
Abstract
Clustered regularly interspaced short palindromic repeats (CRISPR)-caspase 9 (Cas9) genome editing technology holds great promise for the field of human gene therapy. However, a lack of safe and effective delivery systems restricts its biomedical application. Here, a folate receptor-targeted liposome (F-LP) was used to deliver CRISPR plasmid DNA co-expressing Cas9 and single-guide RNA targeting the ovarian cancer–related DNA methyltransferase 1 (DNMT1) gene (gDNMT1). F-LP efficiently bound the gDNMT1 plasmid and formed a stable complex (F-LP/gDNMT1) that was safe for injection. F-LP/gDNMT1 effectively mutated endogenous DNMT1 in vitro, and then expressed the Cas9 endonuclease and downregulated DNMT1 in vivo. The tumor growth of both paclitaxel-sensitive and -resistant ovarian cancers were inhibited by F-LP/gDNMT1, which shows fewer adverse effects than paclitaxel injection. Therefore, CRISPR-Cas9-targeted DNMT1 manipulation may be a potential therapeutic regimen for ovarian cancer, and lipid-mediated delivery systems represent promising delivery vectors of CRISPR-Cas9 technology for precise genome editing therapeutics.
Introduction
T
DNA methyltransferase 1 (DNMT1) is a key nuclear enzyme of DNA methylation and an important molecular target for epigenetic therapy of cancer. 25 –27 The abnormal overexpression of DNMT1 inactivates tumor suppressor genes and is indispensable for the maintenance of cancer stem cells. 28,29 Moreover, DNMT1 closely correlates with tumorigenesis, relapse, and resistance of ovarian cancer, and high DNMT1 levels have been frequently observed in patients with poor prognoses. 30,31 Inconsistent with this observation, a series of studies have shown that inhibition of DNMT1 suppressed ovarian cancer growth and resistance. 32 –34 Consequently, DNMT1 is a promising molecular target of ovarian cancer treatment. 35,36 As a targeted genome editing technique, CRISPR-Cas9 has been used to edit endogenous genes in a wide variety of cell types and in organisms. 24,37,38 Therefore, using the CRISPR-Cas9 system to edit DNMT1 of ovarian cancer cells is a potential approach for ovarian cancer therapy.
Delivery systems, including lentiviral, adenoviral, adeno-associated viral, naked plasmid DNA, nanoparticles, and cationic liposomes, have been employed and are known to work well in CRISPR-Cas9-mediated research. 37 –39 However, the lack of safe and effective delivery systems restricts further application of CRISPR-Cas9 in cancer gene therapy. 40,41 To enable the application of Cas9 in vivo broadly, new delivery systems must be researched and developed. 42 Ideally, Cas9 delivery vehicles should be safe, induce little to no immune response, have a short half-life and few limitations when delivering larger genetic payloads, not integrate into the genome, and efficiently regulate target cell functions. 14,37,42 Cationic liposomes as a nonviral delivery system are easily modified in order to meet the requirements of an ideal Cas9 delivery vehicle. 42,43 Furthermore, cationic liposomes have been utilized to deliver Cas9:single-guide RNA (sgRNA) complexes into cultured human cells in vitro and a mouse inner-ear in vivo and efficiently modified the target genomes (80% in vitro, 20% in vivo). 44 Therefore, cationic liposomes are an available vector for CRISPR-Cas9 delivery.
In the present study, a folate receptor-targeted cationic liposome (F-LP) was developed to deliver a CRISPR plasmid expressing Cas9 and sgRNA targeting the ovarian cancer-related DNMT1 gene (gDNMT1). The physicochemical properties of the F-LP/gDNMT1 complex were characterized in vitro. Then, the ability of these complexes to edit the genome of ovarian cancer cells precisely was evaluated both in vitro and in vivo. Furthermore, using paclitaxel (PTX) as a positive control, the antitumor effects of F-LP/gDNMT1 were also explored in a mouse model of ovarian cancer.
Materials and Methods
Materials
The pX330 plasmid containing a human-encoded optimized Cas9 and sgRNAs was purchased from Addgene (pX330-U6-Chimeric_BB-CBh-hSpCas9; Addgene: 42230; Addgene, Cambridge, MA). 45 BbsI, T4 polynucleotide phosphorylase, and T4 ligase were obtained from Life Technologies (Carlsbad, CA). 1, 2-Dioleoyl-3-trimethylammonium-propane chloride salt (DOTAP) was purchased from Avanti Polar Lipids, Inc. (Alabaster, AL), and cholesterol (Chol) was obtained from Shanghai Bio Life Science & Technology Co. Ltd. (Shanghai, China). Methoxy-polyethylene glycol (PEG)-succinyl-Chol and folate-PEG-succinyl-Chol conjugates were synthesized and purified, as previously described. 46,47 F-LP was composed of DOTAP, Chol, methoxy-PEG-succinyl-Chol, and folate-PEG-succinyl-Chol and was prepared by film dispersion, as previously described. 48,49 Coumarin was incorporated into F-LP to prepare F-LP-coumarin for in vivo imaging. Plasmid DNA was extracted according to the EndoFree Plasmid Purification Handbook (Qiagen, Hilden, Germany). Pre-stained protein and DNA ladders were purchased from Fermentas (Thermo Fisher Scientific, Waltham, MA). Glucose (5%) and NaCl (0.9%) injections were obtained from Sichuan Kelun Pharmaceutical Co. Ltd. (Chengdu, China). PXT injection was purchased from Taiji Group Sichuan Taiji Pharmaceutical Co. Ltd. (Chengdu, China).
