
Abstract
Invasiveness of growth hormone–producing pituitary adenomas (GHPAs) causes difficulties in safe and complete adenoma removal during surgery and often leads to high recurrence. Epidermal growth factor-like domain 7 (EGFL7) has been shown to be able to promote tumor angiogenesis, growth, invasiveness, and metastasis through the Notch signaling pathway. It was previously demonstrated that EGFL7 was overexpressed in GHPAs. This study reports that EGFL7 and Notch2 (positive correlation with EGFL7) are overexpressed in invasive GHPA. A long-rank test (Kaplan–Meier method) shows that invasive GHPAs with EGFL7 strong expression results in reduced recurrence-free survival. Multivariate Cox regression analysis reveals that weak EGFL7 expression is an independent prognostic factor for recurrence-free survival. In addition, knockdown of EGFL7 expression suppresses proliferation and invasion of GH3 and GT1-1 cells in vitro. Moreover, attenuation of EGFL7 inhibits human GHPA growth in vivo. The data suggest that as a Notch agonist, EGFL7 may potentially be an appropriate novel molecular target for future development of GHPA medical therapy.
Introduction
G
Epidermal growth factor-like domain 7 (EGFL7) is a secreted angiogenic signaling molecule involved in the regulation of blood vessel formation and cell migration, 5 –7 linked with many signaling cascades. 8 –11 Recent studies have found that EGFL7 expression is highly elevated in human tumors, 6,9,11 –18 and EGFL7 is capable of promoting tumor angiogenesis, growth, invasiveness, and metastasis through the Notch signaling pathway. 11,12,19 –23 It was previously demonstrated that EGFL7 is overexpressed in GHPAs. 24 However, the mechanisms underlying the tumorigenic effects of EGFL7 in GHPAs are not completely understood, and whether EGFL7 indeed promotes GHPA invasion remains to be determined.
This study evaluated the expression of EGFL7 and Notch2 in GHPAs, as well as the correlations of EGFL7 expression with tumor clinicopathologic characteristics and prognosis. Lentivirus-mediated knockdown of endogenous EGFL7 expression was used to investigate the role of EGFL7 and downstream Notch signal transduction pathway in GHPAs. A xenograft mouse model was also used to determine the effect of EGFL7 on in vivo tumor growth.
Materials and Methods
Patients and tissue specimens
GHPA tissue specimens were obtained from 48 patients who underwent GHPA resection by either endoscopic trans-sphenoidal surgery or craniotomy at the Department of Neurosurgery, Tiantan Hospital of Capital Medical University (Beijing, China) from April 2011 to December 2014. GHPAs were diagnosed according to clinical symptoms, radiologic features, hormonal levels, expression of transcriptional factor Pit-1, and electron microscopic examination. Patients with previous radiation therapy, medical therapy, or tumors with plurihormonal expression were excluded from this study. Included were 21 males and 27 females, with a mean age of 38.19 ± 12.32 years (range 13–64 years; median 38 years). The mean tumor volume was 5.24 ± 7.98 cm3 (range 0.09–48.67 cm3; median 3.09 cm3), and the mean preoperative serum GH was 23.75 ± 13.77 ng/mL (range 5.47–40 ng/mL; median: 21.7 ng/mL). All the specimens were fixed for tissue microarray (TMA) construction and immunohistochemistry (IHC). These samples were divided into two groups: the noninvasive GHPA group (n = 23) and the invasive GHPA group (n = 25). Invasive pituitary adenomas (PAs) are defined as Hardy–Wilson grade IV and/or Knosp grade III and IV. 4 Ten samples of normal pituitary glands obtained from a donation program were also included as a control. The donors included six men and four women aged 21–45 years (M age = 35 years). Another eight cases of GHPA specimens were obtained from November 2016 to Match 2017 for Western blot verification. This work was performed in accordance with the Code of Ethics of the World Medical Association (Declaration of Helsinki) for experiments involving humans. Prior informed consent was obtained for experimentation with human subjects. The privacy rights of human subjects were observed, and the study protocol was approved by the Ethics Committee of Beijing Tiantan Hospital.
Follow-up and prognostic study
Follow-up data were obtained after GHPA resection for all 48 patients. The follow-up period was defined as the interval between the date of operation and that of the patient's recurrence or the last follow-up. Recurrence was diagnosed by clinical examination, serial GH measurement, and magnetic resonance imaging (MRI) scans compared to the postoperative MRI scans within 72 h and confirmed histologically. The recurrence was 10/25 in invasive GHPAs but only 3/23 in noninvasive GHPAs. Five conventional variables together with EGFL7 expression were tested in all 48 patients: sex, age, tumor volume, preoperative serum GH level, and tumor invasiveness.
TMA construction and IHC
Formaldehyde-fixed, paraffin-embedded tissue blocks were sliced and stained with hematoxylin and eosin (H&E). H&E staining was performed using a standard peroxidase-based staining method according to the manufacturer's instructions (Beyotime Biotechnology, Haimen, China). Three core biopsies with diameter of 2.0 mm were selected from each block. These cores were then transferred to TMA using the Minicore Tissue-Arraying Instrument (Mitogen, Alphelys, France). TMAs were built according to an online protocol (
IHC analysis with mouse monoclonal anti-EGFL7 antibody (1:200; 2H2 sc-101349; Santa Cruz Biotechnology, Santa Cruz, CA) and rabbit polyclonal anti-Notch2 (1:500; ab8926; Abcam, Cambridge, MA) were performed on the sections using Leica BOND-III (Leica Biosystems, Bensheim, Germany), an automated, random, and continuous-access slide staining system. The primary antibodies were detected by Bond Polymer Refine Detection System (Leica Biosystems), and the expression of immunostained slides were examined with an Aperio AT2 digital scanner (Leica Biosystems). An H-score was used to evaluate the percentage of immunostaining and the staining intensity (0, negative; 1+, weak; 2+, moderate; and 3+, strong), and the formula for calculating the H-score was as follows: H-score = (% cells 1+) + 2(% cells 2+) + 3(% cells 3+). Based on the H-score, EGFL7 staining in tissue sections was categorized as weak (H-score ≤80.5) and strong (H-score >80.5).
