
Abstract
Coxsackievirus B3 (CVB3), a single-stranded RNA virus of the picornavirus family, has been described as a novel oncolytic virus. However, the CVB3 strain used induced hepatitis and myocarditis in vivo. It was hypothesized that oncolytic activity and safety of CVB3 depends on the virus strain and its specific receptor tropism. Different laboratory strains of CVB3 (Nancy, 31-1-93, and H3), which use the coxsackievirus and adenovirus receptor (CAR), and the strain PD, which uses N- and 6-O-sulfated heparan sulfate (HS) for entry into the cells, were investigated for their potential to lyse tumor cells and for their safety profile. The investigations were carried out in colorectal carcinoma. In vitro investigations showed variable infection efficiency and lysis of colorectal carcinoma cell lines by the CVB3 strains. The most efficient strain was PD, which was the only one that could lyse all investigated colorectal carcinoma cell lines. Lytic activity of CAR-dependent CVB3 did not correlate with CAR expression on cells, whereas there was a clear correlation between lytic activity of PD and its ability to bind to HS at the cell surface of colorectal carcinoma cells. Intratumoral injection of Nancy, 31-1-93, or PD into subcutaneous colorectal DLD1 cell tumors in BALB/c nude mice resulted in strong inhibition of tumor growth. The effect was seen in the injected tumor, as well as in a non-injected, contralateral tumor. However, all animals treated with 31-1-93 and Nancy developed systemic infection and died or were moribund and sacrificed within 8 days post virus injection. In contrast, five of the six animals treated with PD showed no signs of a systemic viral infection, and PD was not detected in any organ. The data demonstrate the potential of PD as a new oncolytic virus and HS-binding of PD as a key feature of oncolytic activity and improved safety.
Introduction
Oncolytic virotherapy is a novel form of therapy for treatment of cancer, whose efficacy has been proven in several clinical trials in the last two decades. Significant achievements were followed by approval of talimogene laherparepvec as the first oncolytic virus (OV) for the treatment of melanoma in 2015 by the U.S. Food and Drug Administration (FDA) and in 2016 in Europe. 1 OVs preferentially replicate in cancer cells, spread within tumor tissue, and lead to tumor destruction. In addition, viruses also induce the innate and adaptive immune responses, resulting in immune-mediated tumor cell killing. 2
Various DNA viruses, such as adenovirus, parvovirus, vaccinia virus, and herpesvirus, and RNA viruses, such as enteroviruses, vesicular stomatitis virus, and reoviruses, have been used as OVs for the treatment of cancer. 3 –6 RNA viruses have a short replication cycle and produce a very large number of progeny, which gives them an advantage relative to DNA viruses in oncolytic virotherapy. Moreover, RNA viruses may be safer, as they lack the potential for genotoxic effects caused by integration into the host genome. 7 Over the course of the last 15 years, enteroviruses such as CVA21, echovirus 1, and poliovirus, which are single-stranded RNA viruses and belong to the Picornaviridae family, were evaluated for their potential as oncolytic agents against melanoma, breast cancer, and prostate cancer. 6,8 –10 Recently, another member of this group, coxsackievirus B3 (CVB3), was described as a novel OV for the treatment of lung carcinomas. CVB3-induced tumor destruction was a result of direct virus-induced cell lysis and virus-mediated stimulation of innate immunity, leading to the recruitment of cytolytic natural killer (NK) cells into the tumor. No treatment-related mortality was reported, but moderate hepatic dysfunction and mild myocarditis were reported to be the main side effects of CVB3 treatment of lung carcinoma in mice. 11
CVB3, like many other RNA viruses, has high mutation rates and can easily adapt to new cellular environments. 12 This important feature resulted in the emergence of several clinical and laboratory strains with varying infection and growth properties. A number of CVB3 strains have been characterized for their tissue tropism and organ toxicity in order to understand better the virus–host interaction and pathogenesis caused by viral infection. Among these CVB3 strains, there are strains that are highly cardiotropic, such as CVB3 H3, 31-1-93, M2, HA, or H310A1, 13 –16 whereas a number of other strains have been found to be low or non-cardiotropic (e.g., Nancy and PD). 16,17 There are also CVB3 strains that preferentially infect the liver. 18,19 Moreover, almost all known CVB3 strains are able to infect the pancreas. 20 One cause for the difference in pathogenicity is believed to be attributable to viral capsid proteins, which are directly involved in virus-cell attachment and virus uptake. In general, most CVB3 strains utilize the coxsackievirus and adenovirus receptor (CAR) as the primary receptor and the decay accelerating factor (DAF) as co-receptors to infect cells. 21,22 In contrast, the strain PD has a unique receptor tropism, as it uses N- and 6-O-sulfated heparan sulfates (HS) to enter the host cells. 23 However, PD also binds CAR and DAF, but the interaction with CAR is very weak. 15 With respect to carcinogenesis, the expression level of CAR and DAF are generally found to be high in cancer cells. However, the levels can be variable. 24 6-O-sulfated HS have an important role in progression of malignant tumors and are upregulated in many tumor types. 25
The aim of this study was to analyze the oncolytic efficiency and safety of the CVB3 strains Nancy, H3, 31-1-93, and PD in treating human colorectal cancer with respect to their ability to use different virus receptors for infection. It was found that all strains could infect colorectal carcinoma cell lines in vitro, but the highest efficiency was observed for PD. In vivo administration of Nancy, 31-1-93, and PD in a xenograft mouse model of colorectal carcinoma resulted in strong reduction of tumor growth. However, only PD showed a promising safety profile, whereas Nancy and 31-1-93 led to mortality of the animals.
Materials and Methods
Cell lines
HeLa cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Karlsruhe, Germany) supplemented with 5% fetal calf serum (FCS) and 1% penicillin-streptomycin. Colorectal carcinoma cell lines (DLD1, Colo205, Colo680h, Colo320, HCT116, and Caco-2) were grown in RPMI 1640 supplemented with 10% FCS, 1% penicillin-streptomycin, 1% L-Glutamine, and 1 mM of Na-Pyruvate (Invitrogen, Karlsruhe, Germany). LS174T cells were maintained in Eagle's minimum essential medium (EMEM; Lonza, Basel, Switzerland) and supplemented with 10% FCS, 1% penicillin-streptomycin, 1% L-Glutamine, and 1 mM of Na-Pyruvate (Invitrogen). SW480 and SW620 cell lines were cultured in Leibovitz L15 (Biowest, Darmstadt, Germany) supplemented with 10% FCS, 1% penicillin-streptomycin, and 1% L-Glutamine.