sgRNA design and Cas9 vector construction
For targeting DNMT1 with the pX330 Cas9 vector, sgRNA sequences (Table 1) were designed using online software (
gRNA sequences
eGFP, enhanced green fluorescent protein; DNMT1, DNA methyltransferase 1.
Preparation and characterization of lipoplexes
FR-targeted lipoplexes were prepared by mixing F-LP or F-LP-coumarin with plasmid DNA for 30 min at room temperature. The appearance of F-LP and lipoplexes were photographed with a digital camera (Canon, Tokyo, Japan). The mean particle size, polydispersion index, and zeta potential of lipoplexes were determined by a Zetasizer NanoZS ZEN 3600 (Malvern Instruments Ltd., Malvern, United Kingdom). Morphological characteristics of the F-LP/gDNMT1 complex were examined by a Tecnai G2 F20 transmission electron microscope (FEI Co., Hillsboro, OR). After lipoplexes were prepared, agarose gel electrophoresis was conducted in pH 7.4 Tris-base/acetic acid/EDTA buffer containing Gold View nucleic acid stain, as previously described. 49
A hemolysis study of F-LP/gDNMT1 complexes was performed in normal saline solution, with plasmid weights ranging from 2.5 to 15 μg. Briefly, lipoplexes with various plasmid weights were diluted in 2.5 mL of normal saline and added to 2.5 mL of rabbit erythrocyte suspension (2%) in normal saline at 37°C; normal saline and distilled H2O were used as negative and positive controls, respectively. The erythrocyte suspensions were centrifuged 3 h later, and the color of supernatants were compared to that of the controls. 51
Cell culture
SKOV-3 human ovarian cancer cells were obtained from American Type Culture Collection (Manassas, VA). The cells were cultured as a monolayer in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum in a humidified atmosphere containing 5% CO2 at 37°C. Human ovarian cancer PXT-resistant SKOV-3/TR cells were selected from SKOV-3 cells by step-wise increases in PXT concentration (0.1–300 nM). 52 An eGFP control lentivirus was used to infect SKOV-3 cells to generate eGFP-stable cells (SKOV-3eGFP+), and stable clones were selected by green fluorescence sorting to purify SKOV-3eGFP+ cells.
In vitro transfection experiments
In vitro transfection experiments were performed, as previously described. 53 F-LP/gDNMT1 and F-LP carrying CRISPR plasmid DNA co-expressing Cas9 and sgRNA targeting eGFP (geGFP) (F-LP/geGFP) were used to transfect SKOV-3 and/or SKOV-3eGFP+ cells. SKOV-3 cells transfected with F-LP/gDNMT1 were stained with an anti-DNMT1 antibody (1:100; Proteintech Group, Inc., Chicago, IL) and 4′,6-diamidino-2-phenylindole and photographed using a Leica TCS SP5 confocal laser-scanning microscope (Leica Microsystems CMS GmbH, Mannheim, Germany). After cellular nuclei were stained with Hoechst 33258, the transfection efficiency of F-LP/geGFP into SKOV-3eGFP+ cells was determined using an Olympus IX 70 inverted fluorescence microscope (Olympus Corp., Shinjuku, Tokyo, Japan) and FACSCalibur flow cytometry (BD Biosciences, San Jose, CA).
Target site (DNMT1) identification
Genomic DNA was prepared from SKOV-3 cells transfected with an empty CRISPR vector or CRISPR vectors harboring DNMT1 target sequences. DNA sequences containing each sgRNA target site were amplified by polymerase chain reaction (PCR). These PCR products were analyzed by T7E1 or restriction enzyme digestion, and the DNA was sequenced to screen for possible effective sgRNAs and quantify the indel efficiency. 54,55 The forward and reverse PCR primers for DNMT1 were 5′-TCTTCTGGAGAGTTGAAAGC-3′ and 5′-GGGCTTCTACCCGTTTACC-3′, respectively.
