
Abstract
High-risk human papillomavirus (HPV) infection is a causal factor in oropharyngeal and gynecological malignancies, and development of HPV-targeted immunotherapy could be used to treat patients with these cancers. T cell–mediated adoptive immunotherapy targeting E6 and E7, two HPV16 proteins consistently expressed in tumor cells, appears to be both attractive and safe. However, isolation of HPV-specific T cells is difficult owing to the low frequency of these cell precursors in the peripheral blood. In addition, HPV-positive cancer cells often down-regulate major histocompatibility complex (MHC) class I expression ex vivo, limiting the efficacy of MHC class I-restricted approaches. Of particular interest is that both CD4 and CD8 T cells can mediate the responses. Given that CD4 T cells play a critical role in coordinating effective antitumor responses, the generation of a T helper response in patients with HPV16-associated malignancies would unleash the ultimate potential of immunotherapy. In this view, T-cell receptor (TCR) gene transfer could be a relevant strategy to generate HPV16-E7-specific and MHC class II–restricted T cells in sufficient numbers. An HPV16-E7/HLA-DRB1*04 TCR has been isolated from a cancer patient with complete response, and retroviral particles encoding this TCR have been produced. The transgenic TCR is highly expressed in transduced T cells, with a functional inducible caspase-9 suicide gene safety cassette. TCR transgenic T cells are HPV16-E770-89 specific and HLA-DRB1*04 restricted, as determined by interferon (IFN)-γ secretion. CD8 and CD4 T cells are equivalently transduced and secrete interleukin-2 and IFN-γ when cultured with appropriate targets. We also demonstrate that TCR transgenic T cells recognize the endogenously processed and presented HPV16-E770-89 peptide. In conclusion, our data indicate that the production of MHC class II–restricted HPV16-E7–specific T cells is feasible through TCR gene transfer and could be used for immunotherapy.
Introduction
H
HPV-specific T cells are difficult to isolate due to their low frequency in peripheral blood. Indeed, the detection of cytotoxic T cell (CTL) responses against HPV antigens is challenging in women with natural HPV infections. 9,10 In addition, patients with HPV16-positive genital lesions and cervical cancer display either absent or considerably decreased HPV16-specific CD4 T helper responses. 11,12 However, positive clinical outcomes can occur after adoptive cell transfer of tumor-infiltrating lymphocytes obtained from HPV-positive metastatic cervical cancer patients. 13 Accordingly, an attractive approach for treating patients with HPV-related malignancies is the adoptive transfer of HPV-specific CD4 and CD8 T cells.
The introduction of an engineered TCR into recipient T cells is a promising strategy to generate tumor-specific T cells in high numbers. Recently, both E6- and E7-specific TCRs were used to produce HPV-specific CD8 T cells. 6,14 A recent phase I/II clinical study in HPV16-positive cancers treated with autologous T cells expressing the E6/HLA-A2 TCR was completed, but the results have not been published yet (NCT02280811). In contrast, characterization of major histocompatibility complex (MHC) class II–restricted TCR with specificity for tumor-associated antigens has rarely been performed. 15,16 Nonetheless, it has been previously reported that both CD4 and CD8 recipient T cells can be genetically modified with TCRs derived from CD4 T cell clones. 16,17 Hence, the identification and characterization of human MHC class II–restricted TCR with tumor reactivity is crucial for the further development of clinical applications. This need might be especially relevant for HPV-induced lesions because MHC class I surface expression can be impaired owing to HPV immune evasion strategies, while MHC class II molecules are expressed in high-grade cervical cancer. 18,19 We therefore sought to isolate an HLA-DR-restricted HPV16-E7–specific TCR from a cancer patient in complete remission and to characterize transgenic T cells redirected with this TCR targeting HPV-positive tumor cells.
