
Abstract
The majority of advanced breast cancer patients develop distal metastasis, including lung and bone metastasis. However, effective therapeutic strategies to prevent metastasis are still lacking. Decorin is a natural inhibitor of transforming growth factor β, which plays a pivotal role in tumor metastasis. An oncolytic adenovirus expressing decorin, rAd.DCN, has been developed previously. In an immune-competent breast tumor (4T1) model, intratumoral (i.t.) as well as intravenous (i.v.) delivery of rAd.DCN inhibited growth of orthotopic tumors and spontaneous lung metastasis. It was shown that i.t. delivery of rAd.DCN produced higher levels of transgene expression and evoked stronger oncolysis of the tumors compared to i.v. delivery. However, i.v. delivery resulted in higher amount of virus accumulation in the lungs and produced stronger responses to prevent tumor lung metastasis. Oncolytic adenovirus-mediated decorin expression in the tumors downregulated the decorin target genes and decreased epithelial mesenchymal transition markers. Decorin expression in lung tissues also increased Th1 cytokine expression, such as interleukin (IL)-2, IL-12, and tumor necrosis factor α, and decreased Th2 cytokines, such as transforming growth factor β and IL-6. Moreover, rAd.DCN treatment induced strong systemic inflammatory responses and upregulated CD8+ T lymphocytes. In conclusion, rAd.DCN inhibits tumor growth and lung metastasis of breast cancer via regulating wnt/β-catenin, vascular endothelial growth factor (VEGF), and Met pathways, and modulating the antitumor inflammatory and immune responses. Considering that i.v. delivery was much more effective in preventing lung metastasis, systemic delivery of rAd.DCN might be a promising strategy to treat breast cancer lung metastasis.
Introduction
B
Decorin, a member of the small leucine-rich poteoglycan family, has been suggested to be a natural inhibitor of TGF-β signaling. 12 Decorin can not only regulate the proliferation, migration, and invasion of tumor cells but can also reprogram the tumor microenvironment and modulate inflammatory responses and angiogenesis. 13,14 Moreover, decorin also evokes endothelial cell autophagy via AMPKα, and triggers mitophagy of tumor cells in a mitostatin-dependent manner. 6,15,16 An oncolytic adenovirus expressing decorin, Ad.DCN, has been also developed previously, and it has been shown that it could significantly inhibit breast cancer bone metastasis in an immune-deficient MB-MDA-231 model. 6 In an immune-competent murine colon cancer (CT26) xenograft model, rAd.DCN, a telomerase reverse transcriptase promoter (TERTp) controlled oncolytic adenovirus, inhibited tumor growth and lung metastasis via immune activation. 5 Therefore, there was an interest in evaluating the therapeutic effects of rAd.DCN in treating breast cancer and its lung metastasis in immune-competent models.
This study established a breast cancer orthotopic xenograft model (4T1-luc) in immune-competent BALB/c mice. Intratumoral (i.t.) and intravenous (i.v.) delivery of rAd.DCN significantly inhibited tumor growth and lung metastasis. However, i.v. delivery was much more effective in preventing tumor lung metastasis. rAd.DCN-mediated decorin expression in the tumors and lung tissues downregulated the expression of decorin target genes, inhibited epithelial mesenchymal transition (EMT), and promoted inflammatory responses. These studies suggest that rAd.DCN can be developed for the treatment of breast cancer metastases.
Materials and Methods
Cell lines and adenoviruses
The mouse mammary gland cancer cell line 4T1 was purchased from American Type Culture Collection (ATCC, Manassas, VA). 4T1 cells were transfected with lentiviral vector containing luciferase gene (PLKO.1-CMV-luciferase) to generate luciferase-expressing 4T1 cells, 4T1-luc. Both 4T1 and 4T1-luc were maintained in RPMI-1640 media (Gibco, Gaithersburg, MD) plus 10% fetal calf serum (FCS; Hyclone, Logan, UT). Human embryonic kidney cells, HEK293, were obtained from ATCC and cultured in Dulbecco's Minimal Essential Medium (Gibco) supplemented with 10% FCS.
Oncolytic adenoviruses, rAd.DCN and rAd.Null, in which the expression of E1A protein was controlled by TERTp, were constructed and prepared using a simplified system for generating oncolytic adenovirus vector carrying one or two transgenes, as described previously. 17 The expression of decorin was confirmed by Western blotting and real-time reverse transcription polymerase chain reaction (RT-PCR).
4T1 orthotopic xenograft model and treatment with oncolytic adenoviruses
All the procedures for the animal experiments were approved by the Animal Care and Use Committee of the Beijing Institute of Radiation Medicine. To establish the orthotopic xenograft model of breast cancer, 6.5 × 105 4T1-luc cells (100 μL) were injected into number 3 and number 4 mammary fatty pads of BALB/c mice (4–6 weeks old). When tumors were visible (on day 7 after cells injection), the tumors were measured with a caliper, and the volume was calculated using the formula: tumor volume = width 2 × length/2. The tumor burden was also analyzed by real-time bioluminescence imaging (BLI).