DNA dot blot
DNA dot blot was performed, as described by Blaschke et al. 56 Briefly, isolated genomic DNA was denatured in 0.1 M of NaOH for 10 min at 95°C, neutralized with 1 M of NH4CH3CO2 on ice, and then serially diluted twofold in TE buffer (10mM Tris-HCl 1mM EDTA PH 8.0). DNA samples were spotted on a nitrocellulose membrane and dried at 80°C for 5 min. The membrane was subjected to ultraviolet crosslinking at 120,000 μJ/cm2, blocked in blocking buffer (Tris-buffered saline/Tween 20 with 5% milk) at room temperature for 2 h, and then incubated with a mouse anti-5-methylcytosine monoclonal antibody (1:500; Active Motif, Carlsbad, CA) at 4°C overnight. After washing the membrane three times for 10 min each in Tris-buffered saline/Tween 20, it was incubated with sheep anti-mouse horseradish peroxidase-conjugated immunoglobulin G secondary antibody (1:2,000; ZSGB-BIO, Beijing, China) for 3 h at room temperature. The membrane was then washed three times for 10 min each in Tris-buffered saline/Tween 20 and visualized with a ChemiScope 5300 Integrated Chemiluminescence Imaging System (Clinx Science Instruments Co. Ltd., Shanghai, China).
Animal experiments
Athymic female nude mice (BALB/c, 4–6 weeks old) were purchased from Vital River Laboratories (Beijing, China). The mice were housed and maintained under specific pathogen-free conditions. All mice were handled according to the Guide for the Care and Use of Laboratory Animals (National Research Council, US), and all experiments performed on animals followed the guidelines and regulations of Sichuan University (Chengdu, China). All procedures were approved and supervised by the State Key Laboratory of Biotherapy Animal Care and Use Committee (Sichuan University, Chengdu, China).
Experiments with these mice began when they were 6–8 weeks old. For in vivo imaging, F-LP/gDNMT1 tagged by coumarin was injected into the abdominal cavity. After 6 or 12 h, the vital organs (heart, liver, spleen, lung, kidney, brain, stomach, and intestinal tract) were harvested and observed under a Bio-Real QuickView 3000 in vivo imaging system (Labatech GmbH, Salzburg, Austria). Tumor models were established by intraperitoneal (i.p.) injection of SKOV-3 or SKOV-3/TR cells (5 × 106 cells in 0.2 mL serum-free DMEM), and mice were randomly allocated into four groups. To assess tumor growth, treatment began 7 days after inoculation. Mice were administered i.p. every 3 days with liposomal plasmid DNA (2.5 or 5 μg) in 200 μL of glucose solution. Tumor development (n = 8 per group) were monitored every 3 days. Two days after the 10th treatment, mice were anesthetized and sacrificed. Tumor tissues were harvested and processed, as previously described. 48,57
Reverse transcription PCR
For reverse transcription (RT)-PCR, total RNA from tumor tissues, which were treated as indicated in the section “In vivo gene editing using F-LP/gDNMT1,” was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA) and quantified by spectrophotometry. cDNA was synthesized using a RevertAid First Strand cDNA Synthesis kit (Thermo Fisher Scientific, Rockford, IL) following the manufacturer's guidelines. The primers were designed as follows: Cas9, 5′-ACCATCGACCGGAAGAGGTA-3′ (forward) and 5′-CGATCCGTGTCTCGTACAGG-3′ (reverse); β-actin, 5′-AGCGAGCATCCCCCAAAGTT-3′ (forward) and 5′-GGGTGGCTTTTAGGATGGCA-3′ (reverse).
Western blot and hematoxylin and eosin staining
Total protein concentrations of tumor tissue lysates were determined with a Pierce BCA assay reagent kit (Thermo Fisher Scientific). DNMT1, Cas9, heterochromatin protein (HP)1α and p15INK4b proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis under reducing conditions and then transferred to polyvinylidene difluoride membranes (Millipore Corp., Billerica, MA). Membranes were blocked with 5% skim milk and incubated with anti-DNMT1 (Proteintech Group, Inc.), anti-Cas9, anti-HP1α, and anti-p15INK4b primary antibodies (Abcam, Boston, MA) at 4°C overnight. Antibodies were detected with a horseradish peroxidase-conjugated secondary antibody and developed with an enhanced chemiluminescence detection kit (Luminata Crescendo Western horseradish peroxidase substrate; Millipore Corp.); β-tubulin was used as a control. Tumor tissue sections were also stained with hematoxylin and eosin for histomorphometric analysis. All sections were observed or counted by two investigators or pathologists in a blinded fashion.
Statistical analysis
Statistical analysis was performed with one-way analysis of variance using IBM SPSS Statistics for Windows v19.0 (IBM Corp., Armonk, NY). Differences were considered statistically significance at p < 0.05.