Materials and Methods
Biological material
Peripheral blood was obtained from an HLA-DRB1*04:04 patient (case 1), 20 enrolled in Epitope-HPV01 clinical trial (NCT01845779), conducted according to applicable statutory requirements. This patient was treated for an HPV16-positive metastatic cancer of the anal canal and had achieved a state of complete remission at the time of the sampling. Peripheral blood mononuclear cells (PBMCs) were collected from HLA-DRB1*04 healthy donors at the Etablissement Français du Sang (EFS) as apheresis kit preparations after informed consent and following internal guidelines. The PG13 packaging cell line and the human cervical cancer cell lines SiHa and C33A were obtained from American Type Culture Collection bank (Manassas, VA; ATCC CRL-10686, ATCC HTB-35, and ATCC HTB-31, respectively). All lines were cultured according to the manufacturer's instructions. Cell lines were authenticated and periodically checked for mycoplasma contamination. Epstein-Barr virus–transformed B lymphoblastoid cell lines (BLCLs) were generated from healthy donor PBMCs as previously described. 21
Peptides
The Immune Epitope Database and Analysis Resource 22 (IEDB) was used to predict binding of 20mer peptides derived from HPV16-E7 to HLA-DRB1*04 allele. As recommended by the IEDB, peptides with a low percentile rank were considered as good binders. HPV16-E76-25 (PTLHEYMLDLQPETTDLYCY) and HPV16-E770-89 (QSTHVDIRTLEDLLMGTLGI) were selected and purchased from Proteogenix (Schiltigheim, France).
CD4 T cell clone isolation and amplification
PBMCs from the case 1 patient 20 were isolated by centrifugation on Ficoll gradient (Eurobio, Les Ulis, France, CMSMSL01-01) and plated at 2 × 10 6 to 4 × 10 6 cells per well in a 24-well plate in RPMI-1640 (Fisher, Hampton, NH, 11534526) supplemented with 10% human serum (EFS local production) and in the presence of an HPV16-E7 peptide pool, PepTivator HPV16-E7 (Miltenyi Biotec, Bergisch Gladbach, Germany, 130-095-999), according to the manufacturer's instructions. Recombinant interleukins (ILs) IL-7 (5 ng/mL; Peprotech, Rocky Hill, NJ, 200-07) and IL-2 (20 IU/mL; Novartis, Basel, Switzerland, Proleukin) were added at day 1 and days 3 to 6, respectively. On day 10, a similar round of stimulation was performed in the presence of IL-2 until day 19. HPV16-E7 reactive T cells were isolated by interferon (IFN)-γ T-cell magnetic sorting using the IFN-γ Secretion Assay – Cell Enrichment and Detection Kit (PE), human (Miltenyi Biotec, 130-054-201) according to the manufacturer's instructions. HPV16-E7–specific CD4 T-cell clones were generated according to a previously described procedure. 23
HPV16-E7/HLA-DRB1*04-specific TCR vectorization and retroviral supernatant production
The TCR α and β chains obtained from HPV16-E7-specific and HLA-DRB1*04-restricted CD4 T-cell clones were identified from RNA-isolated samples (Qiagen, Hilden, Germany, Rneasy Mini Kit) using a TCRβ-PCR 24 and multiplex TCRα-PCR, amplifying respectively the VDJ and VJ rearrangements of the TCR β and α chains, including hypervariable CDR3 regions. Sequences of cloned PCR products were identified after alignment against the IMGT database. 25 The sequence of both TCR constant regions were exchanged by minimally murinized variants and mutated to create an additional disulfide bond. DNA coding for both tagged TCR α and β chains, 26 linked by a 2A peptide linker, was synthetized (Eurofins MWG, Luxembourg) and cloned into a pSFG retroviral vector backbone. This vector (LTR-iCasp9-2A-TCRβ-2A-TCRα-IRES-ΔCD19-LTR) harbored a ΔCD19 selection transgene and an inducible caspase-9 (iCasp9) suicide gene safety cassette. 27 Retroviral vector DNA was transfected into PG13 cell line using Lipofectamine LTX Reagent (Fisher, A12621) according to the manufacturer's instructions. PG13 cells producing retroviral particles were magnetically sorted using CD19 microbeads (Miltenyi Biotec, 130-050-301) following the manufacturer's instructions. Supernatant of these modified cells was harvested and stored at −80°C for further utilization.