Lentiviral transduction
Gene-specific shRNA expression vectors and non-effective shRNA cassette in pGFP-C-shLenti plasmid were purchased from Origene Technologies (Beijing, China). The sequence for EGFL7 targeting is TGTTGATGAATGCAGTACAGGAGAGGCCA. In previous studies, it was found that EGFL7 expression could be efficiently downregulated by this gene-specific shRNA. 24 A lentiviral transduction system with this targeting sequence was constructed. HEK293 cells in a subconfluent culture condition (70–80% confluency) were transfected with a third-generation lentiviral vector system, consisting of the packaging plasmids (pMDL, pREV, and pVSVG) and the transfer vector containing either gene-specific shRNA or non-effective shRNA cassette using polyethylenimine (2 mg/mL; Polysciences, Warrington, PA). After 2 days of culture, the cell supernatant was collected, filtered, and centrifuged at 16,000 g for 2 h at 4°C to concentrate viral particles by reducing half of the initial volume. The concentrated viral suspension was used to infect subconfluent cultures of GH3 or GT1-1. Polybrene (8 mg/mL; Sigma–Aldrich, St. Louis, MO) was used to neutralize the cell surface charge and increase the efficiency of infection. Cells with EGFL7 knocked down were named as follows: GH3DE7 or GT1-1DE7 for GH3 or GT1 cells infected with the gene-specific shRNA plasmid; GH3SCR or GT1-1SCR for GH3 or GT1 cells infected with non-specific scrambled vector plasmid.
Quantitative reverse transcription polymerase chain reaction
Total RNAs were extracted from cell cultures using the RNeasy 96 QIAcube HT kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. Messenger RNAs were reverse transcribed using a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA) following the manufacturer's specifications. Gene expression was measured using Platinum® SYBR® Green qPCR SuperMix-UDG w/ROX (Thermo Fisher Scientific). Quantitative reverse transcription polymerase chain reaction (RT-PCR) was performed using Applied Biosystems 7500 Fast System (Life Technologies, Carlsbad, CA). Differences among gene expression were calculated using the comparative CT method (2–ΔΔCT method). Specific primers sequences are listed below: EGFL7 (forward, TCGTGCAGCGTGTGTACCAG; reverse, GCGGTAGGCGGTCCTATAGATG) and GAPDH (forward, ACCACAGTCCATGCCATCACT; reverse, GTCCACCACCCTGTTGCTGTA).
Western blot analyses
Specimens were lysed in radio immunoprecipitation assay (RIPA) lysis buffer (50 mM pf Tris, pH 7.5; 250 mM of NaCl; 10 mM of EDTA; 0.5% NP-40; 1 μg/mL of leupeptin; 1 mM of PMSF; and 4 mM of NaF; Sigma–Aldrich) containing protease and phosphatase inhibitor cocktails (F. Hoffmann-La Roche Ltd., Basel, Switzerland). Lysates were centrifuged at 13,000 g for 10 min at 4°C, and then the protein concentration was determined with a bicinchoninic acid assay protein assay kit (Pierce Biotechnology, Rockford, IL). For Western blot analysis, 40 μg of protein samples per lane were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis using 10% running gels and blotted onto polyvinylidene fluoride membranes. Membranes were incubated with primary antibody overnight at 4°C, including rabbit polyclonal anti-EGFL7 antibody (1:500; 19291-1-AP; Proteintech, Wuhan, China), mouse monoclonal anti-EGFL7 antibody (1:2,000; 2H2 sc-101349; Santa Cruz Biotechnology), rabbit polyclonal anti-Notch2 (1:2,000; ab8926; Abcam), and GAPDH (1:8,000; Sigma–Aldrich) followed by secondary antibodies tagged with horseradish peroxidase (Santa Cruz Biotechnology). The membranes were visualized by enhanced chemiluminescence, and densitometry was performed with an Amersham Imager 6000 (GE, New York, NY).
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay (ELISA) was performed to quantify secreted EGFL7 levels using a rat EGFL7 ELISA kit (Rapid™; Biosensis, Temecula, CA) and mouse EGFL7 ELISA kit (Rapid™; Biosensis), respectively. The EGFL7 secretions from GH3 and GT1-1 cells were measured according to the manufacturer's protocol. The color intensity of the reaction product (which is proportional to the concentration of EGFL7) was measured using a multi-detection microplate reader (Tecan Infinite® M200 pro; Tecan Group AG, Männedorf, Switzerland). The concentration of EGFL7 was calculated using a straight-line regression equation of the standard curve with the standard density and the optical density value.