Results and Discussion
In vitro reporter gene editing based on CRISPR-Cas9 system
Ovarian cancer SKOV-3 cells infected by lentivirus-encoded eGFP and eGFP-stable cells SKOV-3eGFP+ were purified after green fluorescence sorting. Fluorescence microscopy (Fig. 1a) and flow cytometric analysis (Fig. 1b) showed that eGFP expression was disrupted in about 70% of SKOV-3eGFP+ cells after F-LP/geGFP transfection. This in vitro genome modification efficiency was similar to that mediated by other cationic lipid/Cas9:sgRNA nuclease complexes. 44 These results indicate that F-LP is a potential nonviral vector for delivering the CRISPR-Cas9 system. Therefore, the CRISPR plasmid was further designed to target DNMT1, and F-LP was utilized to deliver the gDNMT1 plasmid and disrupt DNMT1 expression in ovarian cancer cells.
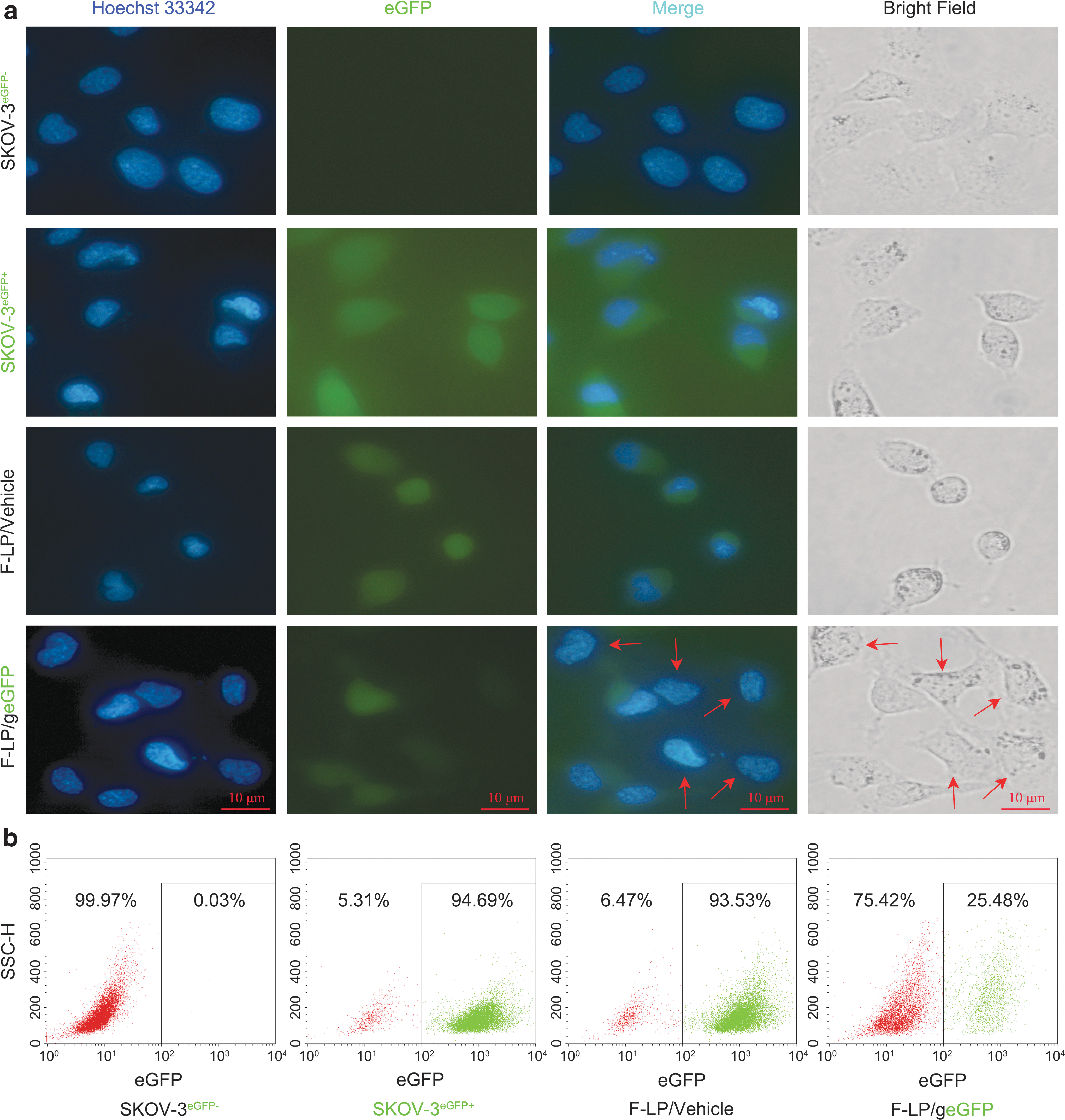
Targeted genome editing for reporter gene based on folate receptor-targeted liposome (F-LP) delivery.
