
Abstract
Long noncoding RNAs (LncRNA), a class of transcripts with lengths >200 nt, play a master role in the regulation of cancer pathogenesis. Recently, the CRISPR-Cas9 system has been explored as a revolutionary genome editing tool for molecular biology. Growing evidence shows that LncRNAs can be targeted by the CRISPR-Cas9 system used for evaluating its function. Thus, the CRISPR-Cas9 systems provide a novel gene-editing strategy for the modification of LncRNA expression. This review summarizes current knowledge of the functions and underlying mechanisms of LncRNA by CRISPR-Cas9. Emerging strategies for non-viral/viral delivery of CRISPR-Cas9 in a clinical context are also discussed.
Introduction
L
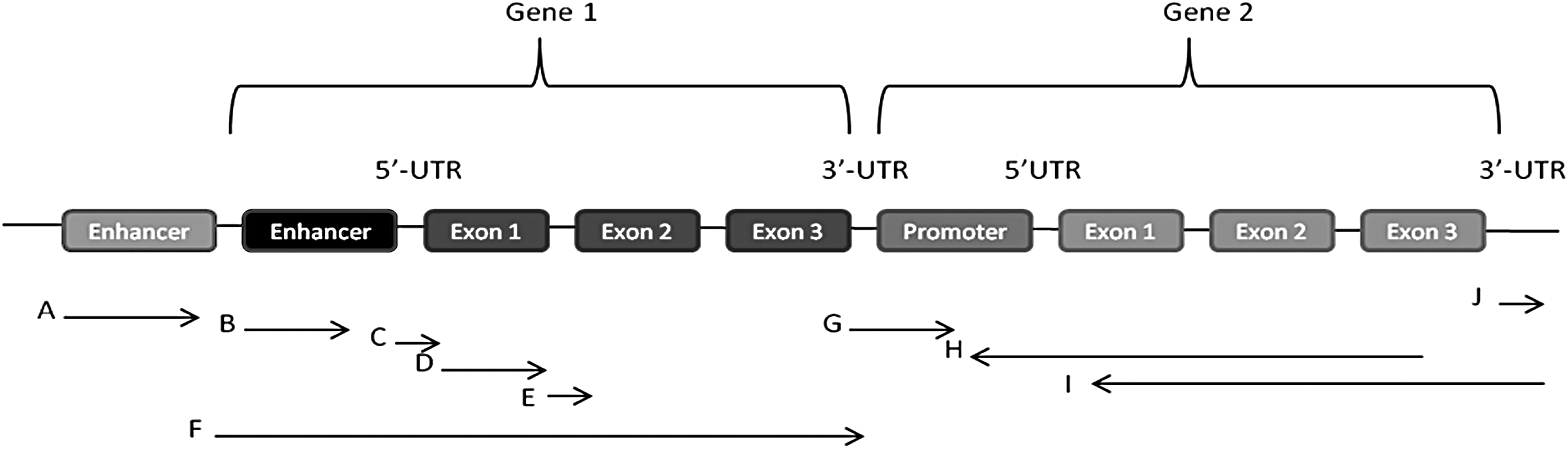
The main origins of long noncoding RNAs (LncRNAs). LncRNAs originate from enhancer sequences (A), promoter regions (B), 5′-UTRs (C), exons (D), introns (E), intragenic regions (F), intergenic sequences (G), antisense sequences (H and I), and 3′-UTRs (J).
