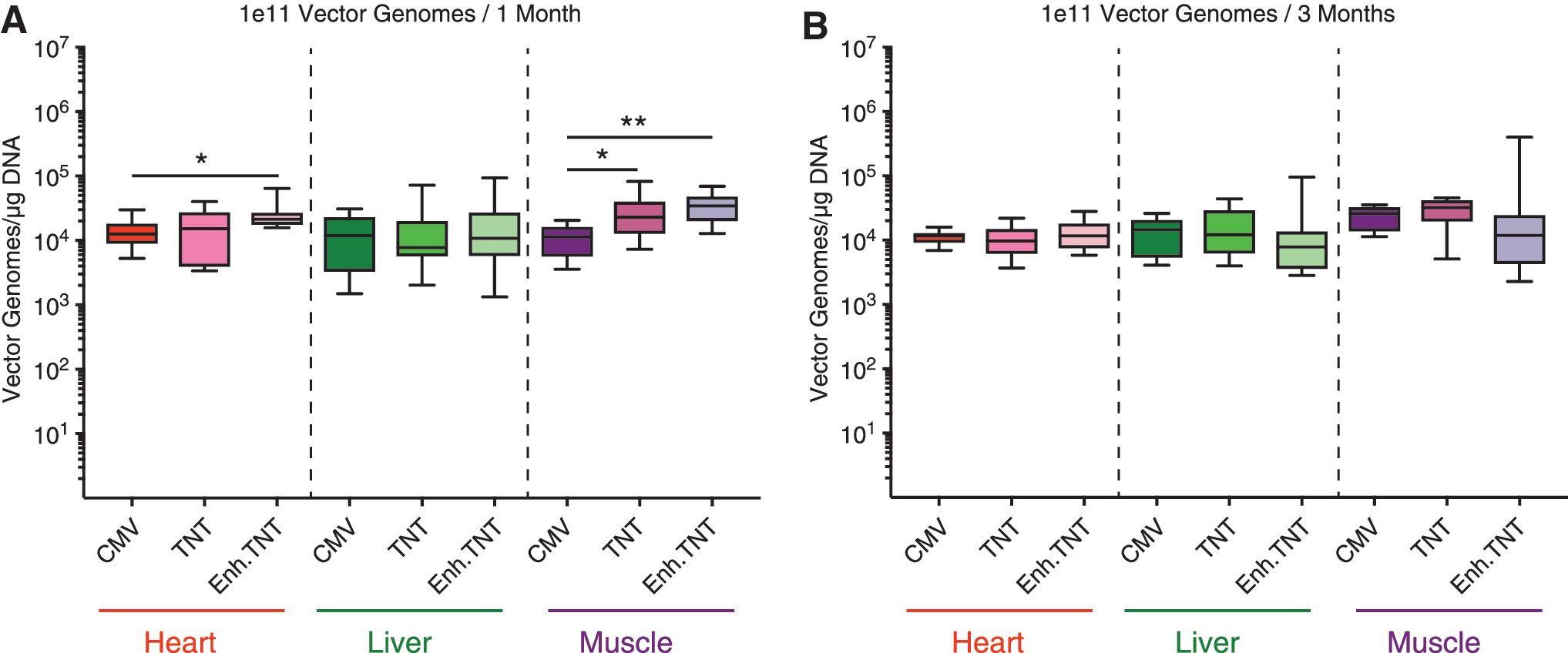
Viral genome copy number per microgram of total DNA in the heart, liver, and muscle of BALB/c mice at 1 and 3 months post injection (p.i.). Adult male and female mice were injected with either 1e11 vg of rAAV9 encoding luciferase under the expression of a CMV promoter (1 month: male n = 4, female n = 4; 3 months: male n = 4, female n = 5), a TnT promoter (1 and 3 months: male n = 4, female n = 4), or an Enh.TnT promoter/enhancer combination (1 month: male n = 4, female n = 4; 3 months: male n = 5, female n = 4) promoter.
One of the major goals of future cardiac gene therapy is the treatment of acquired heart failure, a disease occurring mostly in adults. Consequently, testing potential treatments for heart failure in preclinical models is performed in adult animals. On the other hand, there is a large number of inherited cardiomyopathies 19,20 for which gene transfer as soon as possible after birth, or even in utero, would likely offer the highest therapeutic benefit. Thus, the three different constructs were also compared in neonatal mice.