Physicochemical properties of F-LP/gDNMT1 complexes
The gDNMT1 plasmid is large, with about 8,500 base pairs, and complete F-LP binding did not occur until the weight ratio between F-LP and gDNMT1 was 12:1 or 16:1 (Fig. 2a). The results showed that the lipoplexes (F-LP/vehicle and F-LP/gDNMT1) were formulated successfully and no free plasmid DNA was observed on the agarose gel (Fig. 2b). The appearances of the lipoplexes were clear under the light, similar to F-LP alone (Fig. 2c). The particle size and polydispersion index of F-LP/gDNMT1 were about 150 nm and 0.15, respectively (Fig. 2d and e). The morphology of F-LP/gDNMT1 was spheroidal or rod-like, and F-LP/gDNMT1 exhibited a membrane structure via transmission electron microscopy (Fig. 2f, classical morphology indicated by red arrows). The zeta potential of F-LP/gDNMT1 was about 35 mV, with a uniform distribution (Fig. 2g and h). The results of hemolytic tests demonstrated that F-LP/gDNMT1 did not cause hemolysis under test conditions (Fig. 2i). Overall, the F-LP/gDNMT1 complex displayed good physicochemical properties for injection. Therefore, F-LP/gDNMT1 was further evaluated for targeted genome editing in ovarian cancer cells in vitro.
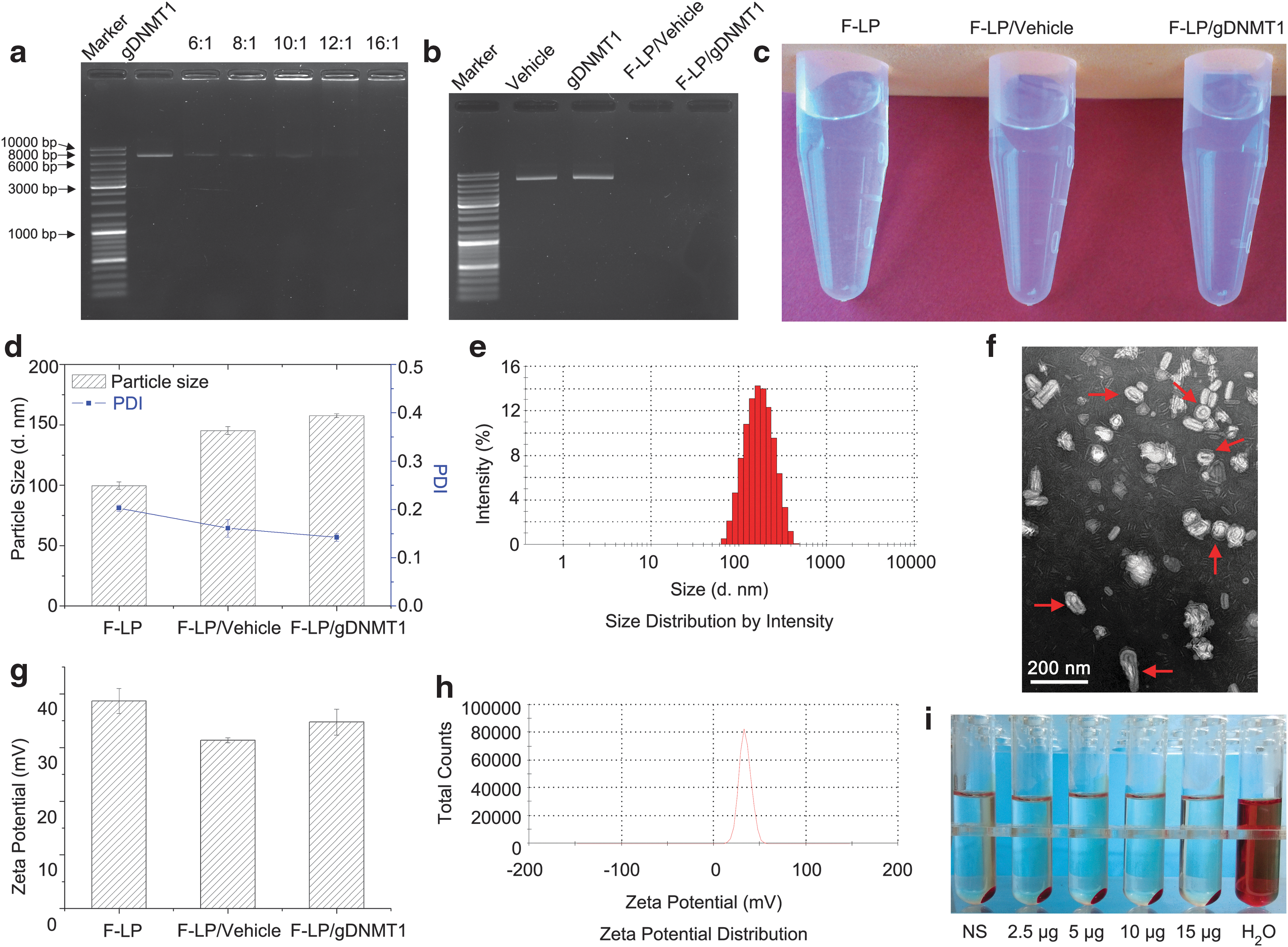
Preparation and characterization of F-LP/ DNA methyltransferase 1 (DNMT1) gene (gDNMT1) complexes.