Genetic modification is an essential approach to study the function of genes. The gene-editing technologies such as zinc finger nucleases, transcription activator-like effector nucleases, and the clustered regularly-interspaced, short palindromic repeats (CRISPR)-associated protein 9 (Cas9) system have accelerated the development of human genome manipulation greatly. 8
Three types (I–III) of CRISPR system were first discovered in bacteria and archaea where they served as an immune defense system conferring resistance to foreign viruses (e.g., phages) and other genetic elements. 9,10 The Streptococcus pyogenes type II CRISPR locus includes the Cas9 nuclease enzyme and a Cas9-associated single-guide RNA (sgRNA). 11,12 Theoretically, any DNA sequence followed by a PAM can be recognized and cleaved by the sgRNA:Cas9 ribonucleoprotein (sgRNP) complex to achieve a desired editing outcome. 11,12 The easy design of the CRISPR-Cas9 system, coupled with its high efficiency targeting and low off-target mutation frequency, has rapidly made it the most commonly used approach for gene editing. 13,14 Thus, the CRISPR-Cas9 has been broadly applied as a genetic-engineering tool in studies of human disease (Fig. 2). 15,16
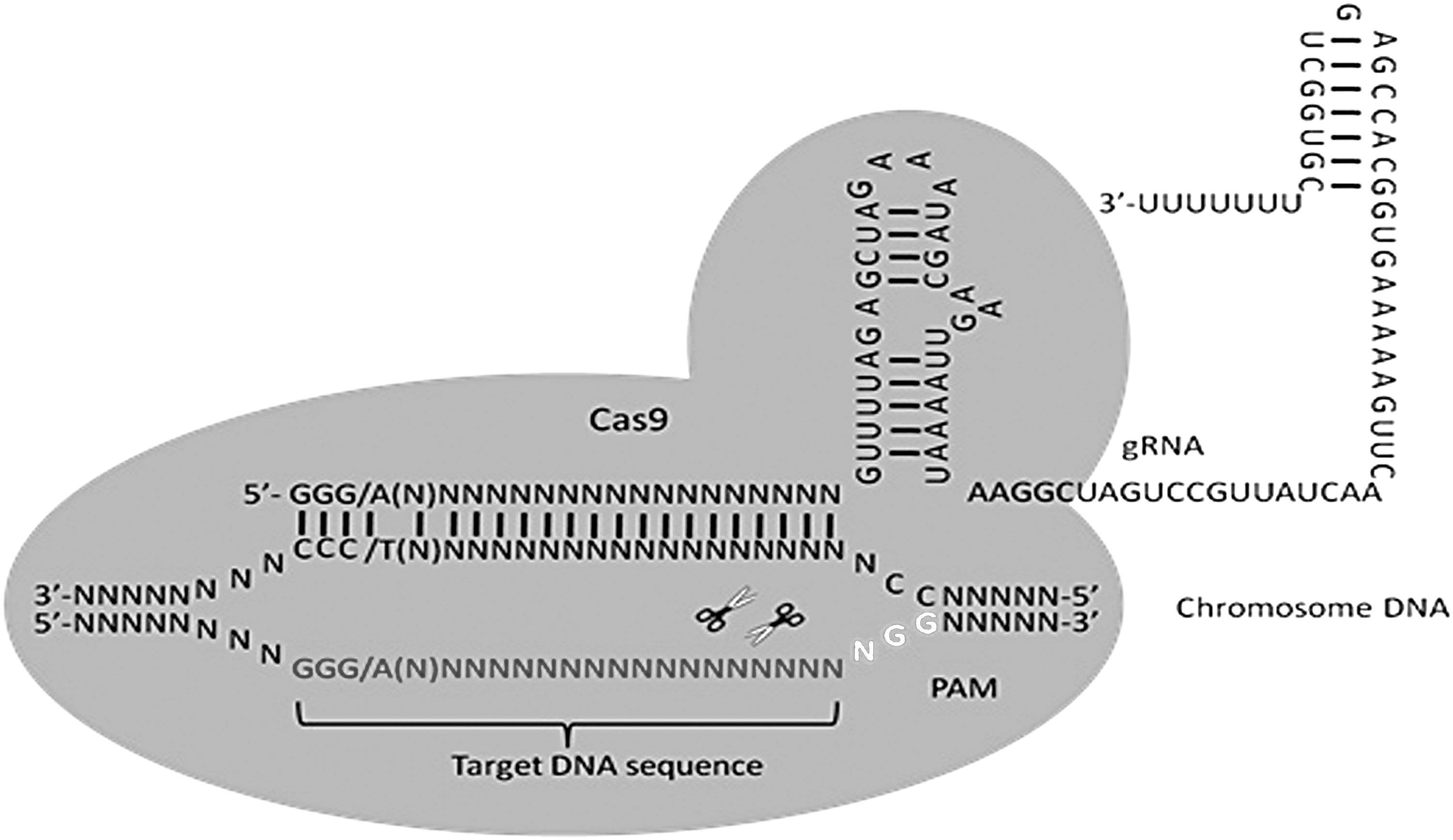
The mechanism of the CRISPR-Cas9 system.