T-cell retroviral transduction, selection, and expansion
PBMCs of HLA-DRB1*04 healthy donors were isolated by centrifugation on Ficoll gradient and activated by anti-CD3/CD28 microbeads (Fisher, 111.31D) according to the manufacturer's instructions. Bead-attached T cells were magnetically isolated and cultured in RPMI-1640 medium with 10% human serum in the presence of 500 IU/mL IL-2. The complete medium was renewed every 2–3 days until day 10 of culture. On day 2, activated T cells were transduced using previously thawed HPV16-E7/HLA-DRB1*04-specific TCR retroviral supernatant, whose retroviral particles were trapped on RetroNectin (Takara, Shiga, Japan, T100B). An activated and untransduced cellular counterpart was further cultured. On day 6, transduced T cells were magnetically sorted using CD19 microbeads.
TCR transgenic T-cell functionality assessment
Transgenic TCR functionality and specificity were evaluated by co-culturing transduced and untransduced T cells with allogeneic HLA-DRB1*04 or HLA-DRB1*15 BLCLs. These BLCLs were previously treated or not with an anti-HLA-DP (Leinco, St. Louis, MO, H260), anti-HLA-DQ (Bio-Rad, Marnes-la-Coquette, France, MCA3796), or anti-HLA-DR (BD Biosciences, San Jose, CA, 555809) antibody for 30 min before pulsing with the HPV16-E7 peptide pool, HPV16-E76-25, or HPV16-E770-89. Nonspecific stimulation of transduced and untransduced T cells was carried out with phorbol 12-myristate 13-acetate (5 μg/mL, Sigma-Aldrich, Hampton, NH, P1585) and ionomycin (50 ng/mL, Sigma-Aldrich, I0634) (PMAi). To establish the recognition of processed antigen by the transgenic TCR, allogeneic monocyte-derived dendritic cells (MoDCs) were obtained from HLA-DRB1*04 PBMCs, and SiHa and C33A cell lysates were generated as previously described. 28 These lysates were pulsed on immature MoDCs for 24 h at 37°C. For the last 6 h of culture, lipopolysaccharide (1 μg/mL, Sigma, L2630) was added. After extensive PBS 1X (Fisher, 11530546) washing, mature pulsed-MoDCs were cocultured with transduced or untransduced T cells. Co-cultures were performed at a 1:1 cell ratio for 18 h at 37°C before IFN-γ secretion assessment. For iCasp9 safety cassette functionality assessment, transduced and untransduced T cells were cultured in the presence or absence of AP1903 dimerizer (rimiducid), kindly provided by Bellicum Pharma, Houston, TX, for 24 h before cell viability investigation.
Immunostaining and flow cytometry analysis
Membrane staining was performed using fluorescein isothiocyanate (FITC)-conjugated anti-CD3 (BD Biosciences, 555332), Brilliant Violet 421-conjugated anti-CD3 (BD Biosciences, 562426), FITC-conjugated anti-CD4 (Diaclone, Besançon, France, 954.031.010), phycoerythrin (PE)-conjugated anti-CD8 (Diaclone, 854.962.010), allophycocyanin (APC)-conjugated anti-CD19 (Miltenyi Biotec, 130-091-248), APC-conjugated anti-mouse (Jackson Immunoresearch, West Grove, PA, 115-135-164), Brilliant Violet 421-conjugated anti-murine β chain (mTCRβ) (BD Biosciences, 562839) antibodies, and Fixable Viability Dye eFluor 780 (eBiosciences, Hampton, NH, 65-0865-14). Intracytoplasmic IFN-γ and IL-2 detection was performed after membrane staining with BD Cytofix/Cytoperm Plus kit (BD Biosciences, 555028), following manufacturer instructions and using APC-conjugated anti-IFN-γ (BD Biosciences, 554702), Brilliant Violet 421-conjugated anti-IFN-γ (BD Biosciences, 562987), and PE-conjugated anti-IL-2 (BD Biosciences, 560709) antibodies. Cell viability was evaluated with Pacific Blue-conjugated anti-CD3 and APC-conjugated anti-CD19 antibodies, Annexin V FITC, and 7AAD staining in Trucounts supply, as previously described. 29 Flow cytometry analysis was performed using a BD FACSCANTO II flow cytometer and the BD FACS Diva software (BD Biosciences).