In vitro targeted genome editing of endogenous DNMT1
Four gDNMT1 plasmids containing different sgRNAs were constructed (Table 1), and then four F-LP/gDNMT1 complexes were transfected into SKOV-3 cells to identify functional DNMT1 sgRNA. After transfection, PCR and T7E1 assay were performed to determine if there were indels in sgRNA target sites within DNMT1. One out of four sgRNAs (DNMT1-gRNA1; Table 1) were found to be effective, and the indel percentage was 28.6% (Fig. 3a). Moreover, the resultant mutations were further verified by Sanger sequencing (Fig. 3c). DNA dot blots indicated that 5-methylcytosine was downregulated after SKOV-3 cells were transfected with F-LP/gDNMT1 (Fig. 3b). Endogenous DNMT1 was disrupted, and DNMT1 expression was inhibited according to immunofluorescence (Fig. 3d). To conclude, F-LP effectively delivered CRISPR plasmid DNA to ovarian cancer cells, and the CRISPR-Cas9 system expressed in ovarian cancer cells efficiently mediated the targeted inactivation of DNMT1 in vitro.
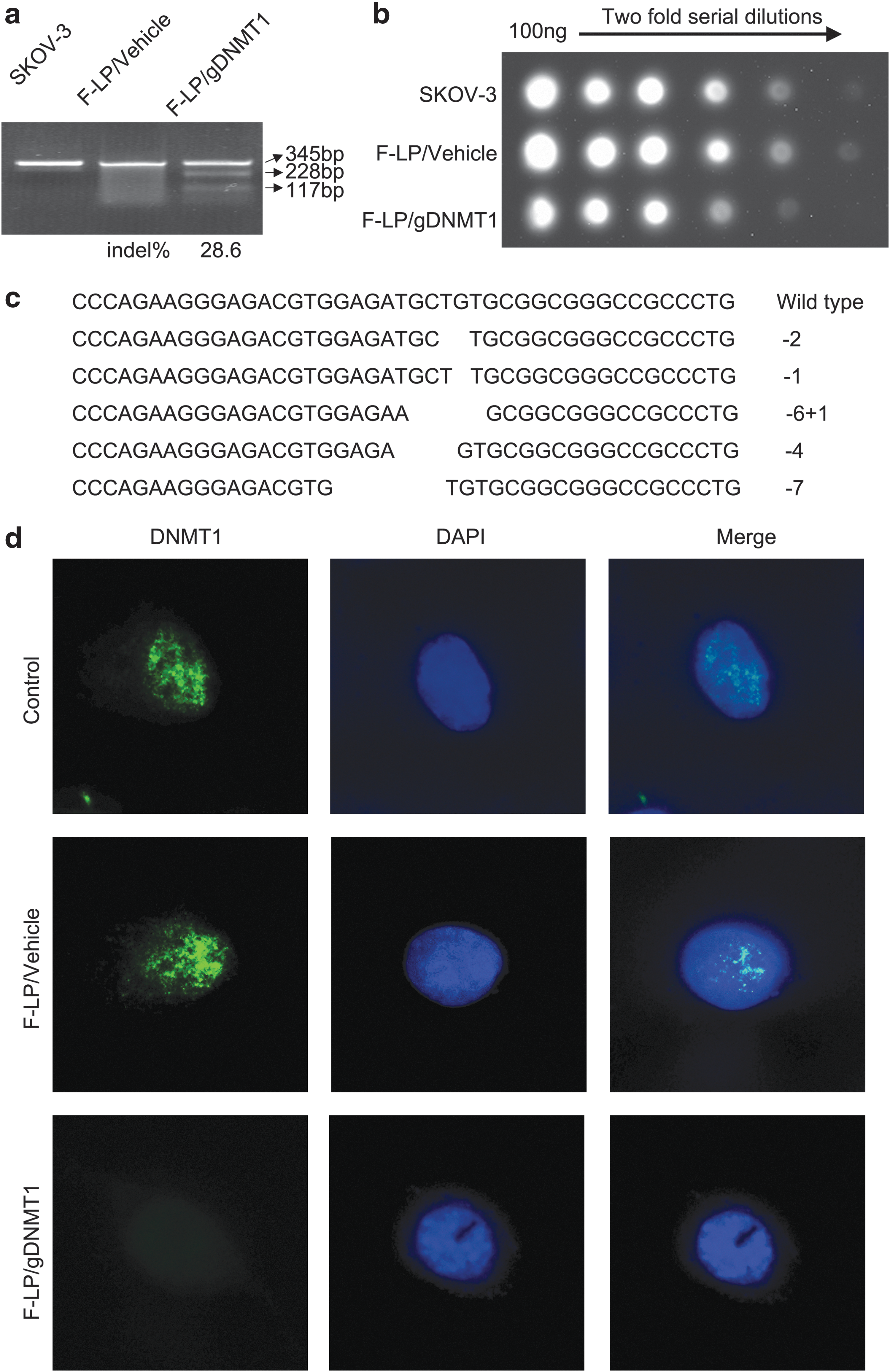
In vitro targeted genome editing in the endogenous DNMT1 gene.