Here, the current status and recent progress of LncRNA-targeted therapeutics through CRISPR-Cas9 are critically examined, along with alternative emerging strategies specifically to combine different delivery platforms and cell-fate engineering for the elucidation of LncRNAs function and LncRNA-based therapeutic intervention.
CRISPR-Cas9: A Strategy For Interrogating Lncrna Gene Function
The discovery of CRISPR-Cas9 has provided a tool to solve problems in the elucidation of LncRNA function. 17 Foremost, this is the discrete and permanent nature of CRISPR-based lncRNA knockout (Fig. 3). However, straightforward insertions/deletions ceased by a single double-strand break are unlikely to cause functional ablation of a noncoding gene. Therefore, one has to consider a more comprehensive approach with respect to LncRNA function. One strategy that ensures complete ablation would be to delete the entire genomic region associated with a LncRNA. 18 This approach is highly effective and has been expanded to create highly informative genome-wide scales of LncRNA depletion screens. 19 However, deletion of the genomic DNA can confound the issue of whether the phenotypic effect is due to loss of the noncoding transcript or potential regulatory sequences of the genomic region itself. 20,21 This was exemplified by the systematic CRISPR-based deletion and insertion of the LncRNA Haunt. 22 In this report, the authors demonstrated discrete and opposing roles for the LncRNA transcript Haunt and its genomic locus in regulating the HOXA gene cluster during embryonic stem cell differentiation. Strikingly, interruption of Haunt transcription via insertion of a transcriptional stop signal increased HoxA activation. Furthermore, insertion of the Haunt cDNA within genomic knockouts of the same region failed to restore HoxA activation, ultimately demonstrating that the Haunt genomic region has an enhancing regulatory function, while the noncoding transcript itself appears to act as a repressor. As outlined above, introducing a transcriptional stop signal can be highly effective in ablating the LncRNA transcript without significantly affecting the role of the DNA sequence itself, as shown by others. 