Cell proliferation and invasion assay
Proliferation of GH3DE7, GH3SCR, and GH3 cells or cells treated with antibodies against EGFL7 was assessed by MTT assay. Cells were cultured in 96-well plates. The dye and solubilization solutions (MTT Cell Proliferation Assay; Promega, Madison, WI) were added, and absorbance was measured at 490 nm with a Spectra Rainbow plate reader (Tecan Group AG). Tumor cell invasion assays were carried out using Transwell® chambers (8 μm pore size; Corning Costar Corp., Cambridge, MA) with Matrigel (50 μg/mL; BD Biosciences, Bedford, MA) in 24-well culture plates (BD Biosciences) according to the manufacturers' protocols. GH3DE7/GT1-1DE7, GH3SCR/GT1-1SCR, GH3/GT1-1, and GH3/GT1-1 cells with antibodies against EGFL7 (105 cells/ well) in serum-free Dulbecco's modified Eagle's medium were seeded in the upper chambers. After 18 h of incubation, cells adhering to the lower membrane were fixed in 4% paraformaldehyde (PFA) and stained using Harris hematoxylin. The average number of migrated cells was quantified by counting five random high-power fields (200 × ) under a phase-contrast microscope.
Xenograft mouse model
Six-week-old male athymic immunodeficient nude mice (SCXK2012-0001; Vital River Laboratory Animal Technology Co. Ltd, Beijing, China) were bred in an aseptic condition according to standard guidelines. Ten mice were divided into two groups, with five animals in each group. Five mice in the experimental group were inoculated subcutaneously on the hind flank with GH3DE7 cells (2 × 106 cells), and five mice in the control group were inoculated with GH3SCR cells (2 × 106 cells) on day 0. The size of the tumors was blindly measured every 4 days with calipers, and the tumor volume was calculated using the following formula: volume (mm3) = (length × width2) × 0.5. Mice were sacrificed 4 weeks after tumor cell inoculation, and the tumors were excised for further analysis. All animal experiments were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory animals (National Institutes of Health Publications No. 8023, revised 1978) and approved by the Animal Care and Use Committee of Beijing Neurosurgical Institute (Protocol KY2013-015-2).
Histoimmunofluorescence
All animals were perfused with 4% PFA after deep anesthesia, and the tumors were removed. The tumor samples were placed in 4% PFA overnight at 4°C, embedded in OCT compound, and frozen by n-hexane (−80°C) after being processed. Frozen horizontal sections (10 μm) were cut by a cryostat microtome (Leica Biosystems). Sections were washed three times with 0.01 M of phosphate-buffered saline and then incubated with the primary rabbit polyclonal anti-EGFL7 antibody (1:100; 19291-1-AP; Proteintech) and rabbit polyclonal anti-Notch2 (1:200; ab8926; Abcam) overnight at 4°C, followed by incubation with donkey anti-rabbit immunoglobulin G (1:1,000; ab150073; Abcam) for 2 h at 4°C. The sections were mounted with coverslips and mounting medium with DAPI (ZSGB-BIO, Beijing, China). Images were captured using a Laser Scanning Confocal Microscope (Leica Microsystems).
Statistical analysis
All experiments were performed at least three times, and all data were expressed as the mean ± standard deviation (SD). Kaplan–Meier analysis and log-rank tests were used to evaluate the prognostic significance of EGFL7 expression level for patient survival. The Cox proportional hazards regression model was also used to evaluate independent prognostic factors for GHPA samples with different levels of EGFL7 expression. One-way analysis of variance (ANOVA) was used to test for differences among at least three groups, and a least significant difference post hoc test was used to obtain individual p-values, followed by ANOVA. The t-test was used to determine differences between two groups. Asterisks indicate the level of statistical significance (*p < 0.05; **p < 0.01).
Results
Overexpression of EGFL7 and Notch2 in invasive GHPAs
To examine EGFL7 and Notch2 expression in GHPAs, TMA from all 48 cases of GHPA were constructed, and the expression levels were measured by IHC. The results showed that the average expression level of EGFL7 protein in invasive GHPA was about two fold of that in noninvasive GHPA, and the average expression level of Notch2 protein was more than 1.7-fold. There was a positive correlation between EGFL7 and Notch2 expression. Meanwhile, the EGFL7 and Notch2 expression in both invasive and noninvasive GHPAs were significantly higher than that in normal pituitary glands (p < 0.05; Fig. 1). To confirm the EGFL7 and Notch2 expression levels further, quantitative RT-PCR (Fig. 2E and F) and Western blotting (Fig. 2A and C) were used to examine their expression in additional four cases of invasive GHPAs and four cases of corresponding noninvasive GHPAs, and the results were consistent with the IHC results.