In vivo distribution and antitumor effects of F-LP/gDNMT1
F-LP/gDNMT1 tagged by coumarin was injected into the peritoneal cavity of mice to explore the distribution of F-LP/gDNMT1 in vivo. As shown in Fig. 4a, coumarin-tagged F-LP/gDNMT1 sustained its high levels at mesenteric tissues along the intestinal tract 12 and 24 h after administration i.p. Due to higher peritoneal levels of F-LP/gDNMT1 and its retention, the antitumor effects of F-LP/gDNMT1 was similar to PTX (1.25 mg/kg) and better than that of F-LP/vehicle (p < 0.01; Fig. 4b and c). Compared to the negative control, F-LP/gDNMT1 had a tumor inhibition rate of 84% and significantly inhibited the growth of PTX-sensitive tumors (p < 0.001; Fig. 4b and c). Because 1.25 mg/kg of PTX had little antitumor effect on PTX-resistant tumors, the dose was increased to 10 mg/kg. Moreover, F-LP/gDNMT1 also inhibited the growth of PTX-resistant tumors, whereas F-LP/vehicle did not (p < 0.05; Fig. 4d). Furthermore, F-LP/gDNMT1 showed an approximate antitumor effect like high-dose PTX (10 mg/kg) and reduced the visible toxicity of PTX (10 mg/kg) (Fig. 4e). Cationic lipid and bacterial plasmid DNA complexes could elicit adaptive cellular immunity in murine i.p. tumor models, 58 thus cationic liposomes (containing cationic lipid DOTAP 50%, mole ratio) and plasmid DNA complexes induced cellular immunity and activated macrophages to inhibit tumor growth in BALB/c nude mice in this study. The tumor growth inhibition rate elicited by F-LP/vehicle was 53.6% (0.41 vs. 0.22 g; Fig. 4b) in PTX-sensitive tumors and 45.9% (0.61 vs. 0.28 g; Fig. 4d) in PTX-resistant tumors compared to F-LP/gDNMT1 (100% in both tumor types). Thus, F-LP/gDNMT1 is a novel and promising antitumor therapeutic agent based on gene editing technology.
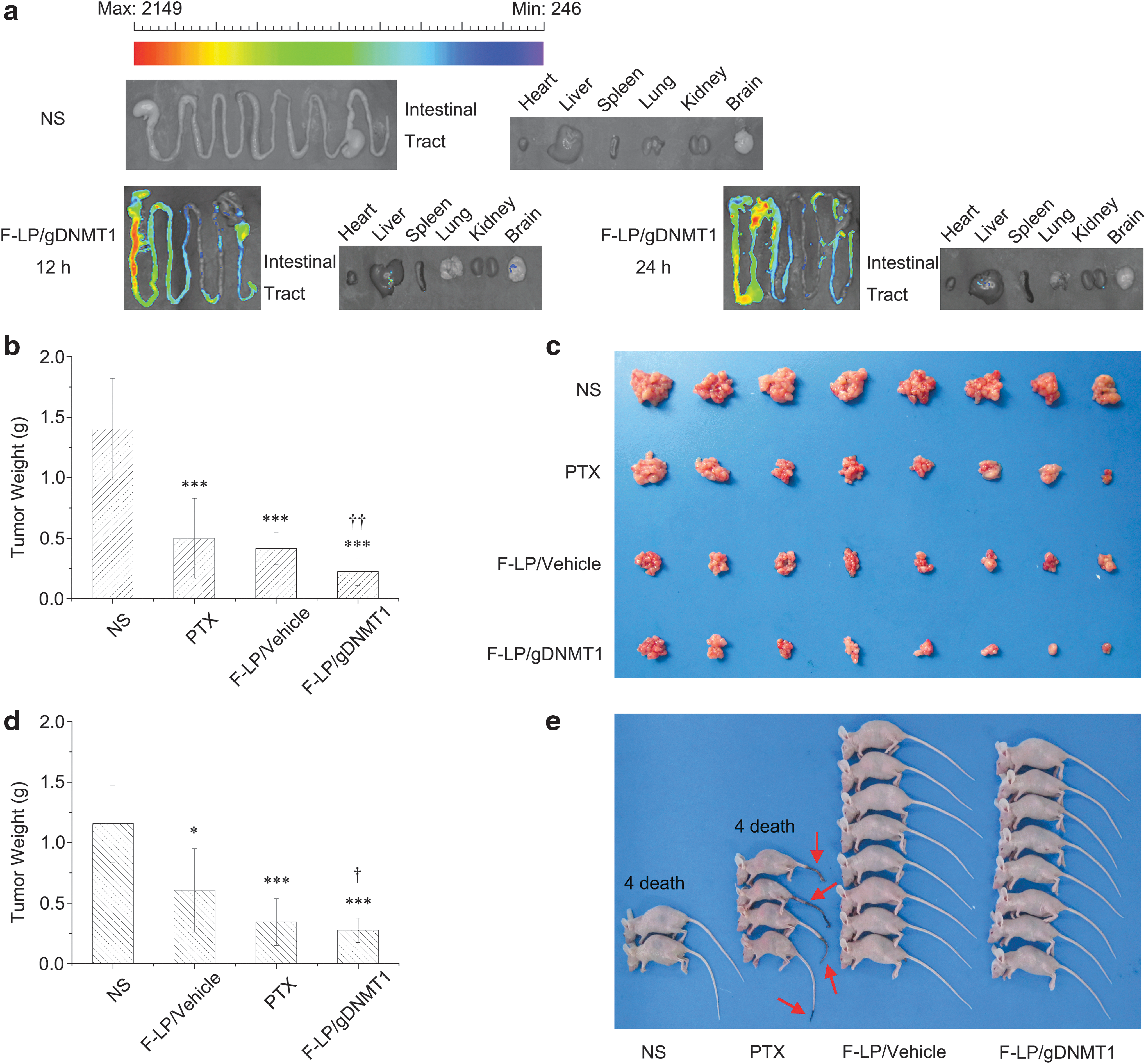
In vivo distribution and antitumor effects of F-LP/gDNMT1 complexes.
In vivo gene editing using F-LP/gDNMT1