Viruses
The CVB3 strain PD was derived by serial passages of the CVB3 strain P on HuFi cells. 26 The cardiotropic CVB3 strain 31-1-93 was isolated from heart tissue after four heart passages of PD in outbred NMRI mice. 14 Both virus strains PD and 31-1-93 were a kind gift from Michaela Schmidtke (Institute of Virology and Antiviral Therapy, University Jena, Germany). CVB3 PD and 31-1-93 were propagated in CAR/DAF-negative CHO-K1 and in HeLa cells, respectively, before use. CVB3 strain Nancy (ATCC VR30) was propagated in HeLa cells. CVB3 H3 was generated by transfection of the cDNA containing plasmid pBK-CMV-H3 (kindly supplied by Andreas Henke, Institute of Virology and Antiviral Therapy, University of Jena, Jena, Germany) into HEK293T cells using Polyethylenimine Max (Polysciences, Inc., Warrington, PA). Completely lysed cells were harvested 48 h post transfection, and the virus was stored in aliquots after three freeze/thaw cycles and removal of the cell debris by centrifugation.
Viral plaque assay
Viral plaque assays were carried out, as previously described. 27 Briefly, HeLa cells were cultured in 24-well cell culture plates as confluent monolayers. After 24 h, the medium was removed, and cells were overlaid with serial 10-fold dilutions of supernatant harvested from virus-infected cell lines or from homogenized mouse organs after three freeze/thaw cycles. Cells were then incubated at 37°C for 30 min and, after removal of the supernatant, overlaid with agar containing EMEM. Three days later, the cells were stained with 0.025% neutral red in phosphate-buffered saline (PBS). Virus titers were determined by plaque counting 3 h after staining.
Cell killing assay
Colorectal carcinoma cells were seeded in 96-well plates. On the following day, when cells reached complete confluence, the medium was carefully removed, and virus solution (100 μL) was added at a multiplicity of infection (MOI) of 1, 10, and 100. After 30 min of incubation at 37°C and 5% CO2, virus-containing medium was removed, fresh medium was added, and cells were incubated for different periods. To fix the cells, the medium was removed, the cells were washed with PBS, and 10% trichloroacetic acid (TCA; Carl Roth, Karlsruhe, Germany) was added. Following incubation for 10 min, TCA was removed, and 30 μL of crystal violet solution (Carl Roth) was added. After 5 min of incubation, wells were washed with PBS several times, and the plate was allowed to dry overnight before being photographed.
Heparin inhibition assay
Colorectal carcinoma cells were seeded in 96-well plates and grown until they reached full confluence. Twenty-four hours later, HS analogue heparin (porcine mucosal intestinal; Sigma–Aldrich, St. Louis, MO) was dissolved in serum-free DMEM to reach a stock concentration of 10 mg/mL and stored at 4°C. Heparin was diluted to different concentrations in serum-free DMEM, and each dilution incubated with MOI 3 of CVB3-Nancy or PD for 1 h at 37°C. Heparin-free DMEM was used as control. Afterwards, the media was removed from the plates, and the cells were infected with HS-treated virus solutions and incubated for 30 min at 37°C. The solution was removed, and 100 μL of fresh cell culture medium was added. Plates were incubated for 48 h and used for plaque assay or XTT assay.
HS6ST2 quantification
To investigate expression levels of HS6ST2 in colorectal cell lines, quantitative reverse transcription polymerase chain reaction (RT-PCR) was performed. Total RNA was extracted by using TRIzol reagent (Life Technologies GmbH, Darmstadt, Germany) according to the manufacturer's instructions. Isolated RNA (1 μg) was reverse transcribed into cDNA using a High Capacity cDNA Reverse Transcription Kit (Life Technologies GmbH). Real-time PCR was performed using the HS6ST2 forward primer, 5′-CCAAGTCAAATCTGAAGCACA-3′ and the reverse primer 5′-TCTGGAAATGGGTCTGAAGGA-3′ in Sso Fast™ EvaGreen® Supermix (Bio-Rad, Hercules, CA). Cycle times were as follows: one cycle at 95°C for 30 s followed by 40 cycles alternating between 95°C for 5 s and 60°C for 5 s. The results were normalized to that of the 18S rRNA gene. The HS6ST2/18S mRNA ratio was calculated for each colorectal cell line to measure the relative mRNA expression. The PCRs were carried out in triplicate, and the relative HS6ST2 expression was determined by the ΔΔCt calculation method.
CVB3 RNA quantification
Colorectal carcinoma cells were seeded in 96 well plates and infected with CVB3 strains. Twenty-four hours later, plates were subjected to three freeze/thaw cycles, and the collected supernatant was centrifuged for 20 min at 2,000 g and 4°C to remove cellular debris. Viral RNA was isolated from the supernatant with a High Pure Viral Nucleic Acid Kit (Roche, Mannheim, Germany) according to the manufacturer's instructions, followed by DNase I digestion (Peqlab, Erlangen, Germany). The viral RNA was reverse transcribed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). For quantification of viral RNA, real-time PCR was performed using the forward primer, 5′-CCCTGAATGCGGCTAATCC and the reverse primer 5′-ATTGTCACCATAAGCAGCCA in Sso Fast™ EvaGreen® Supermix (Bio-Rad). Cycle times were as follows: one cycle at 50°C for 2 min followed by 94°C for 10 min, 40 cycles at 94°C for 15 s, and 60°C for 60 s. A standard curve was used to calculate the number of CVB3 genome copies. The standard curve was prepared using 10-fold serial dilutions of 146 bp PCR fragment of CVB3 plasmid, which was amplified with the forward and reverse primers mentioned above. The PCR reactions were carried out in triplicate, and the relative CVB3 RNA copies were determined by the ΔΔCt calculation method.