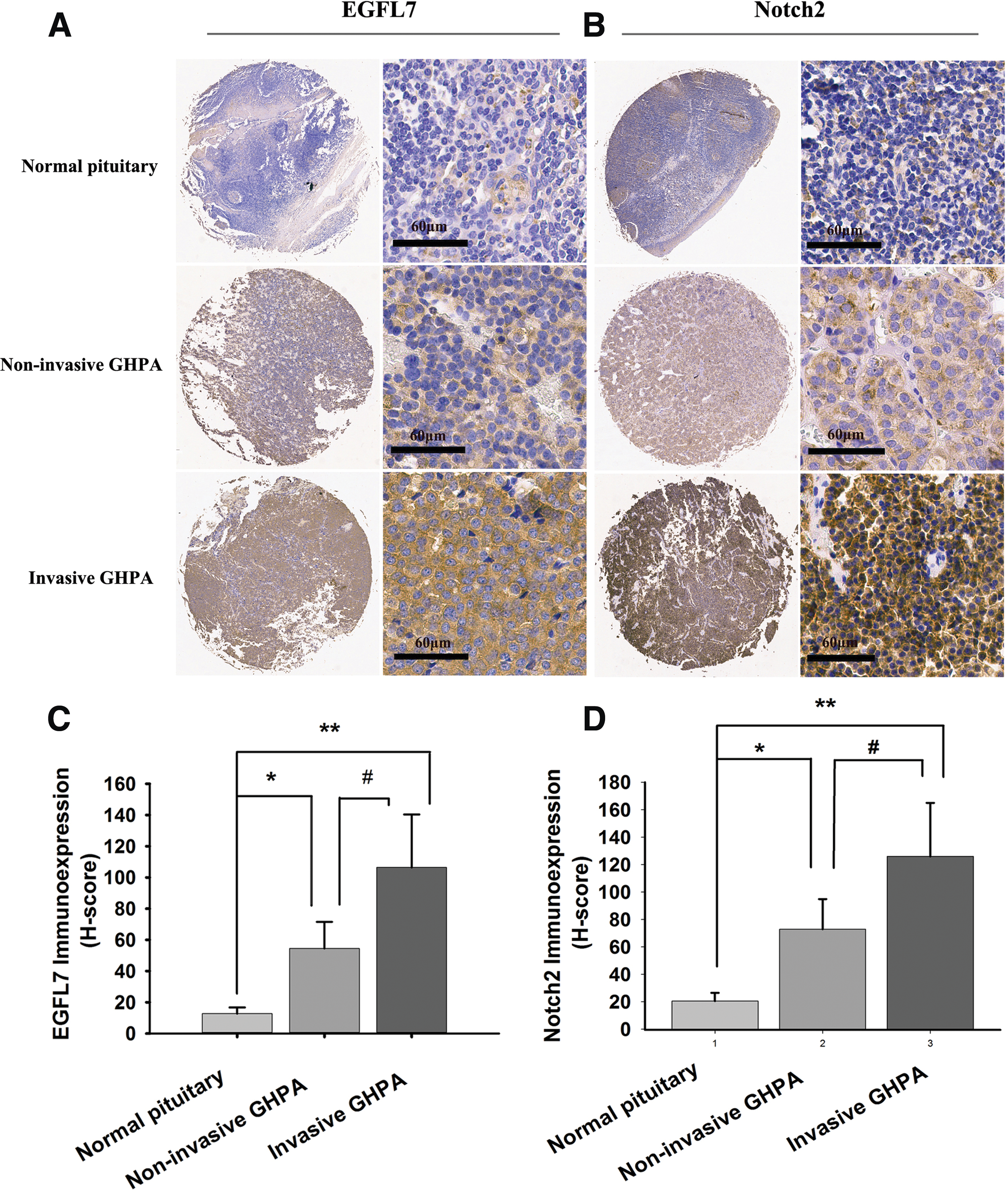
Overexpression of epidermal growth factor-like domain 7 (EGFL7) and Notch2 in growth hormone–producing pituitary adenomas (GHPA) tissues. Representative EGFL7 staining of a tissue microarray (TMA) showed that cytoplasmic EGFL7
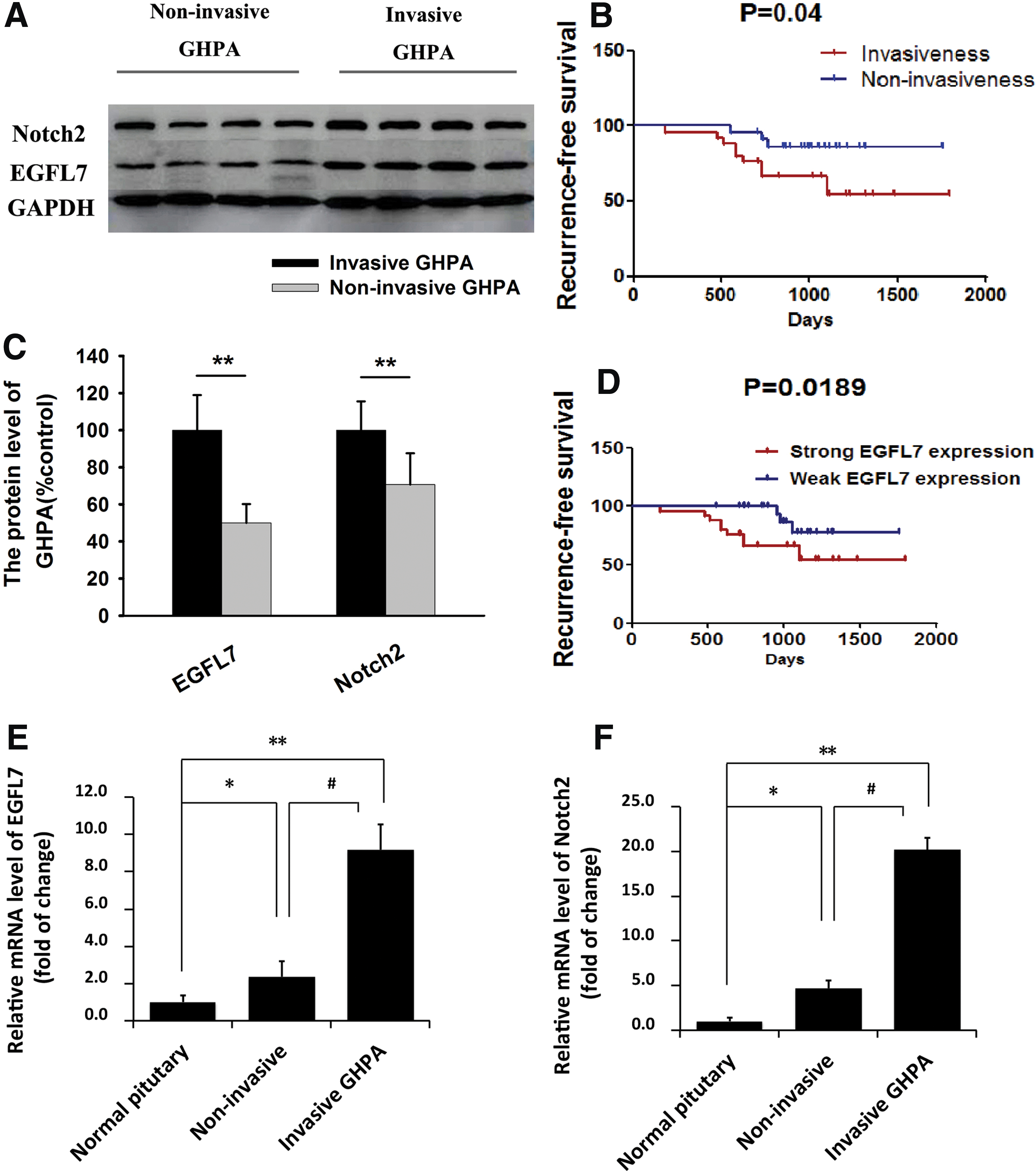
Representative Western blots and mRNA expression of EGFL7 in GHPA tissues and its prognostic implication.
