
Abstract
Umbilical cord blood (UCB) has been increasingly explored as an alternative source of stem cells for use in regenerative medicine due to several advantages over other stem-cell sources, including the need for less stringent human leukocyte antigen matching. Combined with an osteoinductive signal, UCB-derived mesenchymal stem cells (MSCs) could revolutionize the treatment of challenging bone defects. This study aimed to develop an ex vivo regional gene-therapy strategy using BMP-2-transduced allogeneic UCB-MSCs to promote bone repair. To this end, human UCB-MSCs were transduced with a lentiviral vector carrying the cDNA for BMP-2 (LV-BMP-2). In vitro assays to determine the UCB-MSC osteogenic potential and BMP-2 production were followed by in vivo implantation of LV-BMP-2-transduced UCB-MSCs in a mouse hind-limb muscle pouch. Non-transduced and LV-GFP-transduced UCB-MSCs were used as controls. Transduction with LV-BMP-2 was associated with abundant BMP-2 production and induction of osteogenic differentiation in vitro. Implantation of BMP-2-transduced UCB-MSCs led to robust heterotopic bone formation 4 weeks postoperatively, as seen on radiographs and histology. These results, along with the fact that UCB-MSCs can be easily collected with no donor-site morbidity and low immunogenicity, suggest that UCB might be a preferable allogeneic source of MSCs to develop an ex vivo gene-therapy approach to treat difficult bone-repair scenarios.
Introduction
The majority of fractures heal uneventfully, since bone has an excellent intrinsic regenerative potential. However, in cases of difficult biological environments with inadequate blood supply and poor bone stock or fractures with mechanical instability, healing can be compromised, leading to delayed union or non-union. 1 Inadequate bone repair is also seen in settings such as revision total joint replacement and spinal fusion. Autologous bone graft remains the gold standard for the treatment of such cases. However, it is associated with limited graft availability and morbidity at the harvest site, including pain, numbness, infection, and increased risk for fractures. 2 –4 Alternative treatment options include bone allografts, demineralized bone matrixes, cell therapies, and delivery of recombinant growth factors. These options, though effective for certain indications, also have limitations with regards to efficiency, morbidity, and cost for use in the treatment of large bone defects. 5 In particular, recombinant bone morphogenetic protein 2 (rhBMP-2) is a Food and Drug Administration (FDA)-approved osteoinductive agent for clinical use for anterior lumbar spinal fusion and treatment of open tibia fractures. However, it is associated with inconsistent clinical results and several complications due to the supraphysiologic doses needed to induce an adequate biologic response and the rapid release of the protein from the collagen carrier. 6 –8 Thus, there is a clear unmet clinical need for the development of a novel strategy that can consistently promote bone repair in difficult bone-repair scenarios.
Mesenchymal stem cells (MSCs), alone or combined with a growth factor, have been evaluated for their ability to induce bone formation and healing. 9,10 Bone marrow (BM)-derived MSCs first described by Friedenstein et al. 11 are still the most frequently investigated and are often considered the gold standard for cell therapies, but MSCs have been isolated from various other sources, 12 including adipose tissue, 13,14 peripheral blood, 15,16 umbilical cord blood (UCB), 17,18 Wharton's jelly, 19,20 skin, 21 and synovium. 22 UCB has several advantages over other cell therapies as a source of multipotent stem cells for use in regenerative medicine, 23 including sample acquisition and storage, proliferation capacity, and immunologic properties. 24 –26 Since UCB is considered immunoprivileged, and perfect HLA matching is not necessary, 27 UCB could be used as a source of MSCs for allogeneic rather than autologous transplantation, thus allowing for the development of a readily available off-the-shelf product.
Despite the verified osteogenic potential of MSCs in vitro and in animal models of ectopic bone formation, 28 –30 there are concerns that implantation of human MSCs alone in more stringent models of bone repair results in no or limited bone formation. 31 –33 In such cases, progenitor cells combined with an osteoinductive growth factor may be necessary to lead to an adequate biologic response and robust bone formation. Ex vivo regional gene therapy is thus an attractive alternative treatment option, since it allows for the combined delivery of potent cells and an osteoinductive signal to the region of interest (ROI) to promote bone repair. 34 Previous studies from have demonstrated the potential of ex vivo regional gene therapy to promote bone repair in clinically relevant animal models of long-bone fractures, segmental long-bone defects, and spinal fusion. 35 –38
In this study, it was hypothesized that the use of human UCB-MSCs transduced with a lentiviral vector (LV) expressing BMP-2 can lead to sustained production of an osteoinductive signal that will induce robust bone formation in vivo. In brief, the goals of this study were to (1) test the ability of a LV carrying the cDNA for BMP-2 to transduce human UCB-MSCs successfully, leading to abundant BMP production and osteogenic differentiation in vitro, and (2) assess the capacity of BMP-2-transduced UCB-MSCs to induce heterotopic bone formation in a mouse muscle-pouch model. The ultimate goal is to develop ex vivo regional gene therapy using allogeneic UCB-MSC as one aspect of a comprehensive tissue-engineering strategy that can consistently heal challenging bone defects.
Methods
Cell culture
This study used UCB-MSCs that were isolated, cultured, and characterized as previously described. 18 In brief, de-identified UCB samples were collected from full-term deliveries after parental informed consent and Institutional Review Board approval at the New Jersey Cord Blood Bank. Following UCB harvesting, UCB-MSCs were isolated and expanded in culture using a previously published protocol. 18,39 Cells were karyotyped, characterized for cell-surface marker expression and differentiation capacity, 18 and stored in liquid nitrogen for further use. These frozen cells were subsequently thawed, culture expanded, and used to complete this study.
In brief, passage 2 UCB-MSCs were thawed rapidly in a 37°C water bath and then diluted slowly in pre-warmed growth medium. Cells were counted with a hemocytometer using trypan blue and then plated at a concentration of 1 × 106 cells per 10 cm plate in Dulbecco's modified Eagle's medium/Nutrient Mixture F-12 (DMEM/F12) supplemented with 20% fetal bovine serum (FBS; VWR, Radnor, PA) and 1 × penicillin-streptomycin-amphotericin B (Lonza, Walkersville, MD) and maintained in a humidified atmosphere of 5% CO2 at 37°C. Culture medium was changed every 3 days, and non-adherent cells were removed. During each passage, confluent cells were trypsinized and passaged at a density of 0.8–1.0 × 106 cells per plate. Cells were cultured for 2–3 weeks to obtain sufficient numbers for experimental use.