23 An alternative approach is to target the promoter region of the LncRNA. By deleting the region surrounding the promoter of LncRNA-CSR, Basu et al. demonstrated that the absence of the transcript itself (despite an intact genomic region) disrupted long-range chromatin contacts >2.6 Mb away, leading to dysregulation of isotype switching in B cells. 24 A further advantage of targeting the promoter is the small size that needs to be deleted, increasing the efficiency of editing. As such, paired guide-based CRISPR deletions of LncRNA promoters have been adapted in large-scale studies for screening the effects of LncRNAs. 25
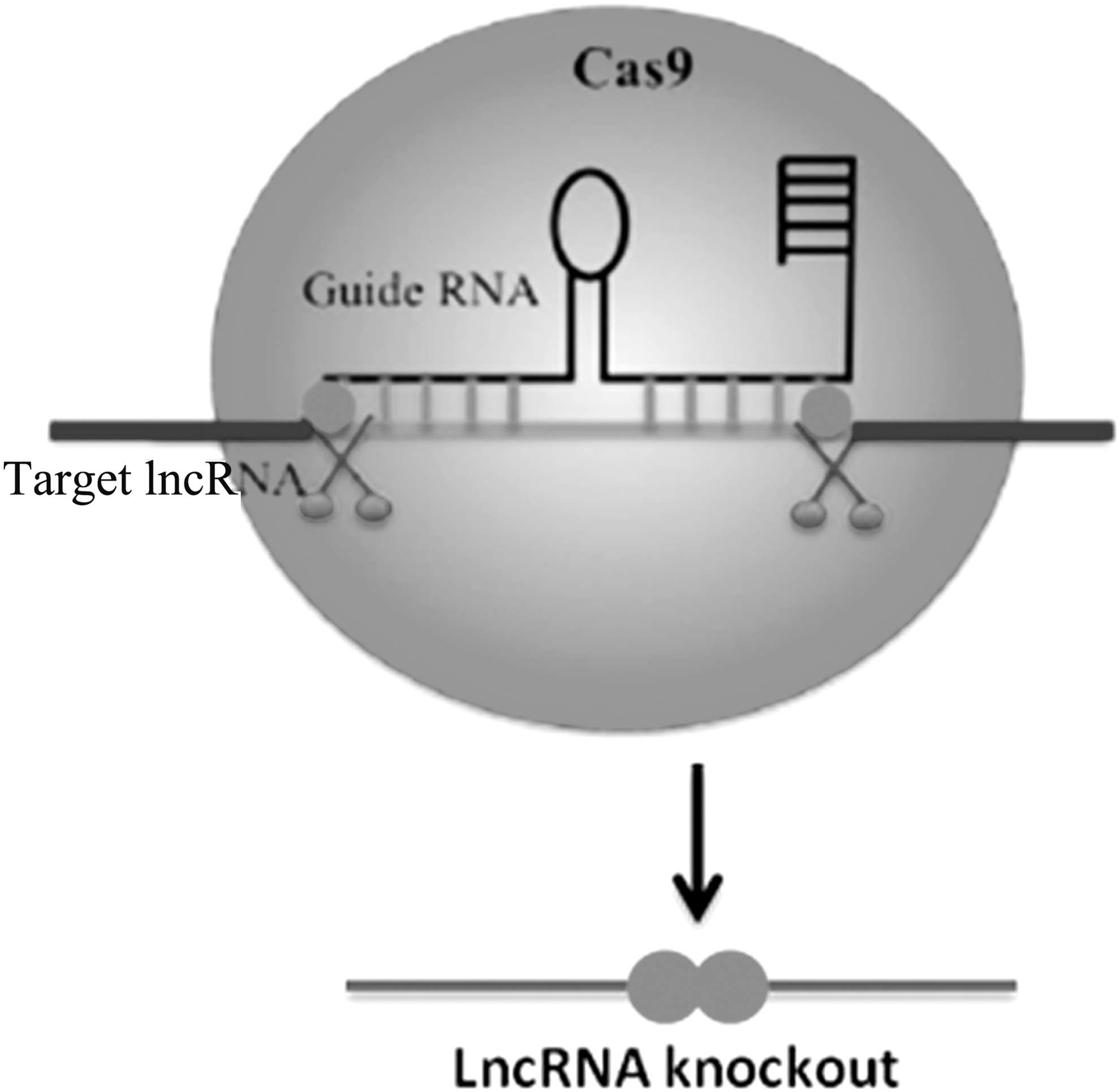
The CRISPR-Cas9 genome editing method utilizes a guide RNA to drive the Cas9 endonuclease, which cleaves a specific sequence in the target LncRNAs.
These genome-engineering approaches can be further enhanced with the use of the iCRISPR system. 26 By integrating an inducible Cas9 construct into the AAVS1 safe harbor locus of induced pluripotent stem cells (iPSCs), LncRNAs could be targeted by the simple addition of RNA guides. The power of this system lies within the ability to differentiate these iPSCs subsequently to specific cell types that may be relevant to the LncRNA function. Furthermore, LncRNAs responsible for lineage specification can also be identified using this system through the systematic ablation of LncRNAs and observing the resultant cell fates.
CRISPR-Cas9 System In Cancer Research
Breast carcinoma
In a recent study, the roles of LncRoR in breast cancer were investigated. 27 Peng et al. manipulated LncRoR expression in breast carcinoma cell lines by means of CRISPR-Cas9 technology. Using this strategy, the authors demonstrated that LncRoR promoted estrogen growth and activated the MAPK/ERK pathway of breast carcinoma. This report is a representative example of the manner in which CRISPR-Cas9 technology could be applied for LncRNA loss-of-function studies and provides evidence that LncRoR could be a potential target in estrogen-receptor-positive (ER+) breast carcinoma.