Flow cytometric analysis of CAR and DAF expression
Colorectal carcinoma cells were washed with PBS and detached from cell culture plates using PBS–2 mM EDTA solution. After washing with PBS, the cells were stained on ice with monoclonal mouse anti-CAR/IgG1 (clone RmcB) antibody (Merck KGaA, Darmstadt, Germany) or with monoclonal mouse anti-DAF/CD55 (Merck) for 1 h at a dilution of 1:200. Cells were washed again with PBS and re-suspended in Alexa Fluor 488-conjugated donkey anti-mouse IgG antibody (Life Technologies GmbH), which was diluted 1:400 in PBS, and incubated for 45 min. After a further wash step with PBS, the cells were re-suspended in PBS +1% formaldehyde and analyzed by flow cytometry using a MACSQuant® Flow Cytometer (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) and FlowJo Data Analysis Software (Tree Star, San Carlos, CA). The mean fluorescence intensity was calculated by determining the geometric mean of CAR- or DAF-expressing cells minus the geometric mean of a negative control sample.
In situ hybridization
Probes for the detection of CVB3 RNA by in situ hybridization were generated using the Dig RNA labeling Kit (Roche) using the pCVB3-R1 plasmid, which was linearized with SmaI, as previously described. 28 Pretreatment, hybridization, and washing steps were done, as described previously. 29 Detection of the DIG-labeled RNA-probe was performed with a horseradish peroxidase-conjugated anti-Digoxigenin antibody (Roche; 1:100) and HistoGreen (Linaris GmbH, Dossenheim, Germany) as substrate. Sections were counterstained with hematoxylin and mounted with Pertex mounting media (Medite, Burgdorf, Germany).
In vivo experiments
Animal experiments were performed in accordance with the principles of laboratory animal care and all German laws regarding animal protection. DLD1 cells (5 × 106 cells) were inoculated subcutaneously into the right and left flanks of 6-week-old BALB/c nude mice. Tumor burdens were measured daily by hand caliper, and CVB3 strains were injected intratumorally at a dose of 3 × 106 plaque forming units (pfu) per animal into one of the tumors when the tumor size reached 0.4–0.5 cm.
Histopathological analysis
The mouse tissues and explanted human tumors were fixed in 4% paraformaldehyde, embedded in paraffin, and 5 μm thick tissue sections were cut and stained with hematoxylin and eosin to visualize and quantify cell destruction and inflammation.
Statistical analysis
Statistical analysis was performed with GraphPad Prism (GraphPad Software, Inc., La Jolla, CA). Results are shown as the mean ± standard error of the mean for each group. Statistical significance was determined by use of the two-tailed unpaired Student t-test for cell culture investigations and by use of the one-tailed Mann–Whitney U-test for in vivo investigations. Differences were considered significant at p < 0.05. Survival curves were plotted according to the Kaplan–Meier method (log-rank-test).
Results
CAR and DAF are differentially expressed on colorectal carcinoma cell lines
The CVB3 strains Nancy, 31-1-93, and H3 use CAR as their primary receptor and DAF as their co-receptor for infection, whereas the strain PD uses HS but can also bind to CAR and DAF. 15,30 To determine the expression of the CVB3 receptors on colorectal carcinoma cells, the level of CAR and DAF of nine colorectal carcinoma cell lines (Colo680h, Colo205, DLD1, Colo320, Caco-2, HCT116, SW480, SW620, and LS174T) was measured using flow cytometry. Seven cell lines expressed moderate levels of CAR, while two cell lines (Colo680h and Colo205) expressed CAR only at a low level. Four cell lines showed high (Colo680h, DLD1, Caco2, and HCT116) and moderate (Colo320, SW480, SW620, and LS174T) levels of DAF expression, whereas one cell line (Colo205) almost completely lacked DAF on the cell surface (Fig. 1). These data demonstrate that CAR and DAF are expressed at variable levels on the cell surface of colorectal carcinoma cell lines, with the majority of cell lines expressing CAR and DAF at high or moderate levels.

Expression levels of coxsackievirus and adenovirus receptor (CAR) and decay accelerating factor (DAF) on the surface of colorectal carcinoma cell lines and on HeLa cell line. Upper panel: Cell surface expression of CAR and DAF (dark gray graphs) was measured by flow cytometry using anti-CAR (RMCB) and anti-DAF (CD55) antibodies, respectively. As negative control (light gray graph), cells were stained only with the secondary antibody (Alexa Fluor® 488). HeLa cells were used as positive control due to high level expression of CAR and DAF. The black highlighted area represents the set gate. The percentages indicate the proportion of CAR- or DAF-positive cells. Lower panel: Diagram of cell surface expression of CAR and DAF in colorectal carcinoma cells. Columns shown means ± standard error of the mean (SEM) of three independent experiments.
CVB3 strains differentially infect and lyse colorectal carcinoma cell lines
To determine whether CAR/DAF expression may influence the ability of CVB3 Nancy, 31-1-93, H3, and PD to infect and lyse colorectal carcinoma cell lines, five human colorectal carcinoma cell lines (DLD1, Colo680h, Colo205, Colo320 and LS174T) exhibiting different levels of CAR and DAF expression were used. These cell lines were infected with a virus strain at a MOI of 1 or 10, and viral infection was determined by measurement of the amount of viral RNA genomes 24 h after infection by quantitative RT-PCR. Each virus strain was detected in its target cell line, but there were significant differences in the infection rates. High and moderate infection rates were detected for PD in all cell lines. Strain 31-1-93 also showed moderate and high infection rates, but only in some of the colorectal carcinoma cell lines (LS174T and DLD1). In the other cell lines, it was low. The infection rates for Nancy and H3 were generally low (Supplementary Fig. S1; Supplementary Data are available online at
To determine cytolytic activity of the CVB3 strains, colorectal carcinoma cell lines were infected with the viruses at a MOI of 1, 10, or 100, and analyzed after 24, 48, and 72 h for cytotoxicity using crystal violet staining. The cytolytic efficiency of the CVB3 strains was highly variable. Much like the infection data, PD showed strong cytolytic activity in DLD1, Colo680h, and Colo205 and moderate activity in Colo320 and LS174T. The strain 31-1-93 induced cell lysis in four of the five investigated cell lines, but high cytolytic activity was only detected in the cell line DLD1. Nancy and H3 had comparatively low cytolytic activity. Cytolysis was only observed at high MOIs in DLD1 and Colo320 cells (Fig. 2 and Table 1). There was no clear correlation between CAR/DAF expression levels and infection/lysis of colorectal carcinoma cell lines by the CVB3 strains. These data demonstrate that the CVB3 strain PD most efficiently infected and lysed each of the colorectal carcinoma cell lines.