ELISA
IFN-γ secretion in the cell culture supernatants was measured by ELISA (Diaclone, 851.560.010) according to the manufacturer's instructions. All analyses were performed in duplicate. Absorbance at 450 nm was read using a Tecan spectrophotometer and the XFluor4 software.
Western blotting
Cells were collected in Tris-buffered saline (TBS) and resuspended in lysis buffer (50 mM Tris/HCl, pH 7.4, 150 mM NaCl, 0.1% SDS, 1% Nonidet P-40, 0.5% Na deoxycholate, 1 mM EDTA) with protease inhibitor (Roche Diagnostics, Meylan, France, 4693132001). After sonication, total protein concentrations were determined by Bio-Rad Protein Assay (Bio-Rad, 5000006). Then, 40 μg of protein sample was subjected to SDS-PAGE and transferred onto Hybond TM-P polyvinylidene difluoride membrane (GE Healthcare, Amersham, UK, 10600021). The membrane was blocked overnight at 4°C with 0.1% Tween 20 in TBS (TBS-Tween) supplemented with 5% nonfat milk and incubated for 2 h with primary antibodies, including anti-HPV16-E7 (Santa Cruz, Dallas, TX, sc-65711) and anti-β actin (Sigma-Aldrich, A5441) antibodies. After being washed with TBS-Tween, the membrane was incubated with appropriate horseradish peroxidase–conjugated secondary antibodies for 1 h and revealed by chemiluminescence with Pierce ECL2 Western Blotting Substrate (Thermo Scientific, Waltham, MA, 32132) using ChemiDoc XRS+ with image lab software (Bio-Rad).
Results
Isolation of a MHC class II–restricted HPV16-E7–specific TCR
For the generation of HPV16-E7–specific CD4 T cells, PBMCs from a patient in complete remission after an HPV16-positive metastatic anal cancer were pulsed with a pool of 15mer sequences with 11–amino acid overlap, covering the complete sequence of the HPV16-E7 protein. After 10 days of stimulation, cells were restimulated with the HPV16-E7 peptide pool. On day 20, IFN-γ secreting T cells were sorted and expanded (Fig. 