Another representative study was reported by Singh et al., 28 who showed that LncBC200 is upregulated in breast cancer. Among breast tumor specimens, the level of LncBC200 is higher in ER+ tumors. To know the significance of ER-regulated LncBC200 expression in breast cancer, they knocked out LncBC200 by CRISPR-Cas9 technology to suppress tumor cell growth in vitro and in vivo. The results demonstrated that LncBC200 plays an important role in breast cancer.
In 2017, Pratirodh et al. reported that LncAK023948 is a positive regulator in the AKT pathway that is dysregulated for many cancers. The results reported that AK023948 is upregulated in breast cancers, and it was knocked out using CRISPR-Cas9, leading to reduced tumor progression. 29 The same results were shown by Ho et al., who designed a dual gRNA targeting AK023948. 30
Bladder cell carcinoma
Recently, the LncRNA-UCA1 employing the CRISPR-Cas9 system was targeted in bladder carcinoma cell lines (5637 cell and T24 cell). 31 This report suggested that knockdown efficiency of the sgRNA targeting the promoter region of LncRNA-UCA1 was >80%, whereas the knockdown efficiency for LncRNA-UCA1-exon1 was between 85% and 90%. Furthermore, CRISPR-Cas9-UCA1-(exon1 + promoter) was co-transfected into bladder cancer cells, and synergistic inhibition on UCA1 expression was observed. Upon deleting Lnc-UCA1, the authors suggested that its downregulation resulted in decreased tumorigenesis. Therefore, the application of CRISPR-Cas9 to study specific LncRNAs for this particular carcinoma was demonstrated.
Prostate cancer
Some studies have revealed the involvement of LncRNA in the pathogenesis and progression of various human malignancies, including prostate cancer. 32,33 For example, LncRNAs have been implicated in different cellular processes in prostate carcinoma. Beaver et al. 34 suggested that sulforaphane (SFN)-mediated alterations in LncRNA expression correlated with genes that regulate cell cycle, signal transduction, and metabolism, such as LINC01116. When LINC01116 was disrupted by CRISPR-Cas9, a decrease in the ability of the prostate cancer cells was found, supporting an oncogenic function for LINC01116. They identified a novel isoform of LINC01116 and suggested that LINC01116 could interact with target genes. Data reveal that SFN can influence the expression of functionally important LncRNAs.
Colorectal cancer
There has been an interest in analyzing the role of LncRNA as oncogene functional components in colorectal cancer (CRC). 35 In a recent study, it was reported that inhibition of proliferation in CRC cells is mediated through the loss of LncRNA CCAT2 expression. In this study, Yu et al. used the CRISPR-Cas gene-editing approach to targeting LncCCAT2. Knockout of LncCCAT2 increased the expression of miR-145 and decreased the expression of miR-21 in colon cancer cells (HCT-116), impaired proliferation and differentiation. This study suggested that LncCCAT2 regulated CRC proliferation and differentiation.
Another representative study was reported by Ho et al., who designed a dual gRNA targeting lncRNA-21A. Co-transfection of dual vectors of donor and LncRNA-21A into CRC cells (HCT-116) resulted in a 16.3-fold increase in colony number. 30
Gastric cancer
Gastric cancer (GC) is one of the most common cancers in the world. Liu et al. 36 manipulated LncPANDAR expression in human GC cell lines by means of CRISPR-Cas9 targeting the sequences of Lnc-PANDAR. Using this strategy, they demonstrated that LncPANDAR knockout suppressed malignant features in GC cells and inhibited tumor growth in vivo. This study is a representative example of how CRISPR-Cas9 technology can be used for LncRNA studies and provides evidence that LncPANDAR may be a potential therapeutic target for GC.