Tumor-bearing mice were divided into six groups without statistical difference. On day 7, phosphate-buffered saline (100 μL), rAd.Null, and rAd.DCN (2.5 × 1010 vp/100 μL/injection) were administrated i.t. (Buffer, rAd.Null, and rAd.DCN groups; n = 15 mice/group) or i.v. (Buffer, rAd.Null, and rAd.DCN groups; n = 15 mice/group). A repeat injection was administered on day 10. The tumor volumes were monitored on days 7, 11, 15, 18, and 25, and the tumor burdens were analyzed by BLI on days 7, 14, and 25. On day 25, the mice were euthanized, and the tumors were harvested and weighed.
Histopathological analysis and immunohistochemistry
On day 12, five mice from each group were euthanized for histopathological analysis. The other mice were euthanized at the terminal time point (day 25). The tumors, liver, and lungs were harvested, processed, and stained with hematoxylin and eosin. The tumor lesions (metastasis) in the lungs were counted and analyzed. Moreover, the proliferation of tumor cells after treatment with oncolytic adenoviruses was detected by Ki67 (Abcam, Cambridge, United Kingdom) staining on day 12. Furthermore, on day 25, the cellular apoptosis in the tumor tissues was examined by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL; Promega, Madison, WI) according to the manufacturer's instructions.
Distribution of adenoviruses and the expression of decorin
On days 12 and 25, the tumor tissues and lungs were processed and stained with mouse anti-human adenovirus antibody (Abcam, Cambridge, MA) and goat anti-human decorin antibody (R&D systems, Minneapolis, MN). The expression of adenovirus structural proteins and decorin protein were observed under an inverted microscope.
Decorin regulates its target genes, EMT markers, and inflammatory cytokines in local tumors and lung metastases
On days 12 and 25, mice were euthanized, and the tumor tissues and lungs were removed. Total RNA was isolated from the tumors and lungs, and cDNA was synthesized. mRNA expression of decorin, its target genes (mouse Met and mouse CTNNB1), EMT markers (mouse N-cadherin and mouse vimentin), Th1 cytokines (interleukin [IL]-2, IL-12, and tumor necrosis factor [TNF]-α), and Th2 cytokines (TGF-β and IL-6) were detected by real-time RT-PCR using SYBR Premix Ex Taq (Takara, Shiga, Japan) on a 7500 Fast Real-Time PCR System (Applied Biosystems/Life Technologies, Foster City, CA). The relative expression level was calculated by 2–ΔCT, using mouse β-actin as control. The primers for these genes are shown in Supplementary Table S1.
Systemic inflammation responses after treatments with oncolytic adenoviruses
On days 12, 19, and 25, peripheral blood samples were collected, and sera were obtained. The expression levels of chemokines and cytokines, mouse monocyte chemotactic protein 1 (MCP-1), mouse MCP-3, mouse macrophage inflammatory protein 1α (MIP-1α), mouse MIP-2, mouse chemokine (C-C motif) ligand 5 (RANTES), IL-1β, mouse IL-12p70, mouse IL-18, mouse TNF-α, mouse interferon γ (IFN-γ), and mouse IFN-γ inducible protein 10, were detected by Mouse Th1/Th2 and chemokine panel 1 (20 plex) and analyzed by Multi Sciences (Hangzhou, China).
Immune activation analysis after treatment with oncolytic adenoviruses
On days 12, 19, and 25, heparin anti-coagulated peripheral blood samples were collected. Blood samples were labeled with APC conjugated rat anti-mouse CD3e antibody, PE conjugated rat anti-mouse CD4 antibody, and FITC conjugated rat anti-mouse CD8 antibody. Then, the erythrocytes were lysed by 1 × RBC lysing buffer. The immune phenotypes of T lymphocytes were analyzed by flow cytometry.
Statistical analysis
Data are presented as mean ± standard error of the mean and were analyzed using GraphPad Prism v5 (GraphPad Software, Inc., San Diego, CA). Longitudinal data (tumor growth curve and chemokine and cytokine expression in sera) were analyzed using two-way repeated-measures analysis of variance (ANOVA) followed by Bonferroni post hoc tests. One-way ANOVA followed by Bonferroni post hoc tests were performed to analyze other data. Differences were considered significant at two-sided p < 0.05.