Correlation of EGFL7 expression with clinicopathologic characteristics and prognosis of GHPAs
Detailed clinical characteristics of all 48 GHPA cases can be found in a previous study. 24 It was previously found that patient age, tumor volume, and histologic subtypes, but not sex and preoperative serum GH level, had significant impacts on invasiveness of GHPA by univariate analysis. 24 Recurrence-free survival related to invasiveness of GHPAs was estimated by the Kaplan–Meier method. The log-rank test shows that the invasive GHPA group had worse recurrence-free survival than the noninvasive GHPA group (Fig. 2B). Based on the H-score of IHC, EGFL7 staining in tissue sections was categorized as weak (H-score ≤80.5) and strong (H-score >80.5). GHPA patients with strong EGFL7 expression had shorter recurrence-free survival than those with weak expression (Fig. 2D). By multivariate Cox regression analysis, weak EGFL7 expression (relative risk = 0.159; p = 0.035) was found to be an independent prognostic factor for recurrence-free survival (Table 1).
Univariate and multivariate analyses for the clinicopathologic correlates of recurrence/progression-free survival
RR, relative risk; CI, confidence interval; GH, growth hormone; EGFL7, epidermal growth factor-like domain 7.
Suppression of EGFL7 expression on proliferation and invasion of GH3 and GT1-1 cells via the Notch signaling pathway
To investigate the function of EGFL7 in GHPA cells, lentivirus-mediated knockdown was used to reduce endogenous EGFL7 expression. Western blotting showed an approximate 60% reduction in EGFL7 protein expression in GH3 cells infected with the lentivirus knockdown vector for EGFL7 (GH3DE7) compared with the scramble vector infected cells (GH3SCR; Fig. 3A and B). Quantitative RT-PCR analysis demonstrated a 90% reduction in EGFL7 mRNA expression in GH3DE7 compared to GH3SCR (Fig. 3C). MTT and Transwell® assays were used to confirm the involvement of EGFL7 in the proliferation and invasion of GH3 cells. MTT assay revealed that the proliferation rate of GH3DE7 cells was significantly decreased compared to GH3SCR (Fig. 3D). Using a Transwell® assay, it was observed that the invasion efficiency of GH3DE7 cells was decreased by approximately 70% compared to that of GH3SCR cells (Fig. 3H). Similar results were obtained from another cell line, GT1-1. In this cell line, the EGFL7 mRNA (Fig. 3G) and protein (Fig. 3E and F) expression were significantly reduced by infection with the EGFL7 shRNA vector (GT1-1DE7) compared to that in the scrambled vector infected cells (GT1-1SCR). The transmembrane invasion assay revealed that knocking down EGFL7 significantly suppressed GT1-1 cells invasion (Fig. 3I), which was consistent with the findings in GH3 cells.