CVB3 strains have different oncolytic efficacy against colorectal carcinoma cells in vitro. The cell killing assay was performed with CVB3 Nancy, H3, PD, and 31-1-93 strains. HeLa cells and DLD1, Colo680h, Colo205, Colo320, and LS174T colorectal carcinoma cells were infected with 1, 10, or 100 multiplicities of infection (MOI) of virus. Cell viability was determined 24
Cytolytic efficiency of PD in colorectal carcinoma cells is determined by binding to heparan sulfate
As distinct from other strains, PD can use N- and 6-O-sulfated HS to attach and infect cells. To ascertain the importance of the interaction of PD with HS for infection and lysis of colorectal carcinoma cell lines, the expression level of HS6ST2 was analyzed by quantitative RT-PCR. HS6ST2 catalyzes the transfer of sulfate groups to position 6 of the N-sulfoglucosamine residue in HS and leads to production of 6-O-sulfated HS. 31 The investigations were carried out with DLD1, Colo205, and Colo680h cell lines, which are susceptible to PD, and with the LS174T and Colo320 cell lines, which are resistant to PD (Table 1 and Fig. 2). As shown in Fig. 3A, HS6ST2 mRNA expression was detected in DLD1, Colo205, and Colo680h cells, but not in LS174T and Colo320 cells, indicating a correlation between HS6ST2 expression and susceptibility of colorectal carcinoma cell lines for PD.
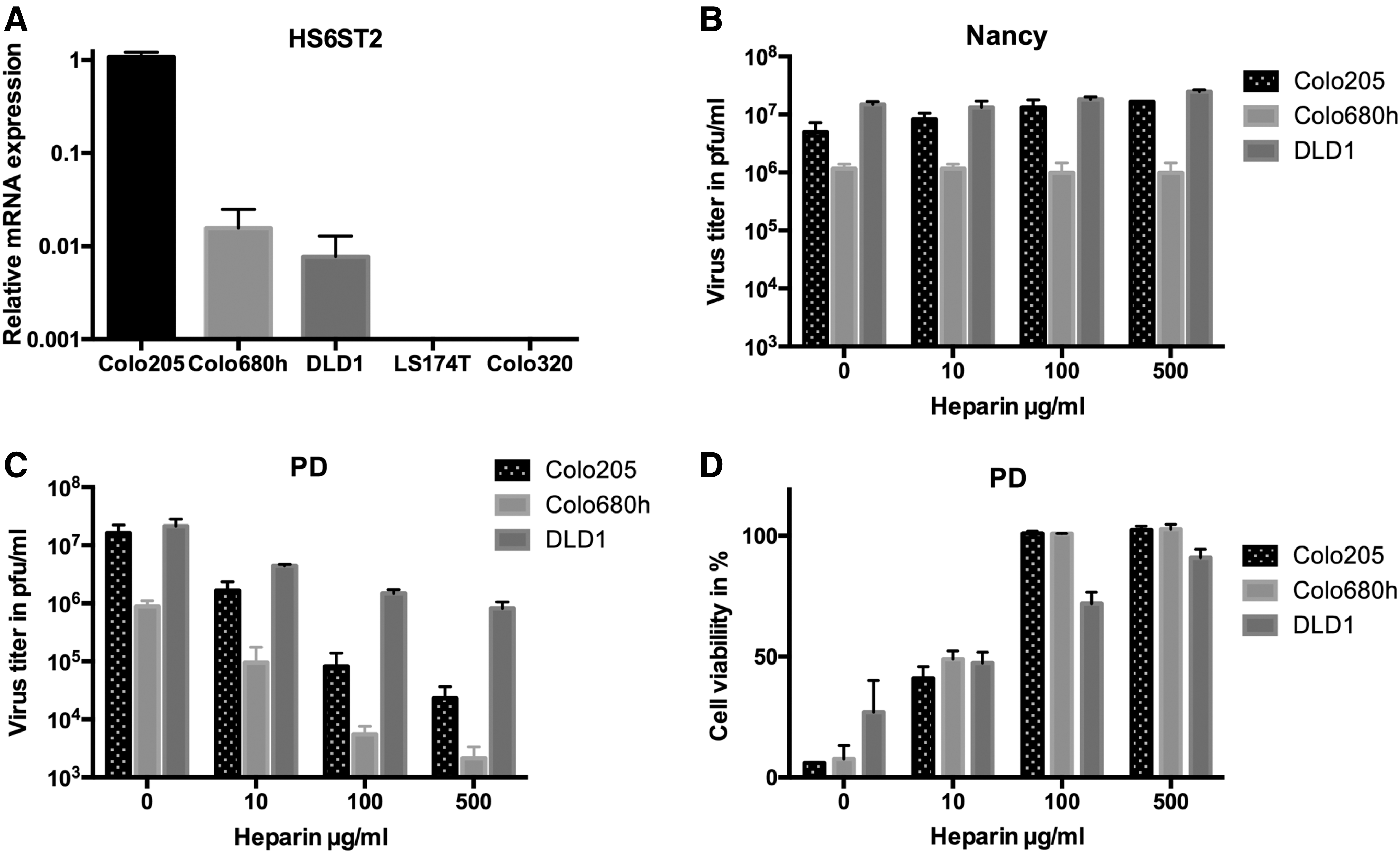
Heparan sulfate mediates infection of PD.
CAR and DAF expression and lytic activity of CVB3 strains in different colorectal carcinoma cell lines
Sensitivity was indicated relatively by the number of +. +++++, complete lysis (100%); ++++, 80% lysed, etc. A free field means resistance of the cell line to the virus variant.
CAR, coxsackievirus and adenovirus receptor; DAF, decay accelerating factor.
To confirm that PD indeed uses HS as receptor to infect colorectal carcinoma cells, the soluble HS-analogue heparin was used. CVB3 Nancy was used as control. PD and Nancy were incubated with different concentrations of heparin before infection and virus infection was analyzed by plaque assay 48 h later. While infection with Nancy was not affected by heparin treatment, and the virus titers were similar at each heparin concentration (Fig. 3B), PD infection was distinctly inhibited by heparin treatment in a dose-dependent manner. The effect was more prominent in Colo205 and Colo680h cell lines than in the cell line DLD1 (Fig. 3C). The HS blocking experiments were repeated, and the cytotoxic activity measured by XTT assay. PD-induced cell lysis was completely inhibited by 100 μg/mL of heparin in Colo205 and Colo680h, whereas inhibition reached about 80% in the DLD1 cell line (Fig. 3D). The lower heparin-induced inhibition of PD infection and cytotoxicity in DLD1 cells compared to Colo205 and Colo680h cells may be due to high levels of CAR on the surface of these cells, as PD can also use CAR for infection. 15
In conclusion, these data demonstrate that PD uses HS to infect colorectal cancer cells, and the interaction with HS is a key factor for the cytolytic activity of PD.