Immunophenotypic characterization
UCB-MSCs were characterized for cell-surface marker expression using multicolor flow cytometric (FACS) analysis. The CD marker panel selected was that of human MSCs, based on the criteria described by the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy. 40 One million cells per donor (passages 4–6) were used for immunophenotypic analysis. After trypsinization, the cells were washed twice and then re-suspended in BD Pharmingen Stain Buffer (BD Biosciences, Franklin Lakes, NJ) at a concentration of 1 × 106 cells/mL. Aliquots of the cell suspension (100 μL) were transferred into 5 mL polystyrene test tubes and stained with one the following antibodies to serve as single-color controls: APC CD73, FITC CD90, PerCP-Cy5.5 CD105, PE CD44, and PE hMSC Negative Cocktail (PE CD45, PE CD34, PE CD11b, PE CD19, and PE HLA-DR). The rest of the cells were split into two tubes. One tube was stained with a combination of the aforementioned markers (hMSC Positive Cocktail and hMSC Negative Cocktail), and the second one was left unstained to serve as a negative control. The tubes were incubated with the antibodies on ice in the dark for 30 min. They were then washed twice and re-suspended in 400 μL BD Pharmingen Stain Buffer (BD Biosciences). Cells were analyzed using a BD LSR II flow cytometer (BD Biosciences). Forward-scatter and side-scatter areas were used to exclude debris and doublets. Compensation was set up using the unstained cells and single-color controls. Data were analyzed using FlowJo software for windows (FlowJo LLC, Ashland, OR).
Transduction with LVs
Two different LVs were used initially to determine optimal transduction conditions for UCB-MSCs (Fig. 1). The first vector was a standard LV encoding BMP-2 or enhanced green fluorescent protein (eGFP) downstream of the RhMLV promoter (LV-BMP-2 or LV-GFP). 41 –43 In order to enhance transgene expression further, a two-step transcriptional amplification (TSTA) lentiviral system was constructed (LV-TSTA-BMP-2 or LV-TSTA-GFP). The TSTA system requires two different vectors: the GAL4-VP16 transactivator vector (LV-RhMLV-GAL4-VP16) and the transgene expression vector encoding the G5 promoter and the BMP-2 or eGFP cDNA (LV-G5-BMP-2 or LV-G5-GFP), under the control of the GAL4 responsive promoter. 44,45 All LV stocks were generated by transfecting 293T cells (American Type Culture Collection, Manassas, VA) and were subsequently titered by quantifying p24 protein contents in vector solution by enzyme-linked immunosorbent assay (ELISA; PerkinElmer, Waltham, MA), as previously described. 41,44 The titers of all vector stocks were approximately 0.5–4 × 108 transducing units/mL.
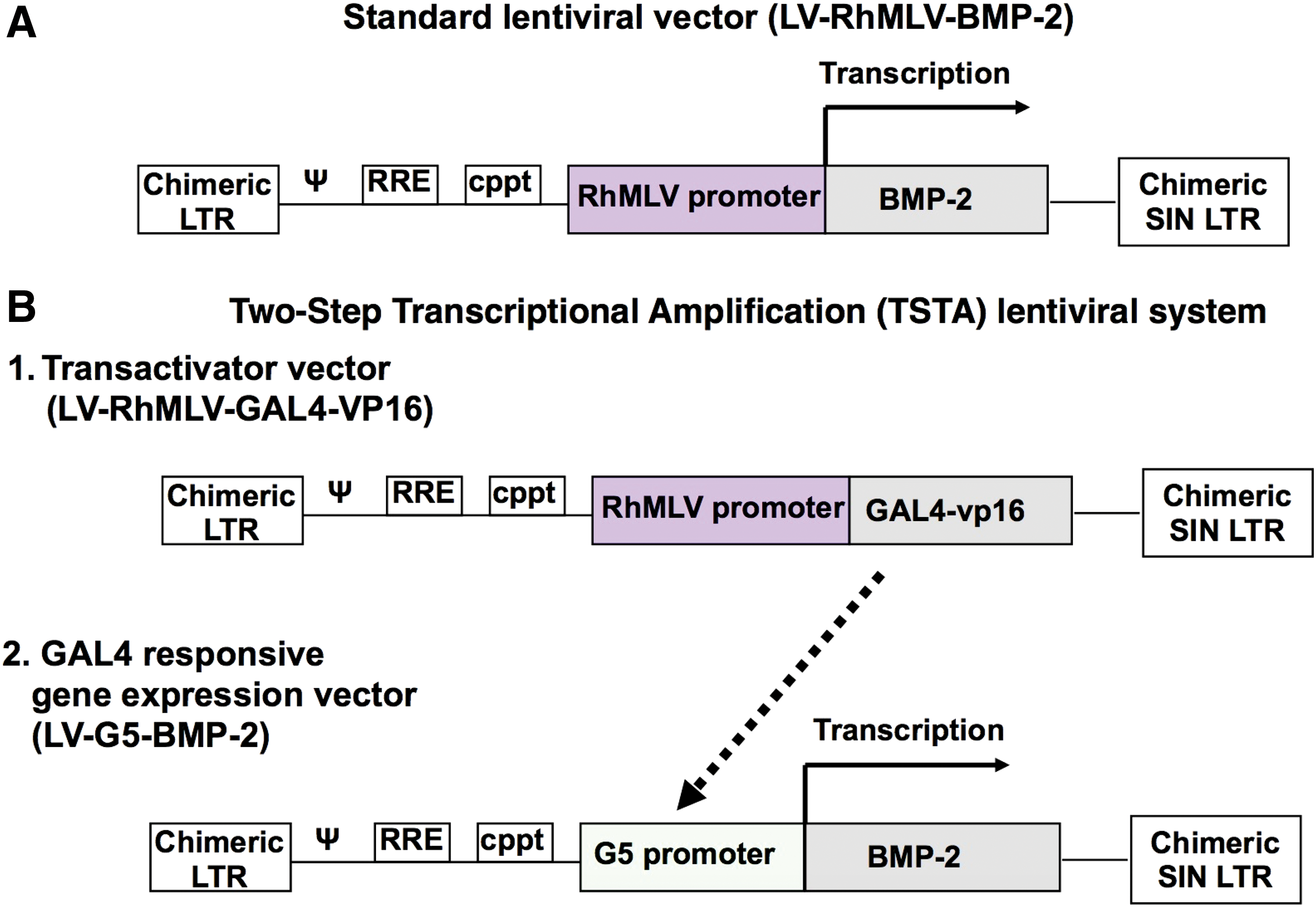
Schematics of the lentiviral vectors (LVs) used to overexpress BMP-2, namely
Transductions were carried out at 37°C, 5% CO2, in the presence of 8 μg/mL polybrene at a multiplicity of infection (MOI) of 5, 10, and 25 for LV-BMP-2 and MOIs of 2, 5, and 25 for LV-TSTA-BMP-2. After overnight incubation, the transduction medium was aspirated, and cells were washed with phosphate-buffered saline (PBS) three times to remove any extracellular viruses.