Results
rAd.DCN inhibits tumor growth in immune-competent 4T1 orthotopic xenograft
In an immune-competent 4T1 orthotopic xenograft model, oncolytic adenoviruses rAd.DCN and rAd.Null were delivered i.t. or i.v. (via the tail vein). Two injections of viruses were given: on day 7 and on day 10. The tumor volumes were monitored over time. The results show that significant inhibition of tumor growth was observed after treatment with rAd.DCN (Fig. 1A). Consistent with tumor volumes, tumor weight was also inhibited by rAd.DCN, and no significant differences were observed between i.t. and i.v. delivery. The tumor burden was also measured by whole-body BLI on days 7, 14, and 25. From day 7 to day 14, BLI signal increased two- to threefold in the buffer-treated mice. However, only a slight increase in BLI was detected in the oncolytic adenovirus-treated groups (Fig. 1C and D). Moreover, oncolytic adenoviruses treatment reduced tumor burden on day 25. However, no obvious difference was shown between the two methods of delivery (Fig. 1E). These results suggested that both i.t. and i.v. delivery of oncolytic adenoviruses inhibited tumor growth, and rAd.DCN produced much stronger responses.

Both systemic and intratumoral (i.t.) delivery of rAd.DCN inhibit tumor growth in an immune-competent 4T1 orthotopic xenograft model. Exponentially growing 4T1-luc cells (1 × 106/100 μL) were injected into number 3 and 4 mammary fat pads of syngeneic BALB/c mice. On days 7 and 10, 2.5 × 1010 vp/100 μL of rAd.DCN orAd.Null or 100 μL phosphate-buffered saline (buffer group) was administered either i.t. or intravenously (i.v.; via the tail vein). On days 7, 11, 18, and 25, tumor volumes were measured. Tumor growth curves are shown in
rAd.DCN produces decorin protein to inhibit proliferation and promotes apoptosis of tumor cells
Targeting the tumor cells and producing therapeutic proteins are key for oncolytic virus-mediated gene therapy. This study shows that both i.t. and i.v. delivery of adenovirus effectively homed to the tumor sites and infected tumor cells. The immunohistochemistry results show that the adenoviruses still could be detected on day 25. However, the accumulation of significantly more adenoviruses was observed after i.t. delivery on day 12 (Fig. 2A and B). Moreover, rAd.DCN treatment produced decorin proteins in the tumor tissues. Interestingly, decorin expression in the tumor tissues was much more on day 25 than it was on day 12, suggesting that viral genomic replication increased transcription and the production of decorin (Fig. 2A and B).

Analysis of adenovirus replication, decorin expression, proliferation, and apoptosis of tumor cells after rAd.DCN treatment in vivo. Tumor tissues from days 12 and 25 were collected and processed for histopathological analysis.
Next, the study analyzed if adenoviruses evoked antitumor responses in the tumor tissues. The proliferation of tumor cells was inhibited by oncolytic adenoviruses treatment. However, i.t. delivery produced a much superior response (Fig. 2A). It was also found that oncolytic adenoviruses induced cellular apoptosis and cell death in the tumor tissues on day 25, by both i.t. and i.v. delivered viruses (Fig. 2B). The results suggest that oncolytic adenoviruses injected i.v. could home to the tumor tissues and inhibit tumor growth via regulating cellular apoptosis and proliferation.
rAd.DCN produces decorin in the tumor tissues and downregulates its target genes and EMT-related molecules
Decorin can not only inhibit TGF-β signaling, but also target other tumor metastasis-related signaling, such as Met and Wnt/β-Catenin. This study shows that the oncolytic adenovirus rAd.DCN expressed decorin effectively in the tumor tissues at mRNA level on day 12. Importantly, intratumoral delivery of rAd.DCN produced more decorin. Interestingly, both rAd.DCN and rAd.Null downregulated decorin target genes Met and CTNNB1 through i.t. delivery. However, only rAd.DCN significantly decreased the expression of Met and CTNNB1 after i.v. delivery (Fig. 3A). It was hypothesized that downregulation of Met and CTNNB1 following i.t. delivery of oncolytic adenoviruses might contribute to rapid destruction of the tumor tissues. Therefore, it was concluded that decorin protein played a pivotal role in downregulating Met and CTNNB1 expression in tumors. It was found that the decorin mRNA expression in the tumor tissues after rAd.DCN treatment could still be detected on day 25. Importantly, the expression level was nearly equal in i.t. and i.v. deliveries. Although i.t. delivery of rAd.Null slightly decreased decorin target genes, rAd.DCN significantly decreased Met and CTNNB1 expression via both i.t. and i.v. delivery (Fig. 3B). Moreover, both oncolytic adenovirus treatments also downregulated the EMT markers N-cadherin and vimentin. However, rAd.DCN inhibited the EMT of tumor cells much more effectively (Fig. 3C). These results suggest that oncolytic adenovirus-mediated decorin expression reduced tumor metastatic signaling and inhibited EMT of the tumor cells.