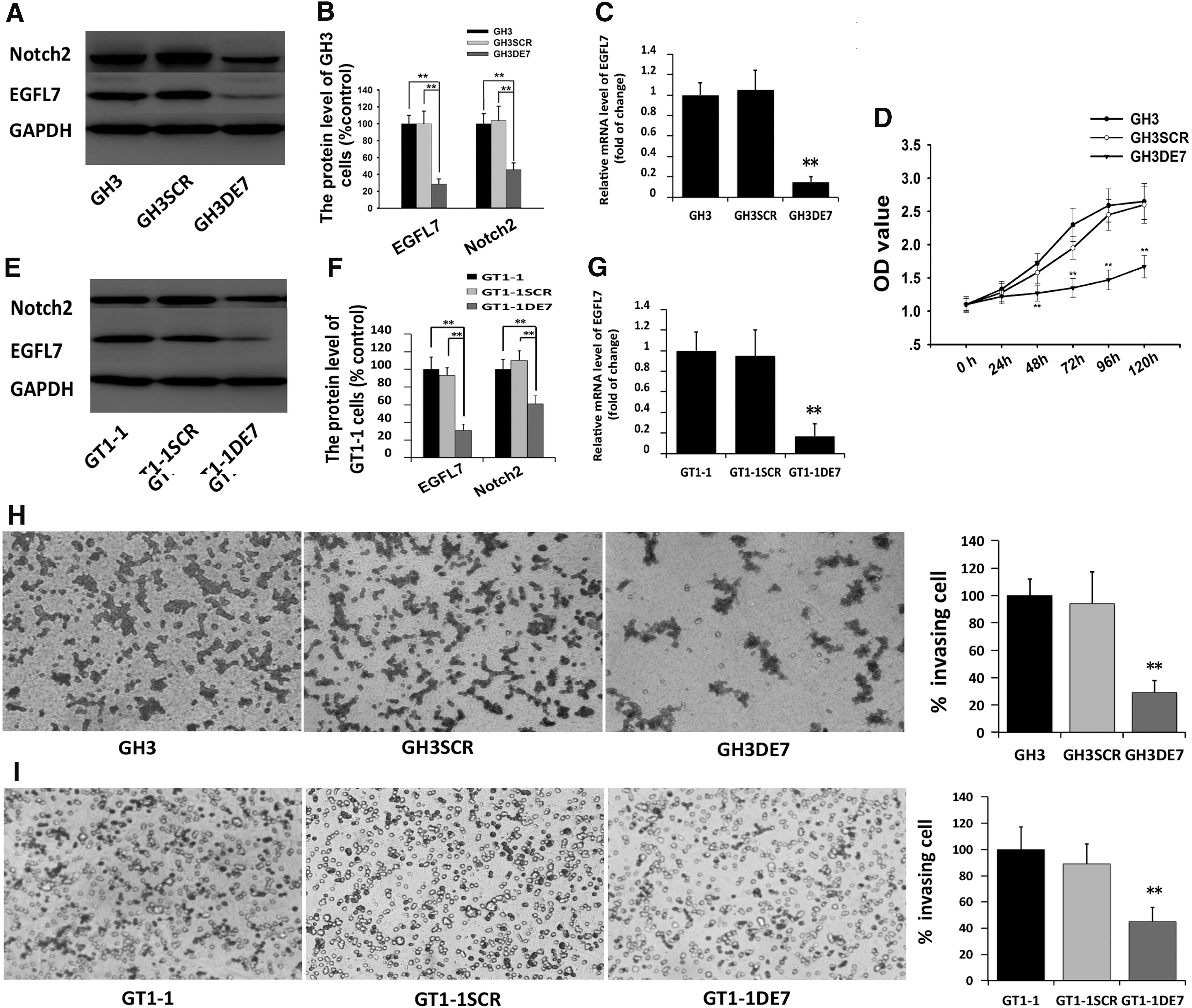
Lentivirus-mediated knockdown of endogenous EGFL7 expression exerted an effect on the biological characteristics of GH3 cells.
Previous studies showed that EGFL7 participates in regulating GH3 cell invasion and proliferation by interacting with Notch2. 24 Therefore, the expression of the Notch2 proteins was assessed in GH3 and GT1-1 cells. Using immunoblot analysis, expression of Notch2 was detected in both GH3DE7/GT1-1DE7 and GH3SCR/GT1-1SCR cells (Fig. 3A, B, E, and F).
EGFL7-blocking antibody impaired proliferation and invasion of GH3 and GT1-1 cells
The level of EGFL7 in the culture media from GH3 and GT1-1 cells was also measured by ELISA. The secretion of EGFL7 was increased in both GH3 and GT1-1 cells in a time-dependent manner (Fig. 4A and B). Next, experiments were performed with an antibody against the EGFL7 protein to block its function. Treatment of GH3 with 5 and 10 μg/mL of anti-EGFL7 antibody resulted in a significant decrease in the viability of both GH3 (Fig. 4C) and GT1-1cells (Fig. 4D) in 72 h. Furthermore, treatment