ELISA for BMP-2 production in vitro
In vitro BMP-2 production by BMP-2-transduced UCB-MSCs during a 24 h period was quantified 48 h after transduction using an ELISA assay (Quantikine; R&D Systems, Minneapolis, MN). In brief, following the overnight transduction period, BMP-2-transduced UCB-MSCs were washed and then re-plated in fresh DMEM/F12. After incubation for an additional 24 h, the culture medium was harvested for quantification of BMP-2. Each sample was run in triplicate. The BMP-2 production was standardized by cell number and reported as nanograms of BMP-2 per 24 h by 1 × 106 cells.
Transduction efficiency using FACS analysis
To determine transduction efficiency, UCB-MSCs were transduced with LV-eGFP at a MOI of 25 and then attained for FACS analysis 2, 7, and 14 days post transduction. Non-transduced UCB-MSCs were used as control. Cells were re-suspended in PBS +1% FBS at a concentration of 1 × 106 cells/mL and then analyzed with a BD LSR II flow cytometer (BD Biosciences). Forward-scatter and side-scatter areas were used to exclude cell debris and doublets. The percentage of eGFP+ cells was determined using FlowJo software (FlowJo LLC).
Osteogenic differentiation potential
Transduction with LV-BMP-2 was used to assess the in vitro osteogenic potential of the cells. Non-transduced cells were used as negative control. Human adipose-derived stem cells (ASCs) were used as a positive control. A total of six different UCB samples were evaluated. Cells were seeded in duplicate at a concentration of 105 cells/well on a 12-well plate. Osteogenic differentiation was induced by culturing the cells in DMEM enriched with 10% FBS, antibiotics/antimycotics, 0.1 μM dexamethasone, 50 μg/mL L-ascorbic acid, and 10 mM β-glycerophosphate. Seven days later, the cells were fixed with 10% formaldehyde for 10 min, washed with PBS, and stained with 2% Alizarin Red S solution for 30 min. After aspiration of the unincorporated dye, the wells were washed with double-distilled water, re-aspirated, and then stored at −20°C prior to dye extraction and spectrophotometric evaluation. Alizarin Red–stained UCB-MSCs cultures were imaged using an EVOS XL inverted microscope (AMG, Mill Creek, WA).
Quantification of Alizarin Red S was performed by a colorimetric assay, as previously described. 46 In brief, cells were incubated with 10% acetic acid for 30 min at room temperature with gentle shaking and then scraped from the plate with a cell scraper and transferred to a 1.5 mL Eppendorf tube. After vortexing for 30 s, the sample was heated at 85°C for 10 min and then cooled on ice for 5 min. The stained layer was centrifuged at 20,000 g for 15 min. An aliquot of the supernatant was transferred into a new 1.5 mL tube and mixed with 200 μL ammonium hydroxide. Small aliquots (150 μL) of the resulting mixture were then placed in triplicate on a 96-well plate and read by a plate reader at an optical density of 405 nm.
Hind-limb muscle-pouch model
Finally, the in vivo bone induction capacity of UCB-MSCs was tested in a mouse hind-limb muscle-pouch model 47,48 after Institutional Animal Care and Use Committee approval. All experiments conformed to all relevant regulatory standards. UCB-MSCs from three different donors were used. NOD SCID gamma mice (NSG; Cannon Lab, Keck School of Medicine, University of Southern California) were used in order to avoid an immune reaction due to the use of human cells. Briefly, after administration of 2% isoflurane, the left limbs of the mice were shaved and prepared with three alternating scrubs. Using an aseptic surgical technique, an incision was made on the posteromedial aspect of the thigh, and a pouch was created using blunt dissection between the hamstring muscles. UCB-MSCs (2 × 106) were loaded on a collagen sponge and then implanted into the surgically created muscle pouch. The muscle layer and skin incision were closed with absorbable sutures. The mice received adequate analgesia and prophylactic antibiotics in the immediate postoperative period. They were allowed to bear weight immediately and to eat and drink ad libitum. The following groups were assessed: UCB-MSCs transduced with LV-BMP-2 (group I), UCB-MSCs transduced with LV-eGFP (group II), and non-transduced UCB-MSCs (group III). Five mice were used per group.
Bone formation was assessed radiographically and histologically. Plain radiographs were performed at 2 and 4 weeks postoperatively using an Ultrafocus60 X-ray device (Faxitron Bioptics, Tucson, AZ). The mice were then euthanized (4 weeks postoperatively), and their limbs were harvested and processed for standard decalcified histology and quantitative histomorphometric analysis. In brief, the specimens were first fixed in 10% formalin (24 h), followed by EDTA decalcification for 2 weeks, and then paraffin embedding. All samples were cut longitudinally, loaded on glass slides, and stained with either hematoxylin and eosin or Masson's trichrome. Slides were imaged with an AxioImager A2 (Carl Zeiss Microscopy, LLC, Thornwood, NY) and analyzed with Bioquant (Bioquant Image Analysis, Nashville, TN). Bone formation was quantified in Masson's trichrome sections at 2.5 × magnification, with the ROI selected to include the whole implant, along with any newly formed bone, in the muscle pouch. The designated ROI represented the tissue area (TA). The amount of new bone within this area was then quantified (bone area [BA]). Finally, the BA/TA ratio was calculated for each sample.
Statistical analysis
Statistical analysis was performed using IBM SPSS Statistics for Windows v22 (IBM Corp, Armonk, NY). All data are reported as the mean and standard deviation. The significance level was set at 0.05. An independent-samples Student's t-test was done to compare BMP-2 production between BMP-2-transduced and non-transduced UCB-MSCs. Mann–Whitney's U-test was used to compare extracellular mineralization potential between the cells. One-way analysis of variance and subsequent post hoc analysis with Tukey's range test were used for between-groups comparisons with regards to histomorphometry results.
Results
Profile of surface markers
Immunophenotypic analysis of UCB-MSCs revealed high expression of the typical MSC markers CD44 (99 ± 0.4%), CD73 (94.7 ± 4.9%), and CD90 (91.7 ± 2.4%). However, expression of CD105 was limited (10.2 ± 2.3%). Finally, UCB cells displayed minimal (<1%) expression of hematopoietic markers (CD45, CD11b, CD19, and HLA-DR; Fig. 2).