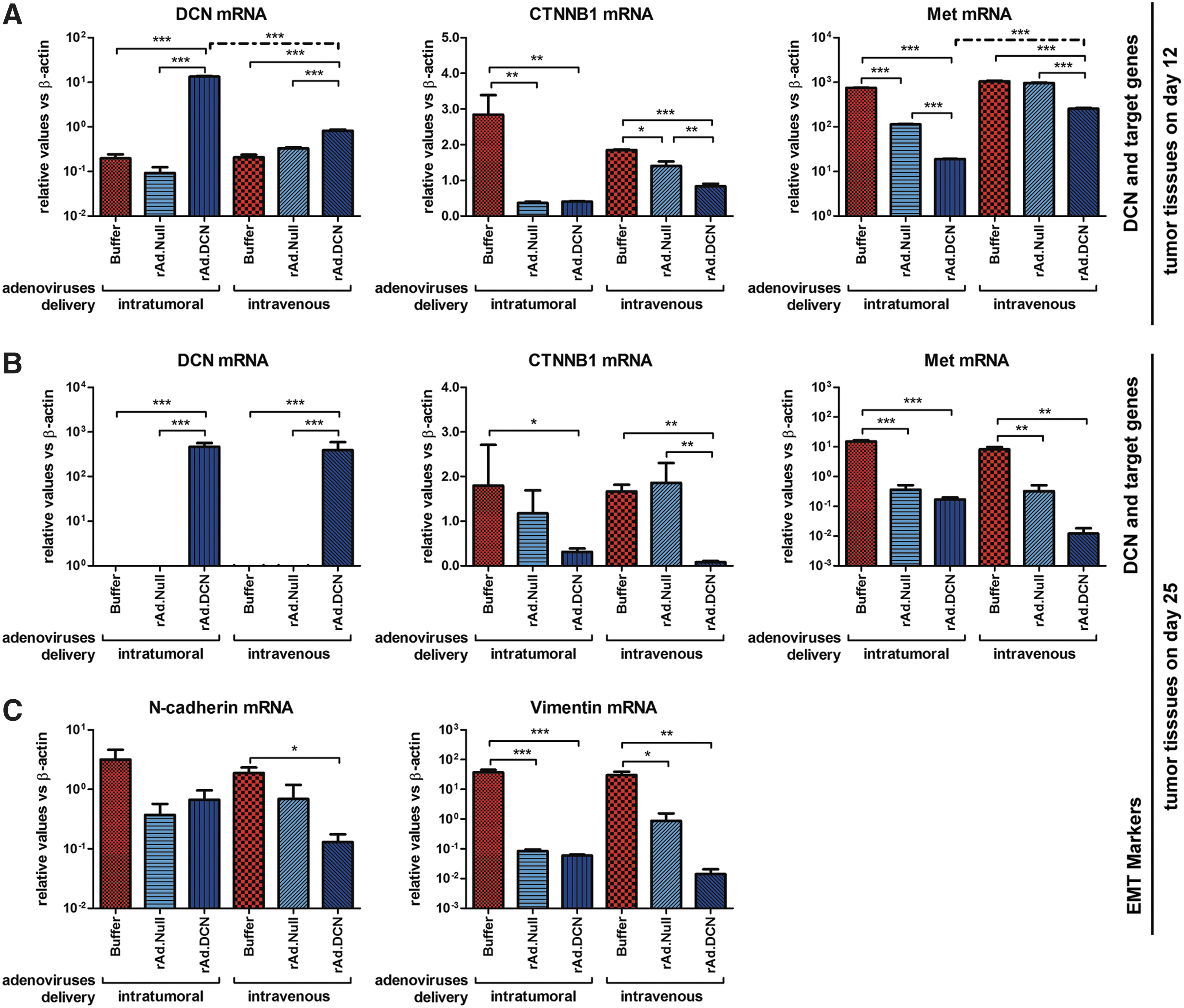
rAd.DCN treatment produces decorin that regulates the expression of decorin target genes and epithelial–mesenchymal transition markers in 4T1 tumors.
rAd.DCN prevents lung metastasis in 4T1 orthotopic xenograft
Since in a 4T1 orthotopic xenograft model spontaneous lung metastasis can be detected, this study evaluated the inhibitory effects of rAd.DCN on lung metastasis. All the mice (6/6) that were analyzed in the buffer group developed lung metastasis 25 days after 4T1 cells were transplanted. However, oncolytic adenovirus treatment inhibited lung metastasis (Fig. 4A and B). Compared to rAd.Null, rAd.DCN treatment increased lung metastasis–free mice (2/6 in the i.t. delivery group; 4/6 in the i.v. delivery group) and reduced the number of metastatic foci. Interestingly, i.v. delivery was much more effective than i.t. delivery (Fig. 4B), which could be due to the increased adenovirus accumulation and decorin expression levels.