CVB3 strains have strong oncolytic activity but different toxicity in vivo
To evaluate oncolytic efficiency of CVB3 strains in vivo, a xenograft BALB/c mouse tumor model was established with the DLD1 human colorectal carcinoma cell line. Tumor cells were inoculated bilaterally into the flanks of the animals, and only one of the two tumors was injected with single dose of 3 × 106 pfu of CVB3 Nancy, 31-1-93, or PD. Treatment with any of the three viruses resulted in significant suppression of tumor growth of both the virus-injected tumor and the contralateral untreated tumor when compared with uninfected mice tumors (p < 0.05; Fig. 4A–C).
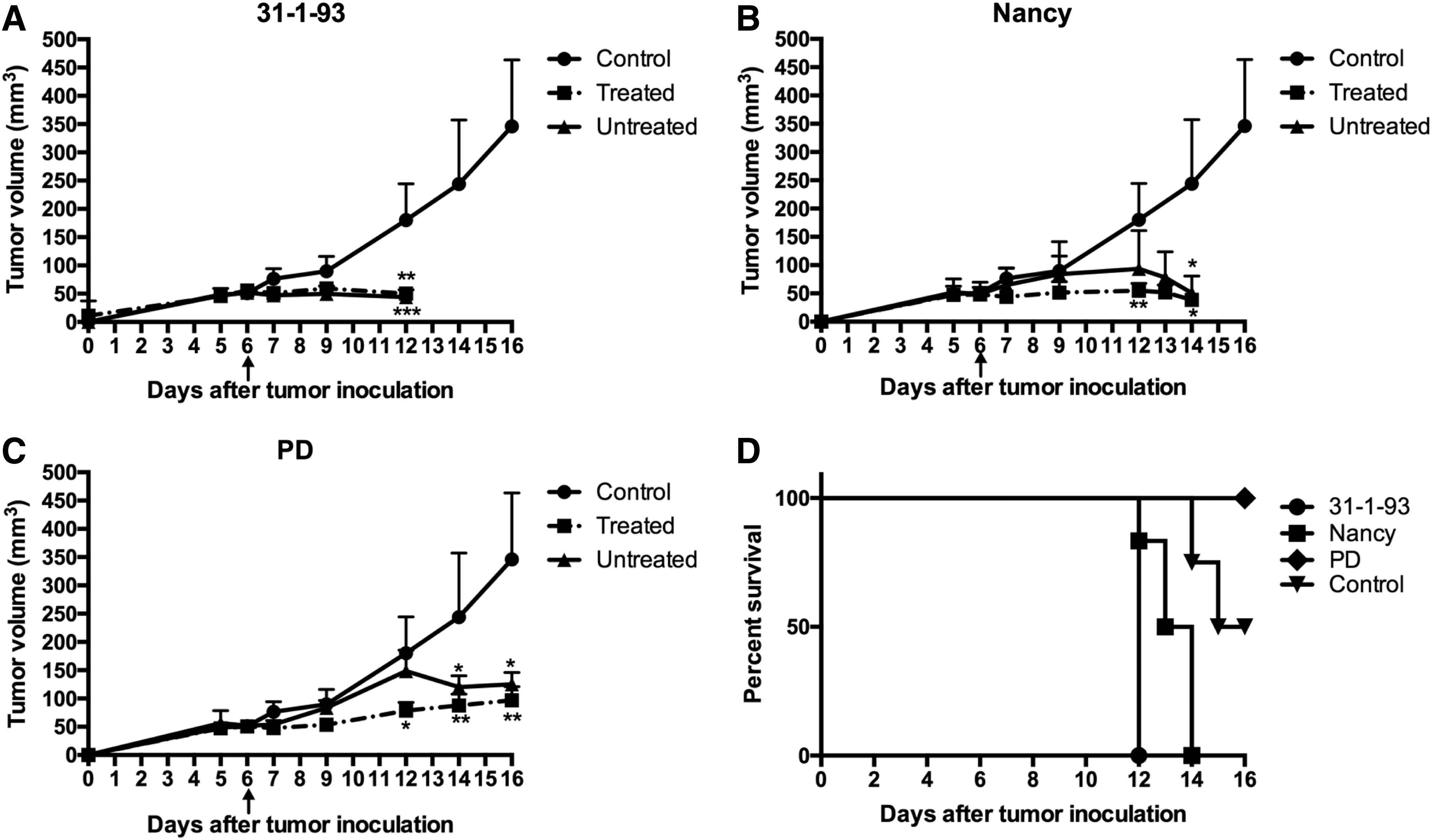
In vivo oncolytic efficiency of CVB3 strains in a DLD1 xenograft mouse model. DLD1 cells were subcutaneously inoculated into the right and left flank of BALB/c nude mice. One of the inoculated tumors was intratumorally injected with a single dose (3 × 106 plaque forming units) of the CVB3 strains
Severe side effects were observed for CVB3 Nancy- and 31-1-93-infected animals. All animals that were injected with 31-1-93 died 6 days after intratumoral virus injection, and animals treated with Nancy were moribund at 6 (one animal), 7 (one animal), and 8 days (four animals) after virus administration and were sacrificed. Animals infected with 31-1-93 were not examined further, as these mice showed signs of autolysis on day 6. In contrast to Nancy- and 31-1-93-infected animals, PD-infected animals showed a significantly prolonged survival (p = 0.0006; Fig. 4D). All six animals treated with PD were alive 10 days after intratumoral virus injection, and five of them showed a normal physical state, similar to the untreated negative control group. One of the PD-infected animals (PD-M1) showed slower movement and lower body weight compared to the other PD-infected animals.
Measurement of the amount of infectious virus in the tumors of Nancy- and PD-infected animals by plaque assay 8 and 10 days after virus injection, respectively, showed that both viruses were detected at similar levels (between 3.2 and 7.2 × 106 pfu/g) in the primary injected tumor. Both viruses were also detected in the contralateral, untreated tumor, but the amount was significantly lower in the PD-injected group than in Nancy-treated animals (p < 0.05; Fig. 5A). Moreover, in two of six investigated animals, PD was not detected in the untreated tumor. In situ hybridization confirmed the presence of viral genomic RNA in the tumors of Nancy- and PD-infected animals, as well as the absence of virus in untreated tumors of two PD-infected animals, which were also negative when measured for replicating virus. The investigations also showed that viruses were located in mononuclear immune cells, probably macrophages, within the tumor mass or on the border between the tumor stroma and tumor cells (Fig. 5B).