Neonatal mice were injected with 0.67e9 vg each on postnatal days 2, 3, and 4. The animals were sacrificed at 4 weeks after the first vector injection, the major organs were harvested, and luciferase expression was measured in vitro. Similarly to adult mice, 4 weeks after vector injection, all three AAV9 vectors showed strong luciferase expression in the heart, with the median luciferase expression from the TnT and Enh.TnT vectors being higher than expression from mice injected with AAV9.CMV.Luc, although this did not reach statistical significance (Fig. 5). In animals injected with AAV9.CMV.Luc, median expression was highest in the heart, followed by the muscle (∼1.7-fold lower), the liver (∼11-fold lower), and the lung (∼24-fold lower). In animals injected with AAV9.TnT.Luc or AAV9.Enh.TnT.Luc, raw luciferase activity values in all organs except the heart were never higher than twofold above background (i.e., luciferase expression was negligible or nonexistent).
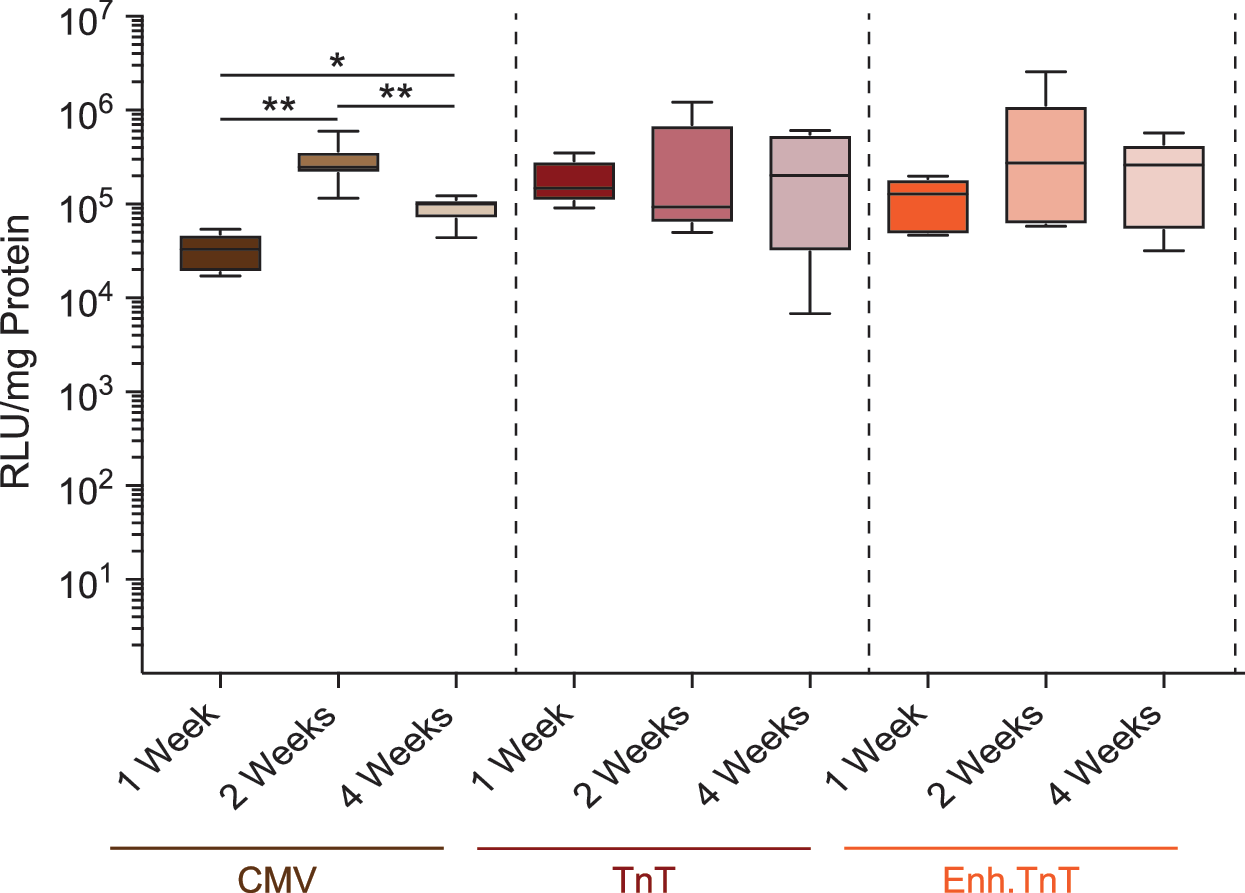
Luciferase activity in the heart of mice injected as neonates at 1, 2, and 4 weeks p.i. Neonatal mice were each injected with three equal doses (0.67e9 vg) of CMV (n = 16), TnT (n = 15), or Enh.TnT (n = 17) luciferase rAAV9 at 2, 3, and 4 days after birth. Mice injected with the three different vectors were sacrificed 1, 2, or 4 weeks p.i., and luciferase activity and protein content were determined, as described in the Methods. Boxes correspond to upper and lower quartiles, the horizontal line represents the median, and the whiskers mark the minimum and maximum values. Statistical significance between individual groups was determined by a two-tailed Mann–Whitney U-test. *p < 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001. ND, not determined.