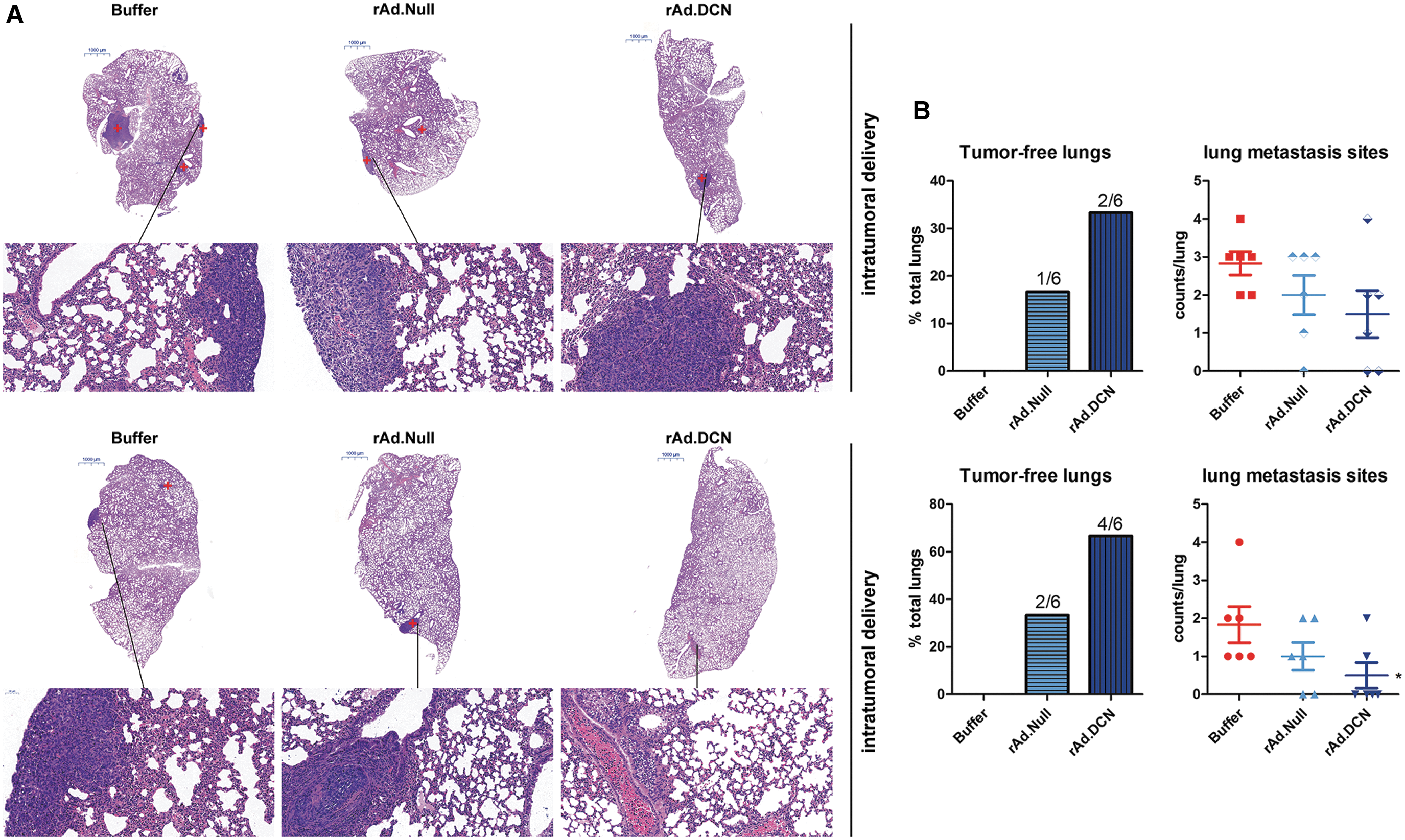
rAd.DCN inhibits lung metastasis from orthotopic tumors following i.t. and i.v. delivery. On day 25, mice were euthanized, and tumors and lungs were removed. The lungs were processed for histopathological assays. The tumor metastatic sites in the lungs were also observed by hematoxylin and eosin staining, and the representative images of the lung sections are shown in

rAd.DCN homes to the lungs and produces decorin that can regulate the expression of target genes and inflammatory responses
On day 12, the distribution of adenoviruses in the lungs was analyzed. It was found that adenoviruses could home to the lungs following both i.t. and i.v. deliveries. However, much more accumulation of adenoviruses was detected in the lungs after i.v. delivery (Fig. 4C). On day 25, rAd.DCN still expressed decorin mRNA in the lungs. However, i.v. delivery produced much more decorin. Moreover, rAd.DCN treatment downregulated the expression of CTNNB1 but not of the Met gene (Fig. 4D).
Decorin is a natural inhibitor of TGF-β signaling, which plays pivotal roles in regulating antitumor responses in the tumor microenvironment. Therefore, the expression of various cytokines was analyzed in lung tissue after treatment with oncolytic adenoviruses. Both oncolytic adenovirus treatments increased the expression of Th1 cytokines, while they reduced Th2 cytokine expression (Fig. 4E). Moreover, i.v. delivery of rAd.DCN increased Th1 cytokine expression, including IL-2, IL-12, and TNF-α, more effectively than i.t. delivery (Fig. 5C). Therefore, it was speculated that rAd.DCN could inhibit tumor metastasis by regulating the tumor microenvironment.