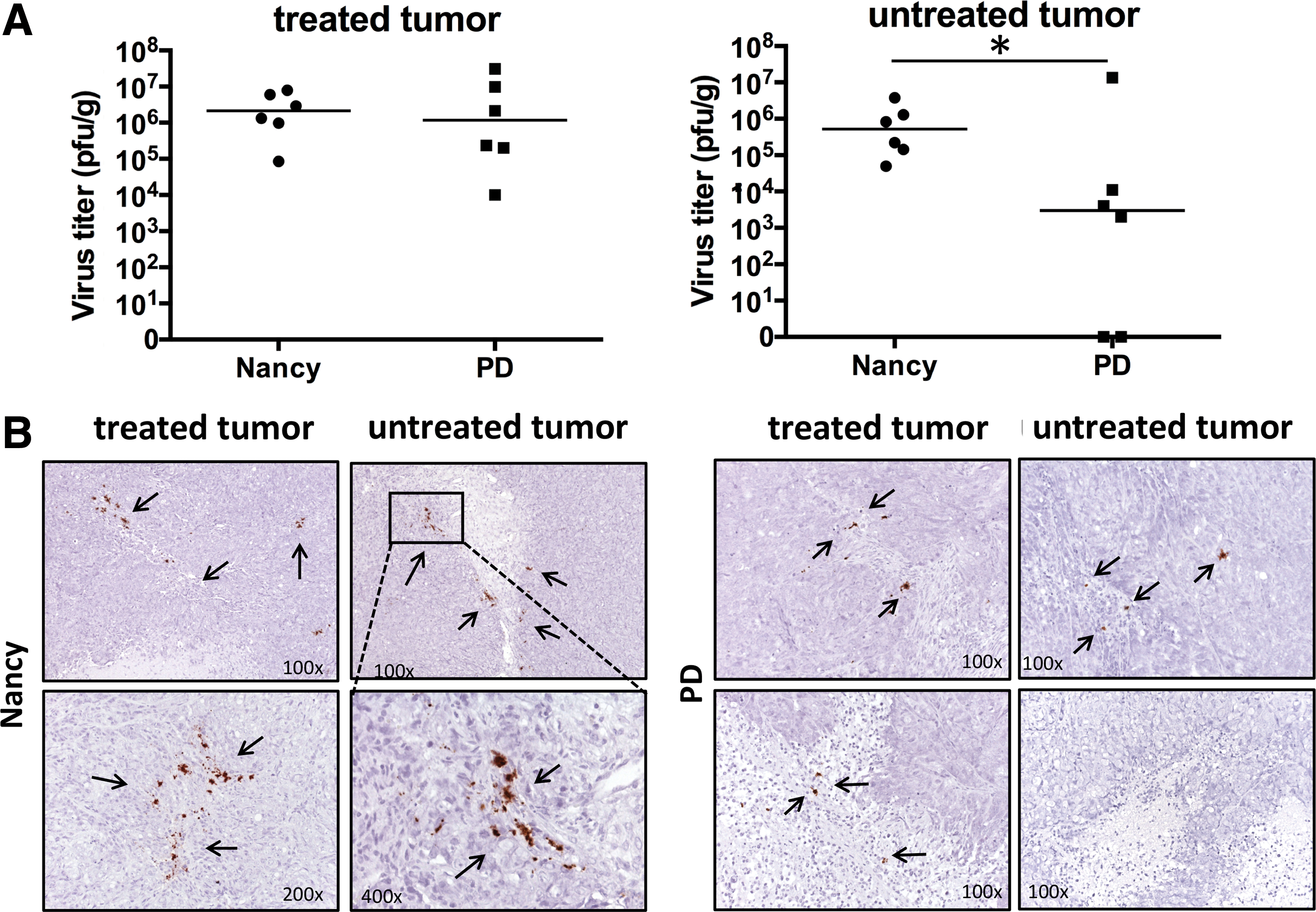
Infectivity and replication activity of CVB3 Nancy and PD strain in harvested tumors.
Taken together, these results demonstrate that CVB3 Nancy, 31-1-93, and PD have strong oncolytic activity in vivo, but only PD had a sufficient safety profile.
CVB3 Nancy but not PD infect normal organs
To elucidate the cause of the severe side effects, the occurrence of Nancy and PD infection in the heart, liver, brain, spleen, kidney and pancreas was investigated. Using plaque assays to detect replicating virus, the highest virus titers (of about 106 pfu/g) were measured in the heart of the Nancy-treated group (Fig. 6A). CVB3 Nancy was also detected in the spleen, kidney, and brain, but at distinctly lower levels, whereas it could only sporadically be detected in the liver (one of six animals) at a very low titer (102 pfu/g). The pancreas is the most susceptible organ for CVB3 in mice. 32 Examination of the pancreas tissue in Nancy-infected animals during dissection showed reduced organ size (not shown), indicating major damage to the tissue. As the destruction of the pancreas by CVB3 made the tissue unsuitable for determination of infectious virus by plaque assay, real-time RT-PCR was carried out to quantify the virus genomes. High amounts (103–105 copies/μg RNA) of Nancy viral RNA was detected in the pancreas (Fig. 6A). In sharp contrast, no infectious virus was recovered from any organ of five of the six PD-treated animals. Only the animal PD-M1 contained a similar amount of virus as the animals in the Nancy group. To elucidate the cause of observed PD toxicity in PD-M1, the virus was isolated from the animal's heart tissue, and the genomic region encoding the viral proteins VP1, VP2, VP3, and VP4 was sequenced. Sequencing revealed the occurrence of two amino acid substitutions in VP1 (V108I and V245I) and one amino acid substitution in the VP2 region (K230E) relative to the sequence of the starting strain of PD, indicating that the virus (hereafter referred to as CVB3-PD-M1V) isolated from PD-M1 was a mutated variant of PD.