The presence of CRISPR-Cas9 transcripts in tumor tissues treated with F-LP/vehicle or F-LP/gDNMT1 was determined by RT-PCR (Fig. 5a). Furthermore, F-LP/gDNMT1 downregulated DNMT1 and HP1α expression in tumor tissues (Fig. 5a). Because DNMT1 interacts with HP1 resulting in increased DNA methylation, 59 low DNMT1 and HP1α expression implies that the CRISPR-Cas9 system based on F-LP/gDNMT1 worked to decrease DNA methylation in ovarian cancer cells in vivo. Furthermore, some necrotic cells were observed in tumor tissues treated with F-LP/gDNMT1 (Fig. 5b and c), suggesting that the antitumor effect of F-LP/gDNMT1 works to decrease methylation and induce necrosis of tumor cells.

Reverse transcription (RT)-PCR, Western blot, and hematoxylin and eosin (H&E) staining of F-LP/gDNMT1-treated tumor tissues
In conclusion, F-LPs loaded with a CRISPR-Cas9 plasmid were successfully prepared and transfected into cancer cells in vitro and in vivo. These cationic liposomes were able to deliver the plasmid efficiently and produce targeted genome editing based on CRISPR-Cas9 technology. Regarding application of CRISPR-Cas9 technology for cancer gene therapy, the present study created ovarian cancer–targeted F-LP/gDNMT1 complexes that were stable for injection. F-LP/gDNMT1 introduced indels into the genome of ovarian cancer cells, inactivated endogenous DNMT1, and reduced genome methylation within ovarian cancer cells in vitro. Furthermore, F-LP/gDNMT1 downregulated expression of DNMT1 and HP1α and then decreased methylation of genomic DNA in ovarian cancer tissues in vivo. In addition, F-LP/gDNMT1 also induced necrosis of cancer cells in vivo. Consequently, F-LP/gDNMT1 effectively inhibited growth of PTX-sensitive and -resistant ovarian tumors in vivo and showed few adverse effects compared to high-dose PTX injection. However, more studies on the nonspecific antitumor effects of the control F-LP/vehicle complex need to be performed. Overall, cationic liposomal vectors were shown to be effective delivery systems for CRISPR-Cas9 technology, and F-LP/gDNMT1 was found to be a promising cancer therapeutic using CRISPR-Cas9 technology to produce precise genomic editing of cancer cells and tissues.
Footnotes
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (No. 81602699), the National High-Tech R&D Program of China (Nos. 2015AA020309 and 2014AA020708), the China Postdoctoral Science Foundation funded project (No. 2015M570791), and Salubris Academician Workstation for Innovative Biopharmaceuticals (No. 2017B090904017).
Author Disclosure
No competing financial interests exist.