Finally, the study determined whether expression levels were stable in the heart of neonates injected with the three AAV9 constructs. As can be seen from Fig. 5B, statistically significant variations between the 1-, 2-, and 4-week groups only existed in animals injected with AAV9.CMV.Luc, whereas expression levels showed no statistically significant differences in mice injected with AAV9.TnT.Luc or AAV9.Enh.TnT.Luc among the three time points.
Discussion
Cardiac gene therapy with AAV vectors is a promising avenue for the treatment of both acquired and inherited cardiovascular disorders. 21,22 Among the most promising therapeutic proteins for the treatment of heart failure are SERCA2A, stromal cell-derived factor 1, and adenylylcyclase 6, all of which have been or are being tested in clinical trials. In addition, overexpression of S100A1, VEGF, and others have already been tested in large-animal models. 21 So far, the treatment of inherited cardiomyopathies 19 –21 has not been tested in clinical trials, although a clinical trial to treat the cardiac manifestations of Friedreich's ataxia is to be initiated soon. In addition, there is a large number of inherited cardiomyopathies that might be amenable for AAV gene therapy. 19 –21 Whereas some of the proteins that if mutated cause cardiomyopathies are too large to be incorporated in a single AAV vector (e.g., myosin heavy chains 6 and 7 [∼5.8 kb] or desmoplakin [∼8.6 kb]), others such as cardiac-type myosin-binding protein C (∼3.8 kb) and desmoglein (∼3.4 kb) are small enough to be packaged into single-stranded AAV vectors and others (e.g., cardiac troponin I [0.6 kb] or the ventricular isoform of the myosin light chain [∼0.6 kb]) could even be packaged in self-complementary AAV vectors. 17,21,23 In order to bring cardiac gene therapy to the clinic, however, the ability to express the therapeutic transgene at high levels and with high specificity in the myocardium will be essential.
In addition to gene replacement approaches, gene editing with the CRISPR/Cas9 system using AAV vectors as delivery vehicles has recently been shown to improve cardiac function in the mdx model of Duchenne's muscular dystrophy. 24,25 In this particular case, the restriction of gene correction to cardiomyocytes might not be beneficial because correction in skeletal muscle, and especially in the diaphragm, would offer additional therapeutic benefit. However, in different settings, the exclusive expression of Cas9 in cardiomyocytes would undoubtedly be advantageous, as it would limit the risk of potential off-target genome modification to cardiomyocytes and reduce the risk of a strong immune response against Cas9.
Currently, even in rodents, there are no AAV serotypes or variants available that allow the exclusive expression of transgenes in cardiomyocytes. Whereas the widely used AAV9, upon systemic injection, can efficiently transduce cardiomyocytes in the hearts of mice and rats, it also results in expression in other organs, in particular the liver and muscle (Fig. 2). 3
Depending on the transgene, expression of the therapeutic protein in nontarget cell types can be either inconsequential or potentially result in serious adverse effects. For instance, the treatment of limb girdle muscular dystrophy by delivering the deficient protein, calpain 3, with an AAV9 vector construct in which calpain 3 expression was driven by a desmin promoter, which is active in both skeletal muscle and cardiomyocytes, resulted in extensive cardiac fibrosis and increased mortality. 26 Moreover, expression of the therapeutic protein in antigen presenting cells can give raise to an immune response against the therapeutic protein and/or transduced target cell. 27 In particular, because AAV can lead to long-term expression of its therapeutic payload, the restriction of expression to cardiomyocytes is of considerable importance for the future safe use of cardiac AAV gene therapy. One approach to suppress the ectopic expression of transgenes is the inclusion of miRNA target sequences for miRNAs that are expressed at high levels in the off-target organ but not in cardiomyocytes. That this is a promising approach has been demonstrated by Geisler et al. 28 who showed that the inclusion of three target sequences for mir122, which is expressed at very high levels in the liver but not the heart, into the 3′ untranslated region of the transgene mRNA essentially eliminated expression in the liver. 28 However, if ectopic expression occurs in more than one organ, this approach has its limitations because it requires the identification and inclusion of multiple different miRNA target sequences into the mRNA of the therapeutic transgene.