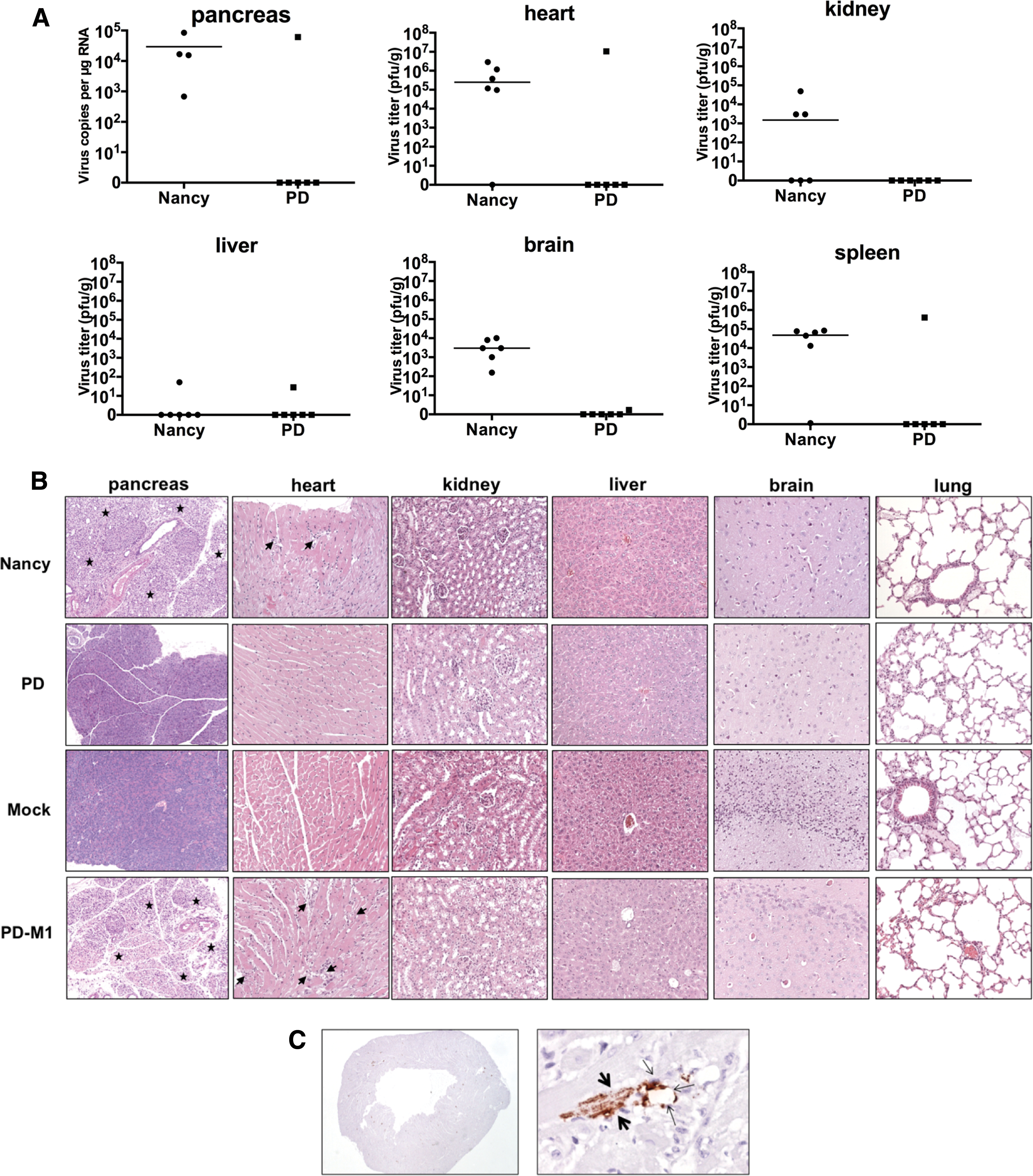
Viral load and pathological alterations of CVB3 Nancy and PD in organs of tumor-bearing mice. Organs of the animals (n = 6 for each group) were harvested at the time points when animals were sacrificed, as described under Fig. 4.
Histological examination of mouse tissues (the pancreas, heart, kidney, liver, brain, and lung) confirmed different toxic activity of Nancy and PD. The exocrine pancreas of Nancy-infected animals was nearly completely destroyed, and heart tissue of these animals showed distinct inflammation. All other organs of Nancy-infected animals were unaffected. In contrast, with the exception of the one animal, PD-M1, all organs of PD-treated animals did not show any alterations (Fig. 6B). The animal PD-M1 showed organ alterations, which were very similar to those of Nancy-infected animals. Furthermore, in situ hybridization of the heart of PD-M1 demonstrated that the virus CVB3-PD-M1 infected cardiac myocytes and cardiac endothelial cells (Fig. 6C).
Discussion
Enteroviruses represent one group of viruses containing members with oncolytic activity. Major advantages of the enteroviruses are their fast replication cycle, the production of high amounts of virus progeny, the small virus size, which enables better tumor penetration, and their ability to stimulate a powerful antitumor immune response. 3,7 CVA21 and PVS-RIPO are the best investigated oncolytic enteroviruses. Both are currently under clinical investigation for the treatment of cancer. 9 More recently, Miyamoto et al. 11 showed that another enterovirus, CVB3 (strain Nancy), has oncolytic activity and was highly effective at destroying a lung carcinoma in vivo. However, the preclinical study by Miyamoto et al. also revealed that the Nancy strain induced hepatic and cardiac injury in tumor-bearing mice.
There are several clinical and laboratory strains of CVB3, which are characterized by different tissue tropism, pathogenicity, and course of infections. 15 Among them, PD was shown to be a non-virulent strain due to its unique tropism to N- and 6-O-sulfated HS. 16 Thus, it was hypothesized that CVB3 PD might have an oncolytic activity with an improved safety profile compared to other CVB3 strains. Accordingly, this study compared a panel of different CVB3 strains for their antitumor efficiency and safety in vitro and in vivo using colorectal carcinoma cells as the target. The study shows that the CVB3 strain PD is a potent and efficient OV against colorectal carcinomas in vitro and in vivo, and more importantly, PD does not regularly induce side effects in vivo. No systemic infection with PD was detected, and the virus could not be recovered from the organs of treated animals. In contrast, two other analyzed CVB3 strains, Nancy and 31-1-93, although showing similar antitumor efficiency as PD in vivo, induced severe systemic infection in mice, leading to the death of the animals.