1A). Then, specific CD4 T cells were isolated by limiting dilution and further expanded (Fig. 1B). The TCR α and β chains expressed by one CD4 T cell clone were sequenced in order to generate recombinant retroviruses encoding the appropriate TCRs. Murine TCRα/β constant region sequences were used to enhance the reactivity of TCRs and avoid recombination between transgenic and endogenous TCR, and a CD19 selection marker was included in the retroviral vector backbone alongside the iCasp9 suicide gene (Fig. 1C).
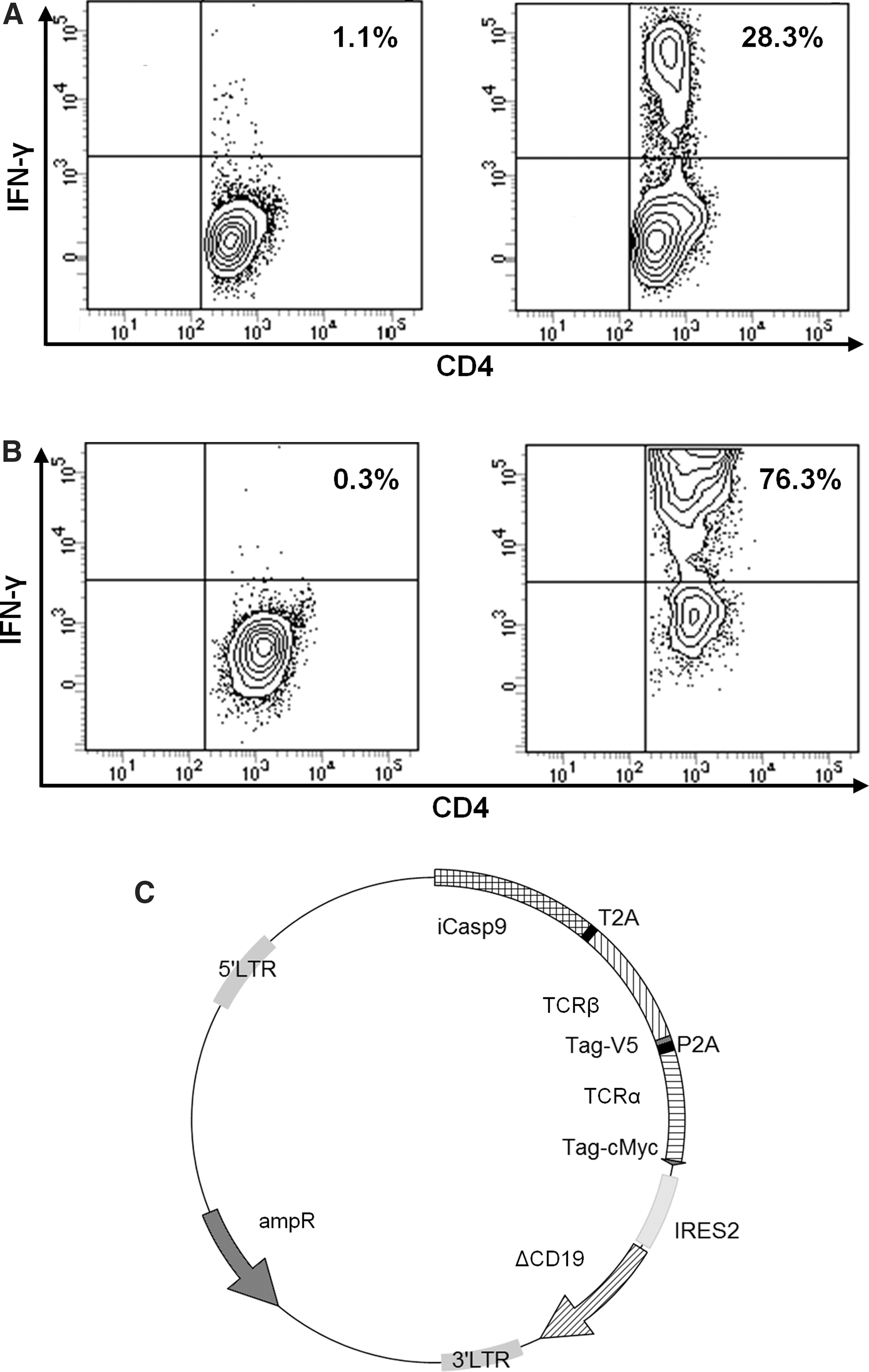
Isolation of HPV16-E7–specific T-cell clone and generation of a transgenic T-cell receptor (TCR).
MHC class II–restricted HPV16-E7–specific TCR transgenic T-cell functionality
We first validated the functionality of TCR transgenic T cells. A transduction efficiency of 21.0 ± 12.2% was obtained after 6 days of culture as revealed by CD19 staining (Fig. 2A and B). After one step of CD19+ cell sorting, the transfected cell purity reached 88.0 ± 15.6% (Fig. 2A and B). The transgenic TCR expression on the cell surface was then determined by mTCRβ staining (Fig. 2C). To assess the functionality of the suicide gene, the killing efficiency was evaluated after AP1903 exposure (Fig. 2D). The mean percentage of cell death observed was 86.8 ± 7.22%, revealing the efficient expression of the iCasp9 suicide gene. After 14 days, untransduced and transduced T cells displayed a similar phenotype, with the majority of cells showing an effector memory phenotype (CD45RA−CD45RO+CD95+CCR7−) (data not shown). Thereafter, T-cell functionality was evaluated after an overnight stimulation with the HPV16-E7 peptide pool. As shown in Fig. 2E, while untransduced T cells did not produced IFN-γ, 14.1 ± 0.929% of transduced T cells were IFN-γ positive. These results validated the retroviral vector and the functionality of the HPV16-E7–specific transgenic TCR.