of GH3 and GT1-1 cells with increasing concentrations of the anti-EGFL7 antibody resulted in a marked decrease in invasion efficiency assessed by the Transwell® assays (Fig. 4E and F).
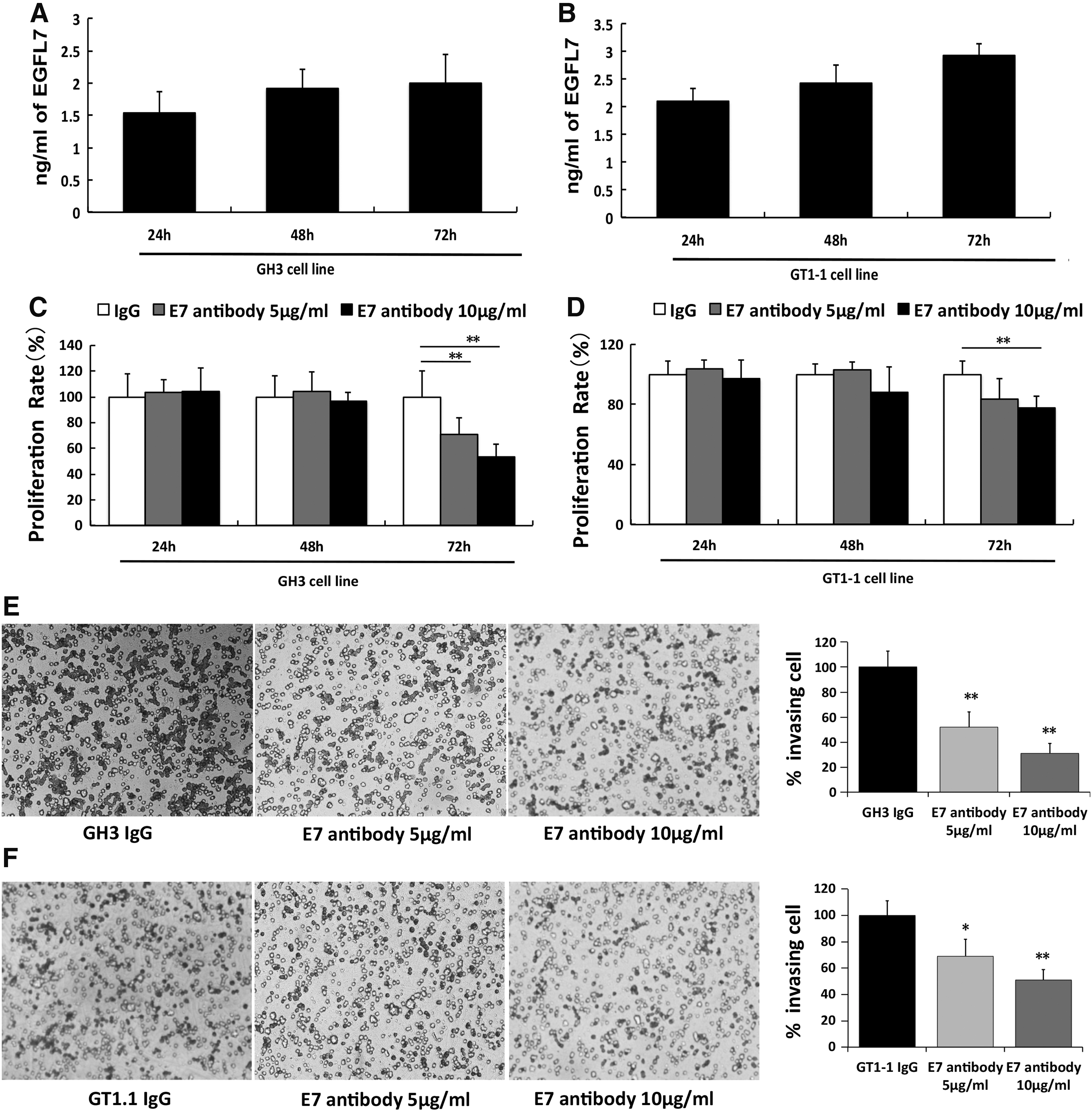
Anti-EGFL7 antibody decreased GH3 and GT1-1 cell proliferation and invasion. Both GH3
Downregulation of EGFL7 inhibits tumor growth in vivo
To validate the observations obtained from in vitro studies, the potential role of EGFL7 in tumorigenesis in vivo was examined using the GH3 xenograft mouse model. It was found that the average size and weight of local tumors in the GH3DE7 group were significantly smaller than those in the GH3SCR group (Fig. 5A–C). In addition, histoimmunofluorescent microscopy showed that the expression of EGFL7 and Notch2 was significantly decreased in the GH3DE7 group compared to that in the GH3SCR group (Fig. 5D). These results demonstrated the knockdown of EGFL7 inhibited GH3 tumor growth via the Notch signaling pathway.
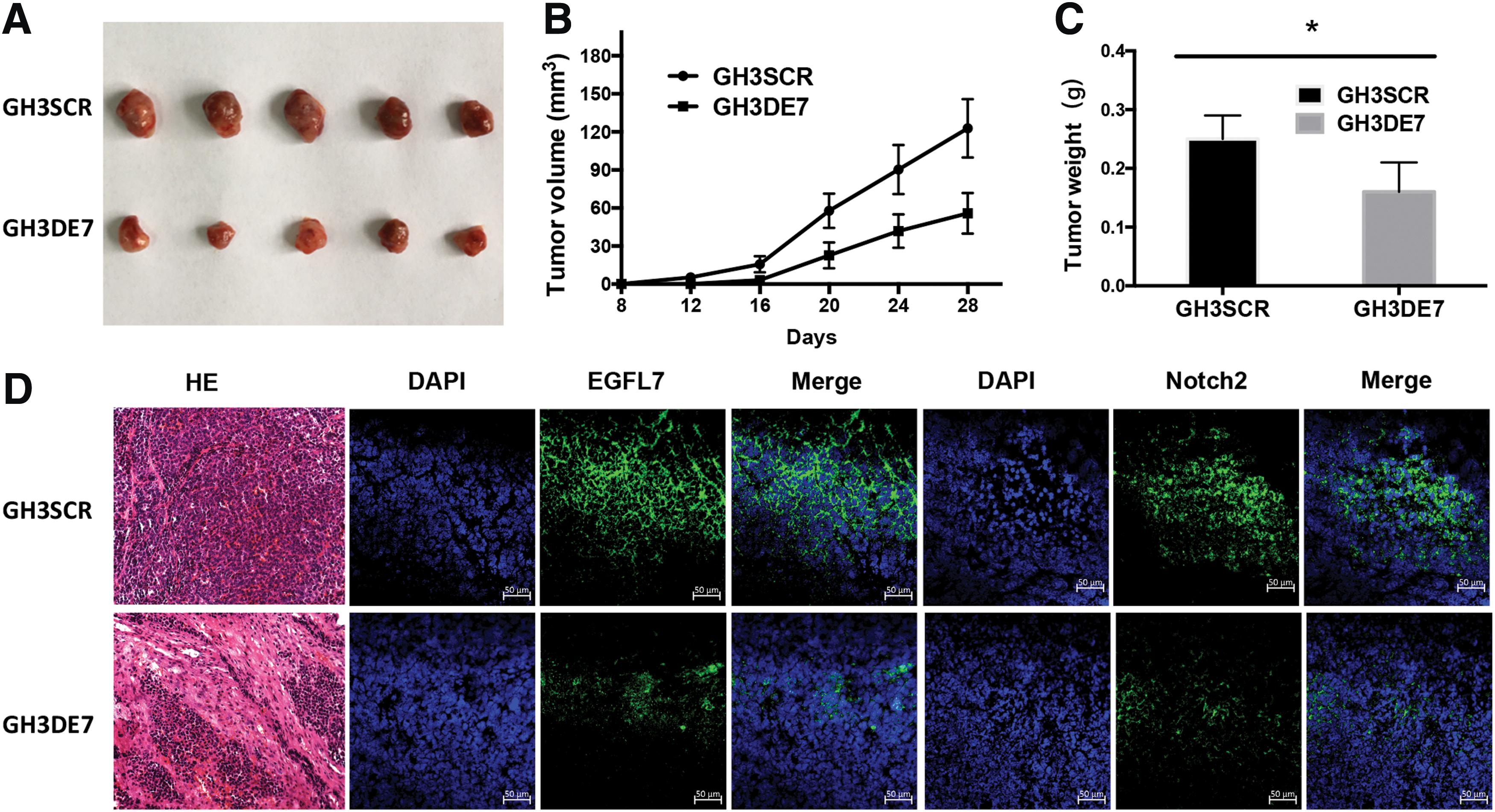
Downregulation of EGFL7 inhibits the growth of GHPA in vivo.