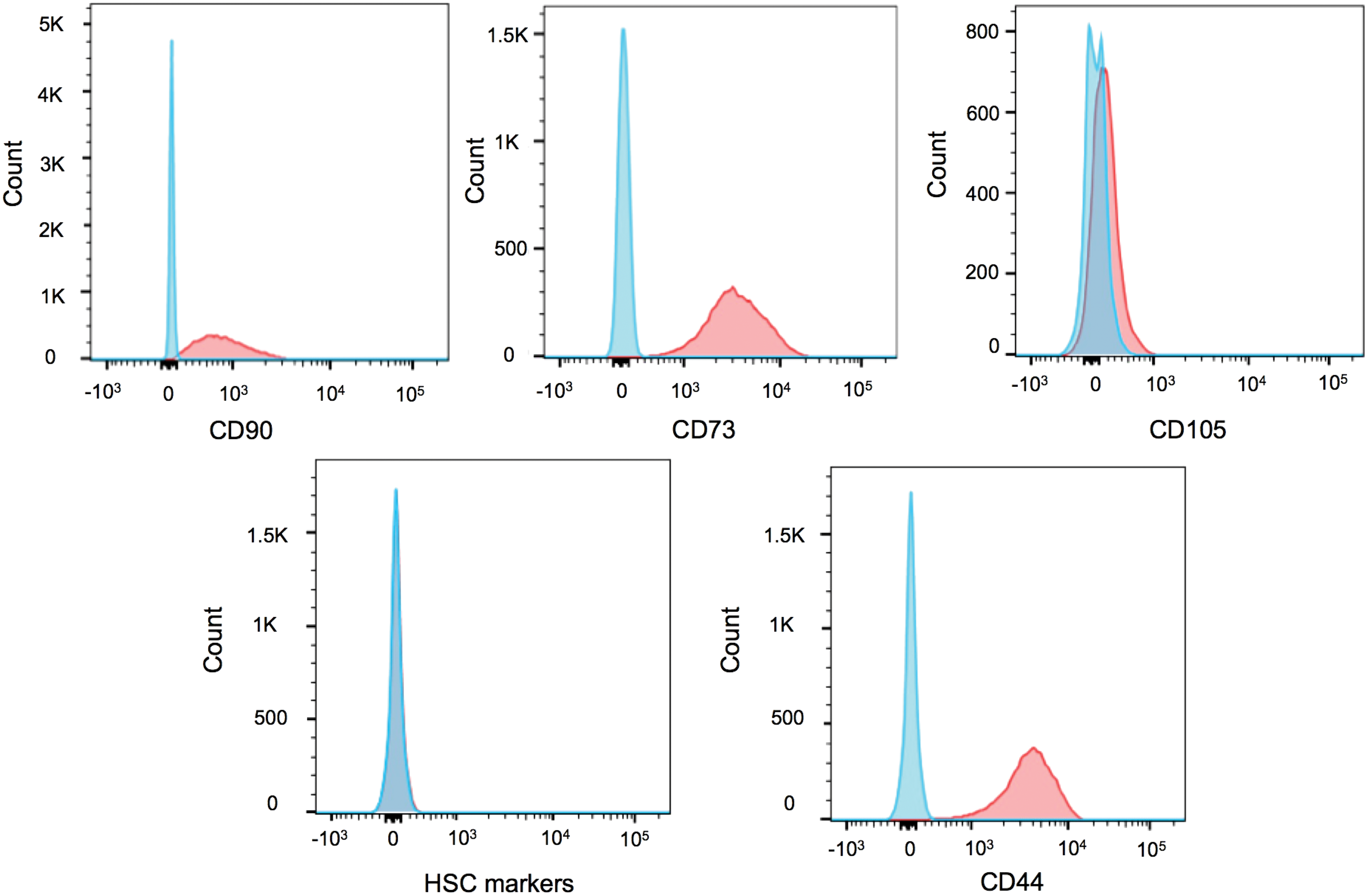
Umbilical cord blood–derived mesenchymal stem cells (UCB-MSCs) were characterized for cell-surface markers using a four-color flow cytometric (FACS) analysis. Unstained cells were used as control. Proper gating was first done to exclude cell debris and doublets. Histograms depicting individual expression of CD90, CD73, CD105, CD44, and hematopoietic stem-cell markers from one representative donor are given. Red histogram, stained UCB-MSCs; blue histogram, unstained control UCB-MSCs. Color images are available online.
In vitro BMP-2 production
Optimal transduction conditions were determined by BMP-2 production and cytotoxicity induced by serial MOIs with the two different LVs. Transduction of UCB-MSCs with either LV-TSTA-BMP-2 or LV-BMP-2 was associated with abundant BMP-2 production, with the highest levels being observed at a MOI of 25 for both vectors. Although the LV-TSTA-BMP-2 vector demonstrated higher BMP-2 production at all MOIs compared to the standard LV-BMP-2 vector, 80–96% of the cells died secondary to the toxicity of the TSTA virus 7 days following transduction. On the contrary, minimal cytotoxicity was observed in LV-BMP-2 transductions. Therefore, a decision was made to use the standard LV-BMP-2 vector at a MOI of 25 for further experiments in this study, since adequate BMP production could be obtained without causing significant viral toxicity-related cell death (Supplementary Table S1).
Following selection of optimal vector and MOI, UCB samples from four different donors were used to determine efficient transduction and levels of BMP-2 production after transduction with LV-BMP-2. ELISA showed that at a MOI of 25, 1 × 106 UCB-MSCs produced 40.50 ± 18.84 ng of BMP-2 in a 24 h period. In contrast, minimal BMP-2 was produced by non-transduced UCB-MSCs (0.36 ± 0.14 ng/24 h/106 cells; p < 0.001 vs. transduced cells).
Transduction efficiency
The percentage of eGFP-expressing cells was determined at 2, 7, and 14 days post transduction with LV-eGFP at a MOI of 25 using FACS analysis. Abundant eGFP expression was noted in all of the LV-eGFP-transduced samples, with an average transduction efficiency of 99 ± 0.35% and a mean fluorescence intensity of 13,540 ± 4,112 two days following transduction. eGFP expression remained strong at 2 weeks, with >95% of cells being eGFP+ in all samples (Fig. 3).

Gene expression analysis of UCB cells transduced with a LV encoding enhanced green fluorescent protein (eGFP) at a multiplicity of infection of 25.