Expression of virus receptors on the cell surface is an important prerequisite for virus infection and one of the major determinants of tumor selectivity for oncolytic RNA viruses. CVB3 Nancy, 31-1-93, and H3 utilize CAR and DAF for infection. 21,22 It was found that the majority of colorectal carcinoma cells lines express moderate to high levels of CAR and DAF, but, with exception of the DLD1 cell line, they were more or less resistant against both strains. It may be possible that the receptor expression was not high enough to mediate efficient virus infection, but lack of a clear correlation between CAR/DAF expression and virus-induced cell lysis, as well as individual differences between Nancy, H3, and 31-1-93, suggests that additional, receptor-independent mechanisms may at least in part be responsible for this result. The CVB3 strain PD lysed colorectal carcinoma cell lines more efficiently than the strains Nancy, H3, and 31-1-93. A major difference of PD relative to Nancy, H3, and 31-1-93 is its receptor tropism. PD uses N- and 6-O-sulfated HS for infection of cells. 30 HS are linear polysaccharides consisting of repeating disaccharides, which are linked to the cell surface by core proteins. Although nearly ubiquitously expressed, HS vary in sulfate content and charge among different tissues and cells. 23,33 In cancer, HS play a complex role in tumor progression, metastasis, and invasion. 34 N- and 6-O-sulfated HS are highly abundant in many cancer cells, including colorectal carcinoma cells. 31 The latter was confirmed in this study by detection of HS6TS2, which is involved in synthesis of N- and 6-O-sulfated HS. Furthermore, the results reveal the importance of HS for the oncolytic activity of PD in colorectal carcinoma cells. In this regard, a clear correlation was found between cellular HS6TS2 expression levels and sensitivity of colorectal carcinoma cell lines to PD, as well as demonstrated that blocking the binding of PD to HS strongly inhibits lytic activity of the virus.
In contrast to recently described oncolytic CVB311 and other CVB3 strains investigated in this study, PD was completely attenuated in vivo. This is in accordance with other studies showing the HS-binding viruses are in general attenuated in vivo. 35 Even though the mechanism is not fully understood, attenuation seems to be caused by a reduced ability of the virus to spread from the site of inoculation via the blood stream 36 and by a more rapid clearing from the circulation. 35 However, in the case of PD, the specific tropism of PD to N- and 6-O-sulfated HS may provide an additional explanation. The pancreas, heart, spleen, and brain are major target organs of CVB3 strains using CAR and DAF for infection. However, these organs do not express HS6ST2. 31 Thus, resistance of these organs against PD as found in this study may be directly related to the lack of the specific PD receptor.
It was found that intratumoral injection of tumor-bearing nude mice with PD, but also Nancy and 31-1-93, led to significant inhibition of growth of the injected tumor. Even more importantly, similar growth suppression was also observed for the distant, non-injected subcutaneous tumor. Whereas these results demonstrate per se the efficacy of CVB3-mediated oncovirotherapy in colorectal cancer, suppression of the untreated tumor reveals the potential of PD to treat metastatic or disseminated colorectal cancer. It is important to mention that the DLD1 cell line, which was used to establish subcutaneous tumors, is highly resistant to radiation 37 and against 5-fluorouracil, which is used as chemotherapeutic agent for the treatment of colorectal cancer. 38 PD may therefore be an alternative treatment modality for colorectal cancers that are resistant to conventional therapies. Interestingly, CVB3 Nancy and PD RNA was detected in immune cells within the tumor masses on the tumor-stroma border zone, whereas tumor cells seemed not to be infected. The failure of virus detection in tumor cells was a surprising result, as virus-induced cell lysis is thought to be a major mechanism of antitumor efficiency of OVs. 39 The reasons for this are currently not clear. However, the detection of CVB3 genomes in immune cells and their specific localization in the tumors suggest that virus-induced immune-mediated mechanisms may be involved in antitumor efficiency of the CVB3 strains. This conclusion is supported by a previous study demonstrating that CVB3 replication in lung carcinoma results in recruitment of NK cells and granulocytes into the tumor tissue. 11 Furthermore, the data indicate that immune cells may be carriers and probably a reservoir for hematogenous distribution of the viruses within a tumor or to distant tumors.
One (animal PD-M1) of the six PD-treated animals became ill and showed high levels of viral progeny in its heart, spleen, and pancreas. Sequence analysis revealed two amino acid substitutions in VP2 and one amino acid substitution in VP1 in the virus compared to PD, indicating that the isolated virus, CVB3-PD-M1V, was a mutant that originated from PD. Even though further functional investigation was not carried out, the detected mutation may be responsible for different phenotypes of PD-M1V compared to PD. In fact, the amino acid substitutions found in VP2 of PD-M1 are linked to be specific for some CVB3 strains inducing myocarditis. 13,40 Two mechanisms, which are closely related to the life cycle of CVB3, the high mutation rate and the quasispecies dynamics, 41 may be responsible for the occurrence of CVB3-PD-M1V. There may be different ways to overcome development of undesirable PD mutants. For example, the use of cDNA clones 13 may reduce the strong heterogenicity of quasispecies and reduce the risk of the emergence of mutants. MicroRNA-mediated virus de-targeting strategies, which have successfully been used for several OVs, 42 –46 may represent another way to prevent toxicity.
In summary, the present findings demonstrate oncolytic efficiency of different CVB3 strains in colorectal carcinoma, but only the strain PD has a favorable safety profile. Due to its specific receptor tropism to HS, PD can infect and lyse tumor cells more efficiently than the CAR-binding CVB3. Therefore, PD may extend the spectrum of anticancer agents.
Footnotes
Acknowledgments
This work has been supported by the TU Berlin through ASF:312Y project grant and by the Wilhelm Sander-Stiftung through 2017.101.1 project grant to H.F. and J.K., by Berliner Krebsgesellschaft through Ernst von Leyden Stipendium to A.H., and by The Scientific and Technological Research Council of Turkey (TUBITAK) to A.H. We thank Michaela Schmidtke (Department of Virology and Antiviral Therapy, Jena University Hospital Jena, Germany) for kindly providing the CVB3 strains PD and 31-1-93, and Andreas Henke (Department of Virology and Antiviral Therapy, Jena University Hospital Jena, Germany) for the supply of CVB3 strain H3 cDNA encoding plasmid. We thank Erik Wade for critical reading of the manuscript and helpful comments.
Author Disclosure
No competing financial interests exist. H.F., A.H., and V.B. have a patent pending for use of CVB3 strain PD in treatment of cancer.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.