It is not surprising then that the development of expression cassettes that include cardiomyocyte “specific” promoters and other regulatory elements has been the focus by the authors and other research laboratories. 4 –9 The present study expands on work by French et al. who demonstrated that a minimal chicken cardiac TnT mediates high level expression of transgenes in the heart with significantly reduced nontarget organ expression compared to animals injected with an AAV9 vector with a CMV promoter expression cassette. 8 The present results demonstrate that the placement of a 192 bp long regulatory element from human calsequestrin 2, which was identified in previous work, 10 further enhances the specificity of cardiac transduction compared to the TnT promoter alone and mediates cardiac expression at levels that are equal to or higher than levels achieved with CMV promoter vectors. The study did not investigate whether the cardiac expression of luciferase from the Enh.TnT promoter construct is restricted to cardiomyocytes. However, because it has been shown previously by French et al. that the TnT construct is expressed in cardiomyocytes 8 but not endothelial cells or smooth muscle cells, and because AAV9 does not transduce cardiac fibroblasts, it is deemed unlikely that AAV9.EnhTnT.Luc leads to significant luciferase expression in cardiac cells other than cardiomyocytes.
It is noteworthy that the Enh.TnT regulatory element/promoter is of a similar size (621 bp) when compared to the CMV promoter (588 bp). Because of the limited packaging capacity of AAV vectors, 29 this is important for AAV gene therapy in general. However, in addition, the small size of the Enh.TnT regulatory element/promoter also allows the creation of self-complementary vectors, which have an even smaller packaging limit but trigger more rapid and robust transgene expression. 17,23 The enhanced specificity of the Enh.TnT vector (Figs. 2 –4 and Supplementary Fig. S1) can be observed in both adult animals and neonates (see above). The latter is important because the treatment of inherited cardiomyopathies with cardiac AAV gene therapy likely requires administration of the vector shortly after birth, or even in utero. Importantly, expression levels in adults are stable for at least 3 months (Fig. 3) and in neonates for a minimum of 4 weeks (Fig. 5), the longest time points tested. Whereas AAV9.Enh.TnT driven expression in the heart does not reach the same levels as those obtainable with the combination of the cis-regulatory motif element with the synthetic Spc-5-12 promoter, 10,11 expression in the liver and skeletal muscle was nearly completely eliminated. The precise mechanism of the reduction in ectopic expression is unknown, but it might involve the absence of transcription factors that bind to the specific sites in the cis-regulatory motif. Another, mutually non-exclusive, possibility is a repressor activity of the cis-regulatory motif in organs such as the liver and muscle.
Taken together, the data demonstrate that at the dose injected (5e 12 vg/kg) and when delivered by an AAV9 vector, the combination of a human calsequestrin 2 enhancer element 10 and a chicken cardiac TnT 8 mediates strong and highly cardiac-specific transgene expression. In fact, the levels of expression in the heart are equal to or higher than expression from both the TnT promoter alone as well as the strong, ubiquitous CMV promoter. The transduction pattern has not been tested when the transgene is delivered by another serotype (e.g., AAV1 or AAV6). However, given that the vector genomes would be identical, it is highly unlikely that the reduction of ectopic transgene expression with the Enh.TnT constructs would differ from the experiments with AAV9 described in this study.
The Calcium Upregulation by Percutaneous Administration of Gene Therapy in Cardiac Disease (CUPID) trial for the treatment of heart failure by delivering the sarcoplasmic calcium ATPase (SERCA2a) with an AAV1 vector was the first-in-human cardiac AAV gene therapy trial. 30 Unfortunately, the primary and secondary endpoints were not met in Phase II, most likely due to a failure of efficient gene delivery at the vector dose employed. 31 Hence, higher AAV vector doses are likely critical for the successful implementation of cardiac gene therapy. 31,32 The dose used in the present study (5e 12 vg/kg) would translate to a dose of 4e 14 vg in a patient who weighs 80 kg. This dose is 40-fold higher than the dose used in the CUPID2b trial. 31 The near absence of ectopic expression with the promoter/enhancer construct described here should allow the use of significantly higher vector doses than the one used in the CUPID trial, without increasing the risk of adverse effects resulting from nontarget organ expression. However, the possibly cannot be excluded that at doses even higher than the one used in this study, significant ectopic expression would also occur with the Enh.TnT constructs.
Taken together, it is important that this promoter–enhancer combination is tested in large-animal species used as model for cardiac AAV gene therapy and in nonhuman primates, including at doses (vg/kg) higher than the dose used in this study. If confirmed in nonhuman primates, the Enh.TnT combination described here shows great promise for future clinical cardiac gene therapy with AAV vectors.
Footnotes
Acknowledgments
We would like to thank Dr. Brent French for providing us with the pACMV-Luc and pAcTnT-Luc plasmids. This work was supported by NHLBI grants HL131404 (R.J.H. and T.W.) a Transatlantic Network of Excellence grant (14 CVD 03) by the Leducq foundation (T.W. and A.J.M.), HL088498 and HL132401 (A.J.M.), the George and Mary Josephine Hamman Foundation (A.J.M.), and VUB GEAR, VUB GROEIER, FWO (T.V. and M.C.).
Author Disclosure
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.