Discussion
Human PAs represent 10–15% of intracranial neoplasms, 25 and GHPAs constitute about 20% of all PAs. 26 The clinical manifestations of GHPAs depend on the hormone secretion, as well as on the growth within and through the sella turcica. GHPAs are often considered to be benign tumors. 1 However, about 33% of these tumors invade the surrounding tissues, including the sphenoid sinus, the cavernous sinus, and even the brain. 2,3 The current treatment for GHPAs includes trans-sphenoidal surgery, radiotherapy, medications, or a combination of these, but these methods are not always successful. Surgical resection is the first choice of treatment for most GHPAs. However, complete resection of GHPAs is unlikely when there is extensive local invasion. Additionally, surgeries for invasive GHPAs carry increased risks of complications. Invasion of GHPAs has been shown to be the most common cause of incomplete tumor resection and tumor recurrence. Radiotherapy or medications after incomplete tumor resection are often needed. Not all residual tumors progress or produce symptoms, and the response to treatment also varies among patients. However, GHPAs with invasive behavior may benefit from these adjuvant treatments, despite apparently complete removal. 27 Compared to noninvasive PAs, pituitary tumors that invade the cavernous sinus show significantly higher proliferation rates. 4 It was previously found that patient age, tumor volume, and histologic subtypes, but not sex and preoperative serum GH level, had significant impacts on the invasiveness of GHPAs. 24 In this study, a log-rank test shows that patients with invasive GHPAs has a shorter recurrence-free survival than patients with noninvasive GHPAs.
EGFL7, also known as vascular endothelial statin, is a secreted, extracellular matrix–bound protein that is highly conserved among species. EGFL7 expression is mostly restricted to the endothelium and is implicated in the regulation of blood vessel formation and cell migration. 5 –7 The EGFL7 protein sequence consists of a putative amino-terminal signal peptide domain, an EMI-like domain, and two centrally located EGF-like domains. 5 –7 EMI and both EGF-like domains of EGFL7 are necessary and sufficient for Notch binding. 28 Recent studies have shown that EGFL7 expression is highly elevated in human tumors, including kidney tumors, malignant gliomas, hepatocellular carcinomas (HCCs), colon cancers, breast cancer, oral squamous cell carcinoma, osteosarcoma, pancreatic carcinoma, and ovarian cancer. 6,9,11 –18 It was previously demonstrated that EGFL7 is overexpressed in GHPAs, and the expression was significantly correlated with pathologic characteristics, clinical progression, poor prognosis, and invasion. 24 The present study we found that GHPA patients with higher EGFl7 expression had reduced recurrence-free survival, and weak EGFL7 expression was an independent prognostic factor for recurrence-free survival. EGFL7 has been shown to be able to promote tumor angiogenesis, growth, invasiveness, and metastasis via the Notch signaling pathway. 11,12,19 –23 This study confirms that EGFL7 is involved in promoting tumor cell invasion and proliferation via the Notch signaling pathway.
The identification of particular molecular markers may provide potential targets for the future development of novel treatment options. Several studies have shown the benefits of blocking Notch signaling in tumor models 22,29 –31 , and several preclinical studies and clinical trials have been conducted to evaluate the effect of anti-EGFL7 therapy on tumor treatment. 32 This study demonstrates that the attenuation of EGFL7 can inhibit GHPA growth via the Notch signaling pathway using GH3 xenograft mouse model. EGFL7 functions as a Notch agonist, which may become a potential molecular target for future GHPA medical therapy.
In conclusion, the present findings show that EGFL7 and Notch2 are expressed in GHPAs, and EGFL7 overexpression significantly correlates with a poor prognosis of GHPAs. Moreover, it demonstrates the critical role of EGFL7 in the proliferation and invasiveness of GH-secreting pituitary tumor cell lines via the Notch signaling pathway. Furthermore, attenuation of EGFL7 expression is able to inhibit GH3 tumor growth in vivo. Taken together, these data suggest that EGFL7 is a novel prognostic marker and a potential therapeutic target of GHPAs.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (81502154, 81602182), Natural Science Foundation of Shandong Province (ZR2016HP42, ZR2017MH077), Research Special Fund for Public Welfare Industry of Health of China (201402008), and National High Technology Research and Development Program of China (863 Program, 2015AA020504).
Author Disclosure
The authors declare that they have no conflicts of interest and that no competing financial interests exist.