In vitro osteogenic potential
The in vitro osteogenic capacity of UCB-MSCs, alone or transduced with LV-BMP-2, was assessed using Alizarin Red staining following culture in osteogenic media for 7 days. All of the non-transduced cell cultures demonstrated minimal or no calcified extracellular matrix deposition. In contrast, a great deal of variability was observed in the BMP-2-transduced UCB samples with regards to in vitro osteogenic potential, ranging from abundant extracellular calcium deposition to minimal osteogenic differentiation. Nevertheless, overall, LV-BMP-2-transduced cells showed a higher mineralization potential compared to non-transduced cells. This difference was confirmed with spectrophotometric quantification of Alizarin Red, which showed higher in vitro extracellular deposition of calcium in BMP-2-transduced UCB cell cultures (p = 0.041; Fig. 4). Finally, UCB/LV-BMP-2 demonstrated similar extracellular calcium deposition compared to ASC/LV-BMP-2 (1.03 ± 1.32 vs. 0.91 ± 0.10; p = 0.548).
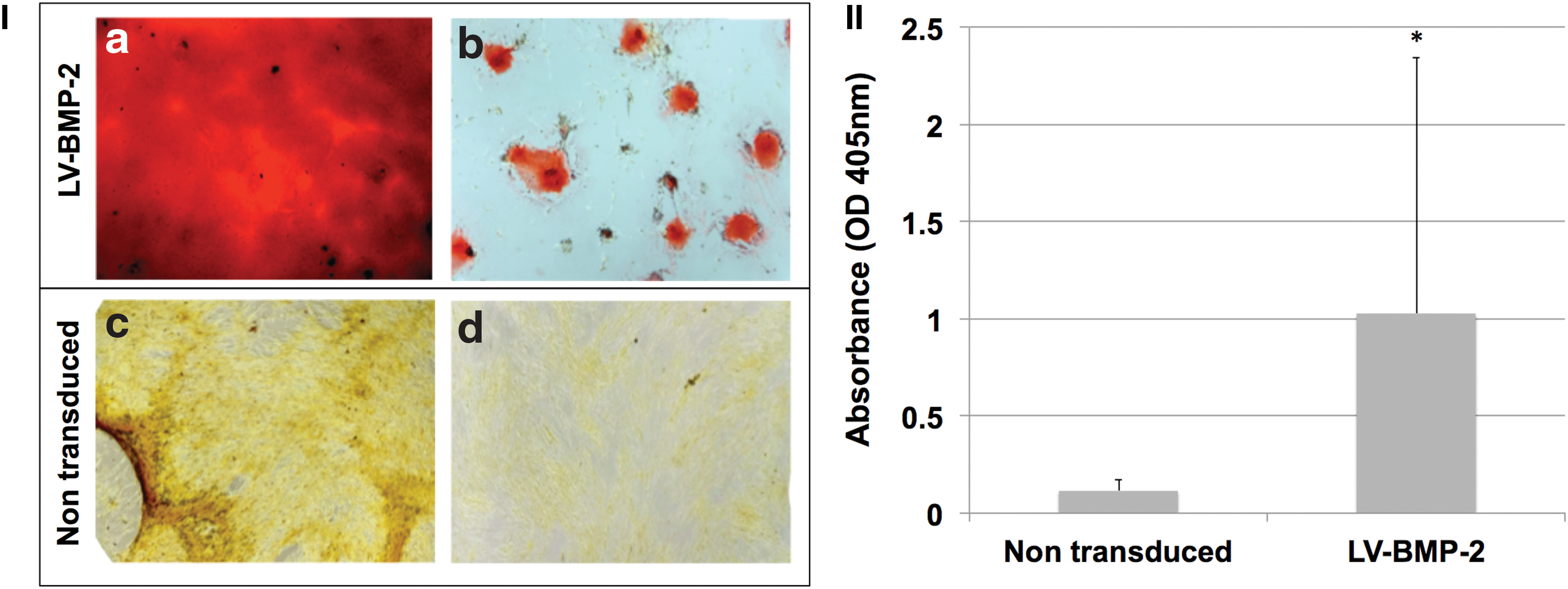
In vitro osteogenic potential of passages 4–6 non-transduced and LV-BMP-2 transduced UCB-MSCs, as seen with Alizarin Red staining following culture in osteogenic medium for 7 days.
In vivo bone formation potential
In vivo implantation of LV-BMP-2-transduced UCB-MSCs loaded onto a collagen sponge in a SCID mouse muscle pouch led to robust bone formation at 2 and 4 weeks postoperatively, as seen on plain radiographs. No bone was noted at any time point in the negative control groups implanted with either LV-GFP-transduced or non-transduced UCB-MSCs (Fig. 5).
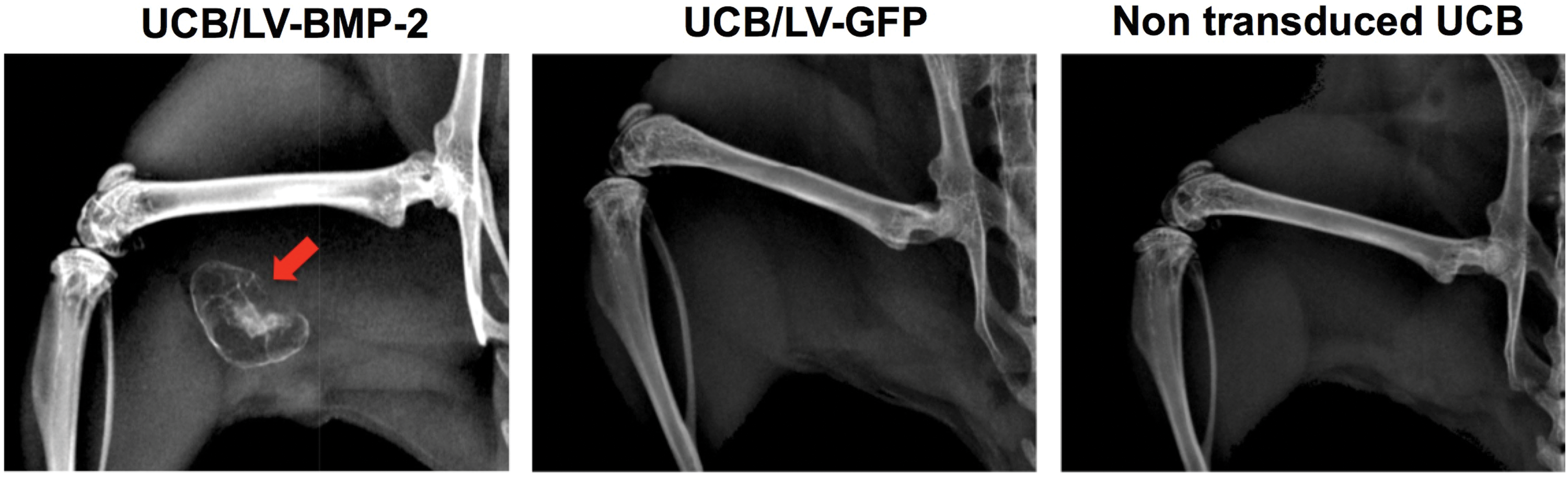
Representative 4-week radiographs. Abundant bone formation is noted in the muscle pouch implanted with UCB/LV-BMP-2 cells (red arrow). No bone is seen in the animals treated with GFP-transduced or non-transduced UCB-MSCs. Color images are available online.