rAd.DCN induces significant inflammatory responses in mouse model of 4T1 orthotopic xenograft. On days 12, 19, and 25, peripheral blood was collected from the mice with various treatments, and sera were obtained. The concentration of chemokines, including mouse monocyte chemotactic protein 1, 3 (MCP-1/3), macrophage inflammatory protein 1α/2 (MIP-1α/2), and RANTES (also named chemokine [C-C motif]) ligand 5
rAd.DCN inhibits tumor growth and metastasis via regulating systemic inflammatory responses
Systemic inflammatory responses play important roles in antitumor responses. It was found that oncolytic adenovirus treatment induced expression of various chemokines, such as MCP-1, MCP-3, RANTES, MIP-1α, and MIP-2. Most of the chemokines were induced at the early stage. However, the upregulation of MCP-1 and MIP-1 could still be observed on day 25 after i.t. delivery of oncolytic adenoviruses. It was speculated that i.t. delivery of oncolytic adenoviruses could lyse tumor cells directly and then release chemokines into the peripheral blood to attract inflammatory cells. Moreover, rAd.DCN treatment produced more chemokines than rAd.Null, although no obvious differences were detected (Fig. 5A).
As described above, oncolytic adenoviruses could upregulate the expression of Th1 cytokines in the lungs. Here, it was demonstrated that both i.t. and i.v. delivery of oncolytic adenoviruses increased the systemic level of Th1 cytokines, such as IL-12, IL-18, TNF-α, and IFN-γ. Moreover, most of the upregulation was detected at the early stage of treatments, 2 days after the last injection of oncolytic adenoviruses (Fig. 5B). These results suggest that oncolytic adenovirus itself could induce Th1 cytokine expression, which might be benefit for evoking strong antitumor responses. Importantly, rAd.DCN increased the expression of Th1 cytokines much more than rAd.Null.
rAd.DCN increased CD8+ T lymphocytes to enhance antitumor responses
It has been reported that the activation of cytotoxic CD8+ T lymphocytes (CTLs) is pivotal to eliminate tumor cells in vivo. This study shows that treatment with oncolytic adenoviruses increased the percentage of CTLs while it downregulated that of CD4+ T lymphocytes. No obvious difference could be detected between i.t. delivery and i.v. delivery on days 12 and 19 (Fig. 6A and B). However, the upregulation of CTLs lasted for a longer period, and a significant increase could be detected, even on day 25, in the i.v. delivery group (Fig. 6C). These results suggest that i.v. delivery of oncolytic adenoviruses promotes the activation of antitumor immune responses more effectively than i.t. delivery.
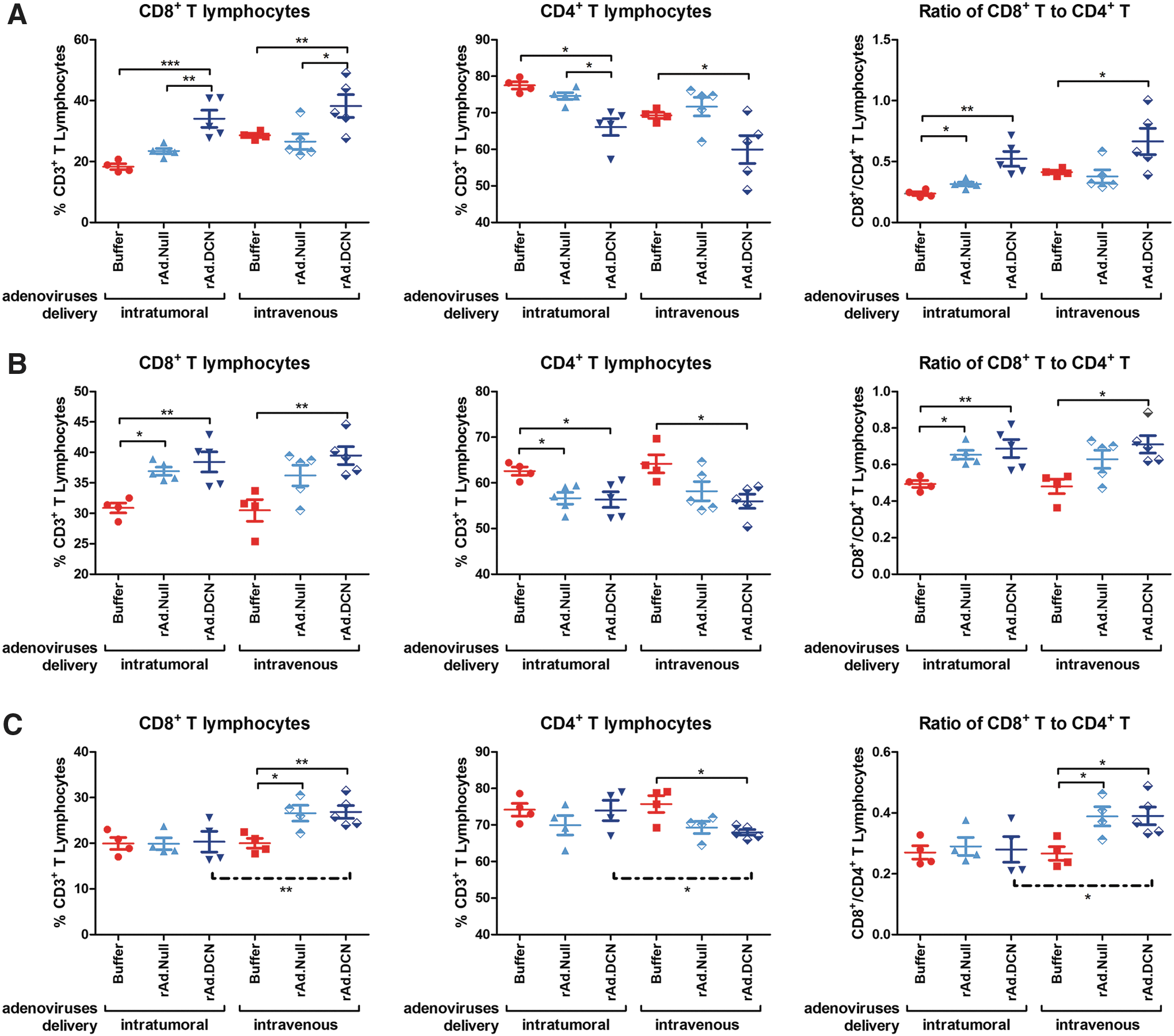
rAd.DCN treatment regulates subtypes of T lymphocytes in the peripheral blood in a mouse model of 4T1 orthotopic xenograft. On days 12, 19, and 25, heparin anti-coagulated peripheral blood samples were collected from five mice from each group. Then, the samples were labeled with APC conjugated rat anti-mouse CD3e antibody, PE conjugated rat anti-mouse CD4 antibody, and FITC conjugated rat anti-mouse CD8 antibody. The immune phenotypes of peripheral blood cells were analyzed by flow cytometry. The statistical analysis of samples on days 12, 19, and 25 are presented in
Discussion