Application Of CRISPR-Cas9 Technology For Lncrna Therapeutics: The Delivery-Related Concerns
Gain- and loss-of-function approaches have been utilized to study the roles of LncRNAs. Due to the potential ability of CRISPR-Cas9, the roles of LncRNAs in therapeutic approaches have become an area of intense research in recent years (Table 1).
LncRNA editing in cancer
LncRNA, long noncoding RNAs.
For therapeutic genome editing, it is desirable to restrict the expression of CRISPR components, in particular tissues, to avoid potential off-target effects. As a result, it is possible to limit the expression of the Cas proteins in a specific tissue using tissue-specific promoters. The general perturbation of cellular or tissue homeostasis by the delivery process itself can also trigger unwanted responses. 37 Such nonspecific effects associated with the delivery process can likely be overcome by minimizing lipid concentration. Nevertheless, the natural function of Cas9 is to act as a nuclease by creating DNA double-strand breaks (DSB) via RuvC and HNH endonuclease domains, each of which cleaves one strand of the target DNA. 38 An alternative to limit off-target activities of CRISPR-Cas9 further is the use of a modified Cas9 enzyme that possesses only one active catalytic domain (designated “Cas9-nickase” or Cas9n) for binding sgRNA specificity, but is only capable of cutting one of the DNA strands, resulting in a “nick” or single-stranded break instead of a DSB. 39,40 Thus, the “dual nickase” or “double nick” CRISPR system is a method that could be intended to decrease off-target cleavage frequency for LncRNA editing by using a single Cas9 nickase and two different sgRNAs. In addition to minimizing off-target effects, sgRNA length is also optimized. For example, it has been reported that truncated sgRNAs with <20 base homology displayed fewer off-target effects. 41 Furthermore, two new Cas9 components have been generated to display low off-target activity at nearly undetectable levels due to rationally designed mutations: (1) enhanced Streptococcus pyogenes Cas9 (eSpCas9) with helicase activity reduced without altering on-target editing efficiency, 42 and (2) a high-fidelity variant of SpCas9, which contains engineered mutations in the SpCas9 residues that normally form hydrogen bonds with DNA. 43 Any of these variants can be used for LncRNAs gene editing.
On the other hand, targeting a single LncRNA region is not sufficient, as some diseases, including cancer, would require more targets of LncRNAs to disrupt the pathological state of the cell associated with them. Thus, different approaches would involve the use of sgRNAs targeting multiple genomic loci for designing multiplex CRISPR/Cas9-based genome-engineering tools. 44 Previously, a system was designed and constructed for expressing and delivering a Cas9 nuclease and two sgRNAs expressed from the independent and different surface antigen-encoding region of hepatitis B virus (HBV). 45 Interestingly, each sgRNA was efficiently expressed, and it was possible to promote multiplex gene editing and inhibit HBV expression and replication in vitro and in vivo, providing a useful methodology that could also be applicable for LncRNA-based biomedical research. Notwithstanding this, employing different promoters or performing several treatments may increase the adverse risk occurring through off-target effects. 46 To overcome these limitations partially, multiple sgRNAs could be expressed from a single promoter simultaneously, and each sgRNA could be flanked by two self-cleaving ribozymes and/or self-cleaving transfer RNA (tRNA) to perform trimming activities for the release of individual and functional sgRNAs 47 against different miRNA sequences. This represents an attractive strategy to avoid co-delivery of multiple sgRNA encoding constructs and to reduce the amounts of CRISPR-Cas9 components and the potential adverse side effects.
However, beyond possible off-target effects, a major challenge of CRISPR-Cas is the in vivo delivery method. Thus, CRISPR-Cas9 technology will require the adoption of appropriate delivery systems in practical applications.