Histologic analysis revealed formation of woven bone with reconstitution of the BM cavity in the muscle pouch 4 weeks after implantation with UCB/LV-BMP-2. In contrast, no evidence of ectopic bone formation was noted in the control groups (Fig. 6).
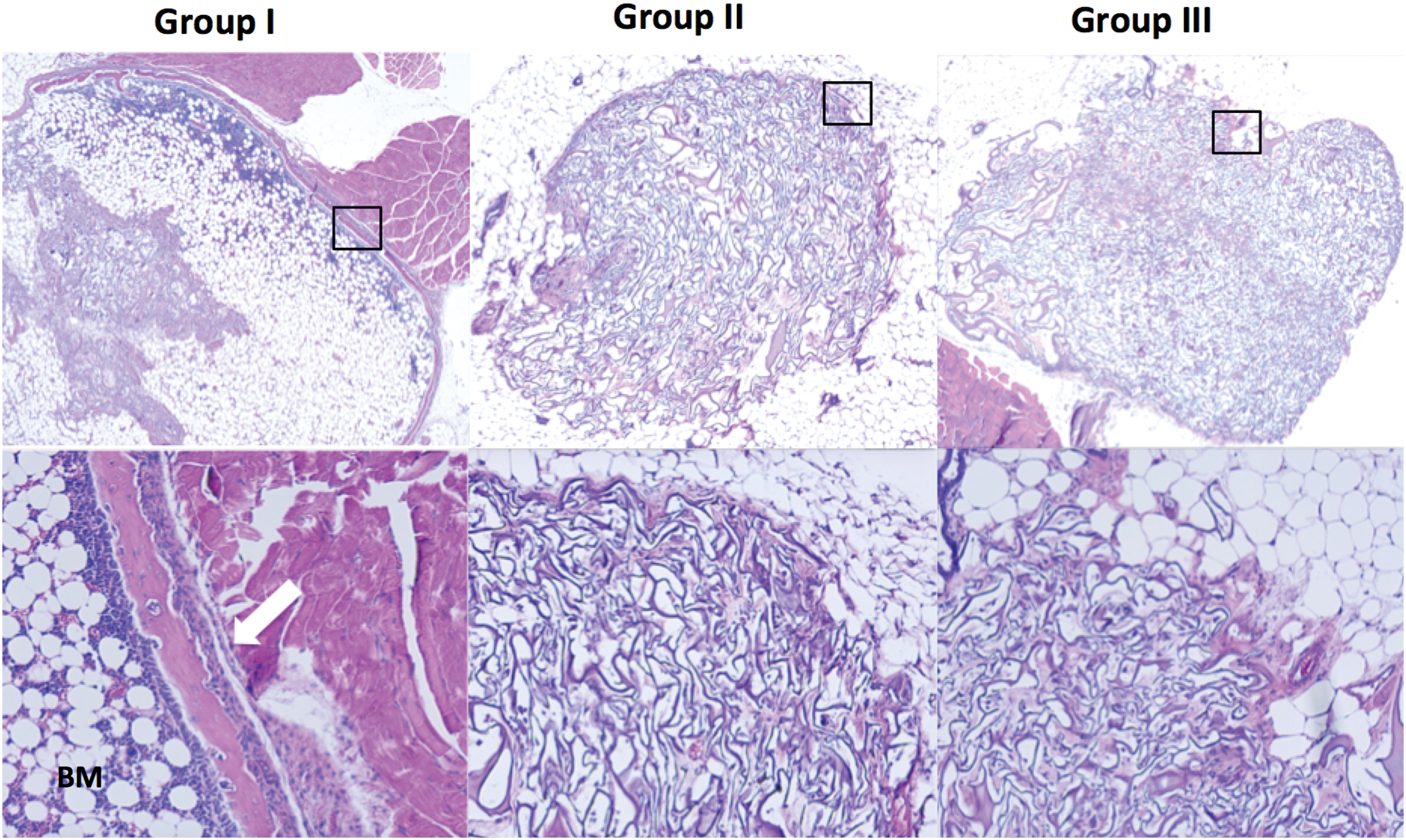
Representative hematoxylin and eosin–stained sections 4 weeks after implantation of human UCB-MSCs. The top image is a transverse section of the implant (1 × magnification), and the bottom image is a 10 × magnification of the black rectangle. Mature osseous components, including woven bone (white arrow) and bone marrow, were observed in mice implanted with LV-BMP-2-transduced cells. No bone formation was induced by implantation of non-transduced or GFP-transduced UCB-MSCs. Color images are available online.
Quantitative histomorphometric analysis of the Masson's trichrome–stained slides (Supplementary Table S2) confirmed the differences between the groups in bone formation demonstrated by X-rays and qualitative histology. Mean BA/TA was significantly higher in mice treated with LV-BMP-2-transduced cells (6.62 ± 1.89%) compared to animals treated with eGFP-transduced UCB-MSCs (1 ± 0.22%; p = 0.001) or non-transduced UCB-MSCs (0.12 ± 0.12%; p = 0.001). No significant difference with respect to BA/TA was shown between animals treated with either non-transduced or eGFP-transduced UCB-MSCs (p > 0.05; Fig. 7).