Oncolytic viruses can not only lyse tumor cells via viral replication but can also release a large amount of tumor antigens to activate antitumor immune responses. 18 Therefore, oncolytic viruses have emerged as promising vectors for immune gene therapy. 19 –23 In this study, TERTp controlled oncolytic adenovirus was armed with decorin, rAd.DCN, and its therapeutic responses were evaluated in an immune-competent breast cancer orthotopic model. The results indicate that: (1) both i.t. and i.v. delivery of rAd.DCN can inhibit tumor growth and metastasis to the lung; (2) i.v. delivery is more effective in preventing breast cancer lung metastasis; and (3) rAd.DCN inhibits tumor growth and metastasis via regulating metastasis-related target genes, modulating inflammatory responses, and activating antitumor immune responses.
It is well known that TGF-β plays dual roles in cancer initiation, development, and progression. In advanced cancers, aberrant TGF-β signaling can promote tumor growth, invasion, and metastases via multiple mechanisms. 24 –27 Several TGF-β inhibitors have been developed to block TGF-β signaling in advanced and metastatic cancers, which could significantly inhibit the progression of tumors in both preclinical animal models and clinical trials. 21,28,29 An oncolytic adenovirus expressing sTGFβRIIFc (Ad.sT) to target TGF-β signaling has been developed previously. In a bone metastasis model of prostate and breast cancers, Ad.sT inhibited and delayed the progression of tumor bone metastasis. 8,9,30 Moreover, sTGFβRIIFc downregulated metastasis-related target genes of TGF-β, such as IL-11, chemokine (C-X-C motif) receptor 4 (CXCR4), parathyroid hormone-related peptide, and CTGF (connective tissue growth factor), in various tumor cells (unpublished data). Decorin has been suggested to be a natural inhibitor of TGF-β signaling. Therefore, decorin might be a potential target for treating advanced and metastatic cancers. 12
Downregulation of decorin has been demonstrated in tumor tissues of various advanced cancer patients, and restoring decorin expression has emerged as a potential therapeutic strategy. 5,31 –34 To the best of the authors' knowledge, this is the first study to show that oncolytic adenovirus expressing decorin can inhibit bone metastasis of breast and prostate cancers in immune-deficient xenograft mouse models. 6 Recently, it was shown that the new TERTp-promoted decorin-expressing rAd.DCN can inhibit lung metastasis of colon cancer cells in an immune-competent syngeneic mouse model. 5 This study shows that oncolytic adenovirus expressing decorin, rAd.DCN, significantly inhibits tumor growth and lung metastasis in an immune-competent breast cancer orthotopic model. Moreover, it has been demonstrated that decorin inhibits tumor growth and invasion by multiple mechanisms, such as promoting cellular and mitochondrial autophagy, and blocking pivotal cellular signaling, such as Met, Wnt/β-catenin, and vascular endothelial growth factor (VEGF) signaling. 6,13,14,35 Decorin restoration in tumor cells in vitro downregulates expression of Met, CTNNB1, VEGFA, and TGF-β. 6,13 This study confirms that decorin decreases Met and CTNNB1 expression in both tumor and lung tissues of a 4T1 orthotopic xenograft. It was hypothesized that rAd.DCN could infect tumor cells in orthotopic sites as well as metastatic sites, and express decorin protein. rAd.DCN then lyses the tumor cells and downregulates target genes of decorin to inhibit tumor growth and metastasis.