Currently, nucleic acid–based aptamers have been successfully adapted for targeted delivery via specific cell surface receptors in vitro and in vivo. 48 Recently, an aptamerliposome-CRISPR-Cas9 chimera was developed incorporating an aptamer that specifically binds to the prostate membrane antigen on cancer cells. 49 Subsequently, CRISPR-Cas9 delivered by cationic liposomes targeted polo-like kinase 1, which is expressed in tumor cells. It was demonstrated that this aptamer-liposome CRISPR-Cas9 chimera-based approach possessed significant cell-type specificity and gene knockdown effect in vitro and in vivo. To the authors' knowledge, this is the only application of the aptamer-cationic liposome approach to achieve CRISPR-Cas9 delivery. Nevertheless, although targeting tumors using aptamers is an attractive application of CRISPR-Cas9, this approach is still a huge challenge because there are few RNA aptamers adapted for the targeted delivery via specific cell surface receptors. In fact, the isolation of high-affinity aptamer ligands is in itself a difficult task per se. This indicates the need to implement other plasmid DNA-free CRISPR approaches for CRISPR-Cas9 delivery.
Recently, Mout et al. 50 reported a highly efficient editing method based on the co-delivery of Cas9 protein complexed with a sgRNA (Cas9-RNPs complex) for efficient delivery into the cytoplasm, Interestingly, it was demonstrated that the nano-assembly-mediated Cas9-RNP delivery occurred preferably through a cholesterol-dependent membrane fusion-like process, avoiding the endosomal accumulation of both Cas9 and sgRNA. By means of this DNA-free CRISPR approach, the authors achieved up to around 90% delivery efficiency in a range of cell types and gene-editing efficiency of up to 30%.
Cas9 and gRNA complex can be delivered into the cell using the expression vectors like virals. 51 Five major viral vectors—retroviruses, 52 lentiviruses, 53 adenoviruses, 54 adeno-associated virus (AAV), 55 and baculoviruses 56 —have been used for genome editing. Among these, AAVs have provided major advantages for therapeutics, including a very mild immune response and toxicity. Moreover, AAVs remain primarily episomal upon transduction, avoiding random integration of the viral genetic materials into the host genome that can disrupt neighboring gene function and cause insertional mutagenesis.
The therapeutic applicability of CRISPR-Cas9 for LncRNA editing represents other concerns for translational potential. For example, for LncRNAs with an essential role in cellular physiology regulating many different targets, partial loss-of-function studies are important. 57 Currently, technologies have been reported that are based on nuclease-deactivated Cas9, which can be directed near the promoter sequence of a target gene to exert RNA-guided transcription regulation without permanently modifying the genome. Hence, by activation (CRISPRa) or repression (CRISPRi), transient or stable control of LncRNA expression without altering the genomic sequence would be intended. For example, LncRNA-transcription regulation can be modulated through sgRNA-guided DNA-binding proteins, which can be fused with an epigenetic modifier (e.g., methylase or acetylase). Furthermore, several approaches have been attempted to design inducible systems that use minimal elements to achieve Cas9 expression. 58,59 Similar to miRNA, Hirosawa et al. 60 developed a miRNA-responsive CRISPR-Cas9 system called miR-Cas9 switches, which efficiently responded to miRNA expression in cells, and post-transcriptionally attenuated Cas9 activity only in the presence of each miRNA in these target cells. By means of this strategy, using LncRNA complementary sequences in the 50-UTR of LncRNA encoding Cas9, its genome-editing activity may be modulated through endogenous LncRNA signatures in human cells.
Conclusion and Future Perspectives
Despite the ethical controversies regarding non-research applications of CRISPR-Cas technology, it is clear that CRISPR-Cas represents a cutting-edge tool suitable for basic research and future therapeutic modalities. Given the flexibility in the design of sgRNA, it is now possible to target a wide range of DNA sequences containing PAM. In addition, developments in nanotechnology and materials for nonviral delivery systems, including liposomes, polymers, nanoparticles, and cell-penetrating peptides, have increased the alternatives available for the delivery of Cas proteins and sgRNAs. 61 Therefore, research efforts must be encouraged to investigate the potential application of CRISPR technologies as a traditional LncRNA editing strategy for the elucidation of LncRNA function and LncRNA-based therapeutic intervention, with an emphasis on combining one or more delivery strategies that might increase the success rate of delivery of CRISPR-Cas components in cancer cells.
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (grant nos. 81602295 to S.Z.).
Author Disclosure
The authors have no conflict of interest.