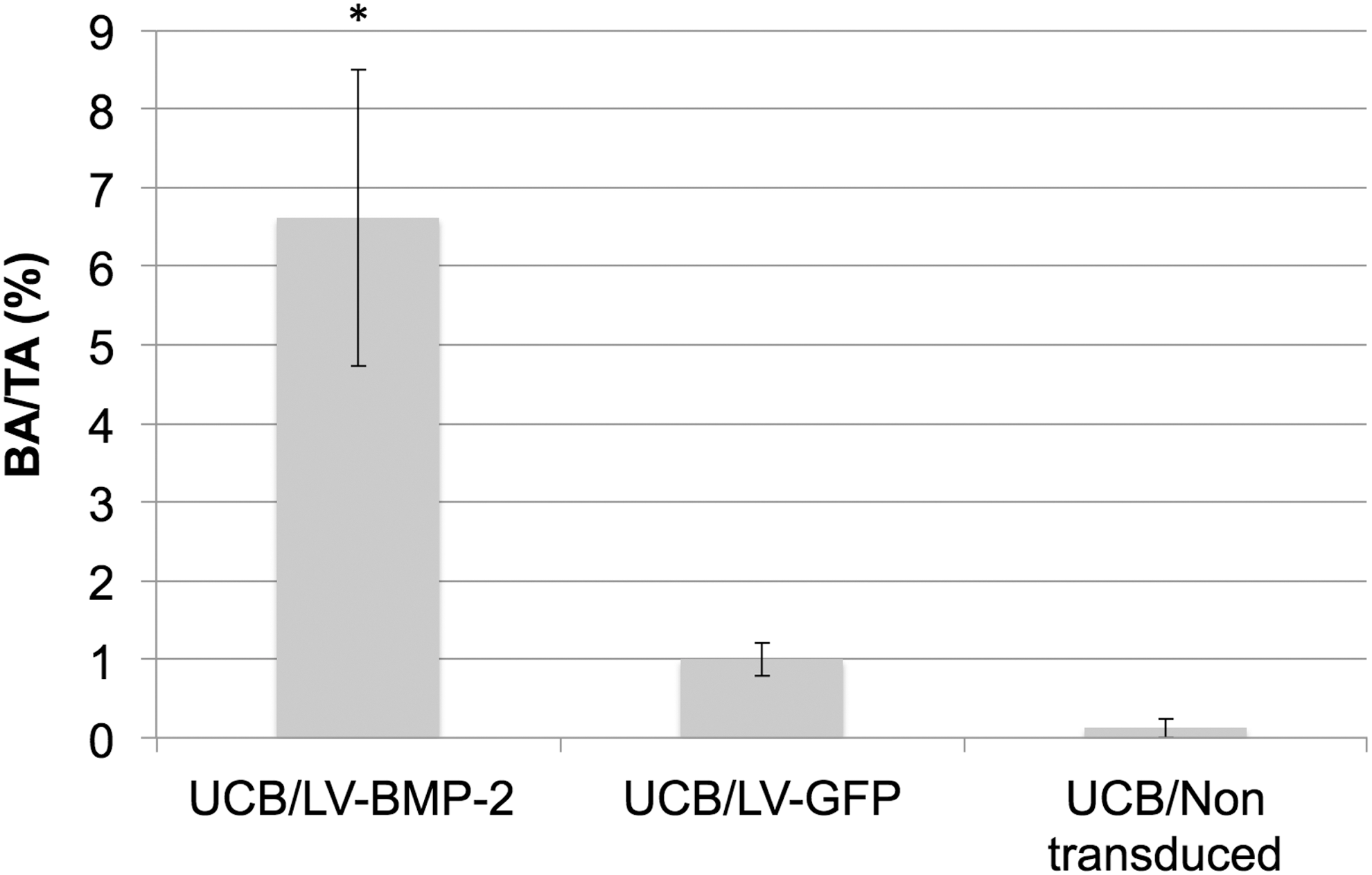
Quantitative histomorphometric analysis of Masson's trichrome–stained sections of the mouse muscle pouch 4 weeks postoperatively (1 × magnification). Mean BA/TA was significantly higher in mice treated with LV-BMP-2-transduced cells compared to animals treated with GFP or non-transduced UCB-MSCs. *p = 0.001.
Discussion
MSCs have been extensively studied for their unique self-renewal and multilineage differentiating features and their potential to improve current treatment regimens in regenerative medicine. 49 Multiple independent laboratories have focused on the potential of human MSCs, isolated from various sources, to differentiate into an osteogenic phenotype in vitro and in vivo. 49,50 However, the use of MSCs alone has not led to clear-cut results in terms of tissue regeneration. 50,51 Human MSCs have been reported to promote osteogenesis in animal models of heterotopic bone formation 52 –56 or calvarial bone defects, 57,58 but have had limited success in more stringent models such as long-bone critical-sized defects. 31 –33,59,60 In clinical studies implantation of autologous BM aspirate or BM MSCs has been reported to lead to enhanced bone regeneration in patients with long-bone non-unions. 61 –64 Although promising, the efficacy of this strategy has not been definitely determined because of the limited number of patients studied and the lack of randomized trials. 50
This study demonstrated that MSCs from human UCB transduced with a LV-BMP-2 vector can induce heterotopic bone formation in a muscle-pouch model. It is hypothesized that in clinical bone-repair scenarios with limited biologic potential (i.e., segmental bone defects), MSCs combined with an osteoinductive stimulus may significantly enhance bone repair. Ex vivo regional gene therapy is a promising tissue-engineering approach that allows for the delivery of both osteoprogenitor cells and osteoinductive growth factors. The advantage over recombinant proteins is that it is associated with a sustained long-term release of the osteoinductive signal lasting for >12 weeks. 42 Another treatment option is in vivo gene therapy. The direct injection of a virus or a plasmid is relatively simple. However, a disadvantage of this strategy in the management of large bone defects or in a setting of a compromised biological environment is that there may be a limited number of local osteoprogenitor cells that are available to be transduced. In contrast, ex vivo gene therapy allows for higher transduction efficiency, direct targeting of a specific cell population of interest, and implantation of the transduced cells into a specific anatomic site. 34,37 Moreover, it is safer, since systemic injection of a viral vector is not needed; target cells are harvested, processed, and genetically manipulated outside of the body prior to implantation. Finally, it has been hypothesized that after transduction, these transduced stem cells not only act as delivery vehicles for the target gene recruiting host osteoprogenitor cells, but also express an osteogenic phenotype and directly participate in bone repair themselves. Over the past few years, gene therapy has emerged as a promising new treatment strategy for the management of hematological and non-hematological malignancies, 65,66 metachromatic leukodystrophy, 67 and Leber congenital amaurosis, 68 with chimeric antigen receptor recently obtaining FDA approval for clinical use in adults and children with relapsed or refractory B-cell precursor acute lymphoblastic leukemia. 69 Therefore, gene therapy has the potential to be adapted for the treatment of musculoskeletal problems and could revolutionize the treatment of large bony defects. The optimal cell type for use in ex vivo gene therapy for bone repair has yet to be identified.