It has been reported that TGF-β is an important molecule in regulating the tumor microenvironment. High TGF-β levels can suppress the maturation of effector cells, such as T helper cells, dendritic cells, and natural killer cells, as well as induce M2 polarization of macrophage cells. Moreover, TGF-β can also regulate the expression of cytokines and chemokines to regulate antitumor inflammatory responses and to drive Th1/Th2 balance toward the Th2 immune phenotype. 36 –38 Preclinical studies suggest that both anti-TGF-β antibodies and small molecules targeting TGF-β signaling can modulate the tumor microenvironment, such as restoring the cytotoxicity of CTLs, enhancing the antigen presentation ability of dendritic cells (DCs), and decreasing the percentage of CD4+CD25+ Tregs. 39 –42 In an immune-deficient MB-MDA-231 model, the therapeutic effects of Ad.DCN in preventing breast cancer bone metastasis have been confirmed. 6 However, in immune-deficient mice, it is not possible to evaluate the role of decorin in regulating the immune functions associated with the alteration of the tumor microenvironment. This study found that after treatment with rAd.DCN, significant systemic inflammatory responses, significant upregulation of Th1 cytokines and downregulation of Th2 cytokines, and activation of CTLs could be detected in an immune-competent 4T1 orthotopic model. The results are consistent with previous reported work, in which it was shown that oncolytic adenovirus-mediated decorin expression downregulated TGF-β expression in the tumor environment and promoted expansion and maturation of DCs in a colon cancer (CT26) xenograft model. 5
This study also compared the therapeutic effects between i.t. and i.v. delivery of rAd.DCN on tumor growth and lung metastasis. It shows that i.t. delivery can infect tumor cells and induce cytotoxicity in tumor tissues much more effectively than i.v. delivery. It has been demonstrated that i.v. delivery of serotype 5 adenovirus can result in the accumulation of viruses in the lung and liver. 7,10,43 –45 This study shows the accumulation of adenoviruses in the liver, lungs, and tumors. The results suggest that i.v. delivery of oncolytic adenoviruses can effectively infect tumor cells, lung epithelial cells, and hepatocytes (data of adenovirus accumulation in the liver not shown). It has been demonstrated that oncolytic adenovirus can modulate antitumor immune responses by releasing tumor-specific antigens and regulating immunological suppression in the tumor microenvironment. 46,47 In this study, oncolytic adenovirus-mediated decorin expression in the lungs downregulated decorin target genes and modulated antitumor immune responses to prevent tumor lung metastasis. Although adenoviruses could also be detected in the lungs after i.t. delivery, less decorin expression and weaker inhibitory effects on lung metastasis were observed. These results suggest that i.v. delivery of oncolytic adenoviruses will be an alternative approach for treating tumor lung metastasis. Moreover, biomaterials have been widely used in gene therapy to increase gene transfer efficiency, as well as to reduce toxicity to normal tissues. 48 –51 Therefore, combining biomaterials might be a potential approach to enhance the antitumor efficiency of oncolytic viruses.
The model used in this study to explain the antitumor responses of oncolytic viruses expressing decorin can be summarized as follows. Intratumoral and i.v. delivery of rAd.DCN infects tumor cells at orthotopic and metastatic sites, as well as epithelial cells in the lungs. Then, rAd.DCN lyses tumor cells to release tumor antigens, which can activate antitumor immune responses. In tumor tissues and lungs, rAd.DCN produces decorin protein to inhibit CTNNB1 and Met expression, inhibits EMT of tumor cells, increases Th1 cytokines, inhibits Th2 cytokine expression, evokes a strong anti-tumor inflammatory response, and activates CTLs. An important observation made here is that i.v. delivery of rAd.DCN is much more effective in preventing tumor lung metastasis compared to i.t. delivery (Supplementary Fig. S1). Therefore, systemic delivery of rAd.DCN might be a promising strategy for breast cancer lung metastasis.
Footnotes
Acknowledgments
This work was supported by Chinese National Natural Science Foundation of China (no. 81402558&81472396), National High Technology Research and Development Program of China (863 Program; no.SS2014AA020515), and Nova Program of Beijing (Z171100001117118). The work at NorthShore was partly funded by a cancer gene therapy grant (P.S.).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.