Autologous BM and adipose tissue have significant potential as sources of adult stem cells for use in cell-based tissue-engineering therapies. A potential limitation of this strategy is that it would be difficult to develop an off-the-shelf allogeneic product because of concerns of inducing an immunogenic response to the recipient following cell implantation.
UCB has been explored as an alternative source of multipotent stem cells for use in regenerative medicine 23 and the treatment of hematologic and oncologic diseases. 70 UCB-MSCs have several advantages over other stem-cell sources. First, the collection of cord blood is easy and noninvasive. UCB is readily available, as it is often discarded after childbirth. Moreover, the collection of UCB happens postpartum and thus does not pose any additional risks to the fetus or the mother, hence eliminating any ethical concerns associated with embryonic stem cells. Cord-blood units can be stored in advance and are therefore rapidly available when needed. 25 UCB-MSCs demonstrate a higher proliferation capacity and no signs of senescence over multiple passages compared to MSCs from other sources. 18 Finally, UCB grafts are considered less immunogenic compared to BM and peripheral blood stem cells, and thus less stringent human leukocyte antigen (HLA) matching is required. 71 –73 Currently, UCB-MSCs are used clinically for malignant 72,73 and nonmalignant 74 –77 conditions in pediatric and adult patients in the absence of a suitable HLA identical sibling donor or HLA-matched BM unrelated donor. 78 Therefore, UCB could be used as a source of MSCs for allogeneic rather than autologous transplantation, providing an off-the-shelf cell delivery vehicle for use in gene-therapy applications for bone regeneration.
UCB-MSCs have been previously demonstrated to possess osteogenic differentiation potential in vitro, even similar to that of ASCs and BMSCs, 24,79 suggesting that these cell types could be used interchangeably for bone-regeneration applications. Moreover, it has been shown that pre-differentiated UCB-MSCs seeded on a collagenous bone matrix or demineralized bone matrix could lead to formation of mineralized bone-like tissue in a rat paravertebral muscle pouch or rat calvarial defect model. 80,81 However, the combined delivery of non-differentiated UCB-MSCs and an osteoinductive factor via a gene-therapy approach has never been tested before for induction of bone formation.
This study demonstrated that human MSCs derived from UCB can be transduced successfully with a LV carrying the cDNA for BMP-2. UCB-MSCs alone cultured in osteogenic media for 1 week consistently failed to differentiate toward an osteogenic lineage in vitro. In contrast, transduction with the BMP-2 LV led to abundant BMP-2 production and induction of an osteogenic phenotype in vitro, albeit at varying degrees. This difference in the in vitro osteogenic differentiation could indicate the heterogeneity of MSCs between donors or preparations or be suggestive of the overall health of the cells. Moreover, it could be due to the fact that UCB-MSCs require a generally longer period of time to differentiate in vitro (3–4 weeks), even in the presence of BMP-2, as previously described. 82
When implanted into the hind-limb muscle pouch of NSG mice, BMP-2-transduced UCB-MSCs were able to induce abundant heterotopic bone formation, as seen on plain radiographs and histologic analysis. In contrast, groups treated with non-transduced UCB-MSCs or GFP-transduced cells did not show any evidence of in vivo bone formation, clearly showing that transduction with LV-BMP-2 can significantly enhance the osteogenic potential of human UCB-MSCs. Interestingly, the in vitro variability in osteogenic differentiation potential between donors did not translate to analogous differences in vivo. When transduced with LV-BMP-2, both the donor that performed best (Donor 3) and the one that performed worst (Donor 4) in the in vitro Alizarin Red assay were associated with successful bone formation as early as 2 weeks following cell implantation in vivo. Moreover, quantitative histomorphometric analysis demonstrated similar mean BA and BA/TA in both donors. These results suggest that though useful, in vitro results are not necessarily indicative of what happens in an in vivo environment of bone formation. The interaction between donor UCB-MSCs and host cells may have pushed the implanted cells toward an osteogenic phenotype in vivo, despite their inability to differentiate in vitro. Another hypothesis is that the regenerative effects of UCB-MSCs were not exclusively associated with the donor cells' osteogenic differentiation potential, but mostly with their contributions to the initiation of the healing process by producing abundant amounts of BMP-2 so as to activate the host's osteoprogenitors successfully to induce bone formation.
Despite the promising results demonstrated with the use of LV-BMP-2 transduced UCB-MSCs to promote bone repair, this study had some limitations. First, the in vivo bone formation in the muscle pouch was not assessed with micro computed tomography. However, both radiographs and histologic and histomorphometric analyses showed successful bone formation in the BMP-2-UCB/MSCs implanted animals and none in the other two groups. Second, this ex vivo gene-therapy strategy was assessed in a mouse hind-limb muscle-pouch model. Though excellent as a proof-of-concept model, a more stringent model of orthotopic bone formation, such as a critical-sized long-bone defect, is needed to assess the translational potential of this treatment regimen further. Finally, the exact role of the transduced UCB-MSCs in the bone-formation process could not be definitively established in this particular model. The transduced cells produce BMP-2, which clearly induced bone formation. However, it could not be determined whether the cells themselves participated in the osteogenesis.
In conclusion, the current study provides proof of concept for potential clinical applications of BMP-2-transduced UCB-MSCs. To the authors' knowledge, this is the first report of LV-mediated gene therapy using allogeneic human UCB-MSCs to promote bone repair. The ultimate goal is to develop ex vivo regional gene therapy using UCB-MSCs as an off-the shelf product that can consistently heal challenging bone defects. These results, along with the facts that UCB-MSCs can be easily collected with no donor-site morbidity and demonstrate a significantly lower immunogenicity compared to other sources of MSCs, indicate that UCB might be a preferable allogeneic source of MSCs to develop an ex vivo regional gene-therapy approach to treat difficult bone-repair scenarios.
Footnotes
Acknowledgments
This work was supported by a National Institutes of Health grant to J.R.L. (R01AR057076). The authors would like to thank Dr. Paula Cannon and Evan Lopez (Keck School of Medicine, Department of Molecular Microbiology and Immunology) for providing the NSG animals needed for the study from their mouse colony. Part of the project was presented as a poster in the ASGCT 21st Annual Meeting, May 16–19, 2018.
Author Disclosure
J.R.L. has received royalties and has served as a paid consultant for Depuy, is a shareholder in Hip Innovation Technologies, Inc., and has received royalties, financial, or material support from Elsevier. S.B., B.S., O.S., A.H.T. J.E., and D.E. have no conflicts to report.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.