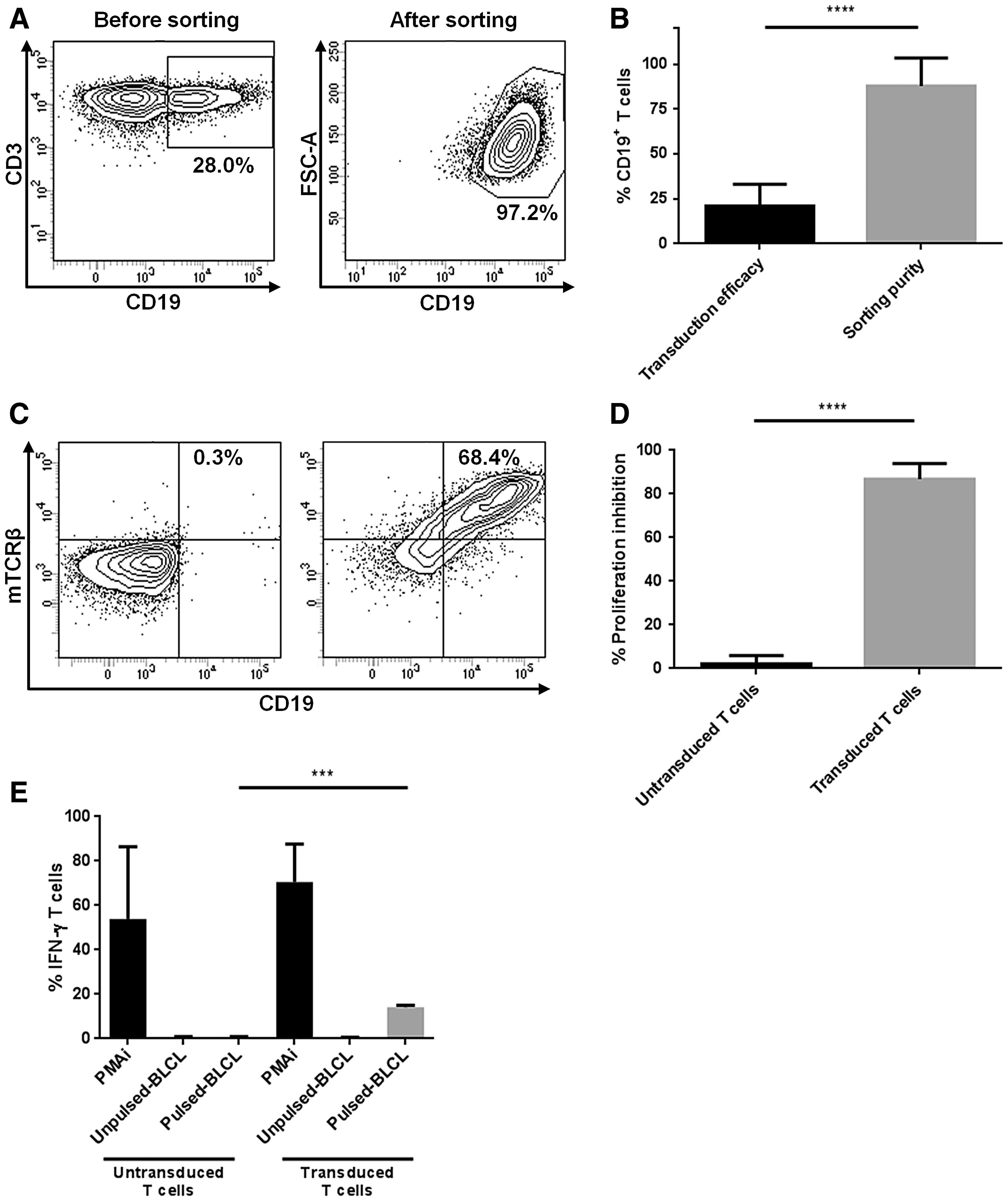
HLA-DRB1*04–restricted HPV16-E7–specific TCR transgenic T-cell functionality.
TCR transgenic T-cell specificity
The HLA restriction of transduced T cells was determined by testing the recognition of HLA-DR-matched BLCLs pulsed with the HPV16-E7 peptide pool and by using HLA-class II blocking monoclonal antibodies. Transduced T cells recognized peptide-pulsed HLA-DRB1*04 BLCLs but not peptide-pulsed HLA-DRB1*15 BLCLs (Fig. 3A). TCR transgenic T cells were thus specific for the HP16-E7/HLA-DRB1*04 complex.
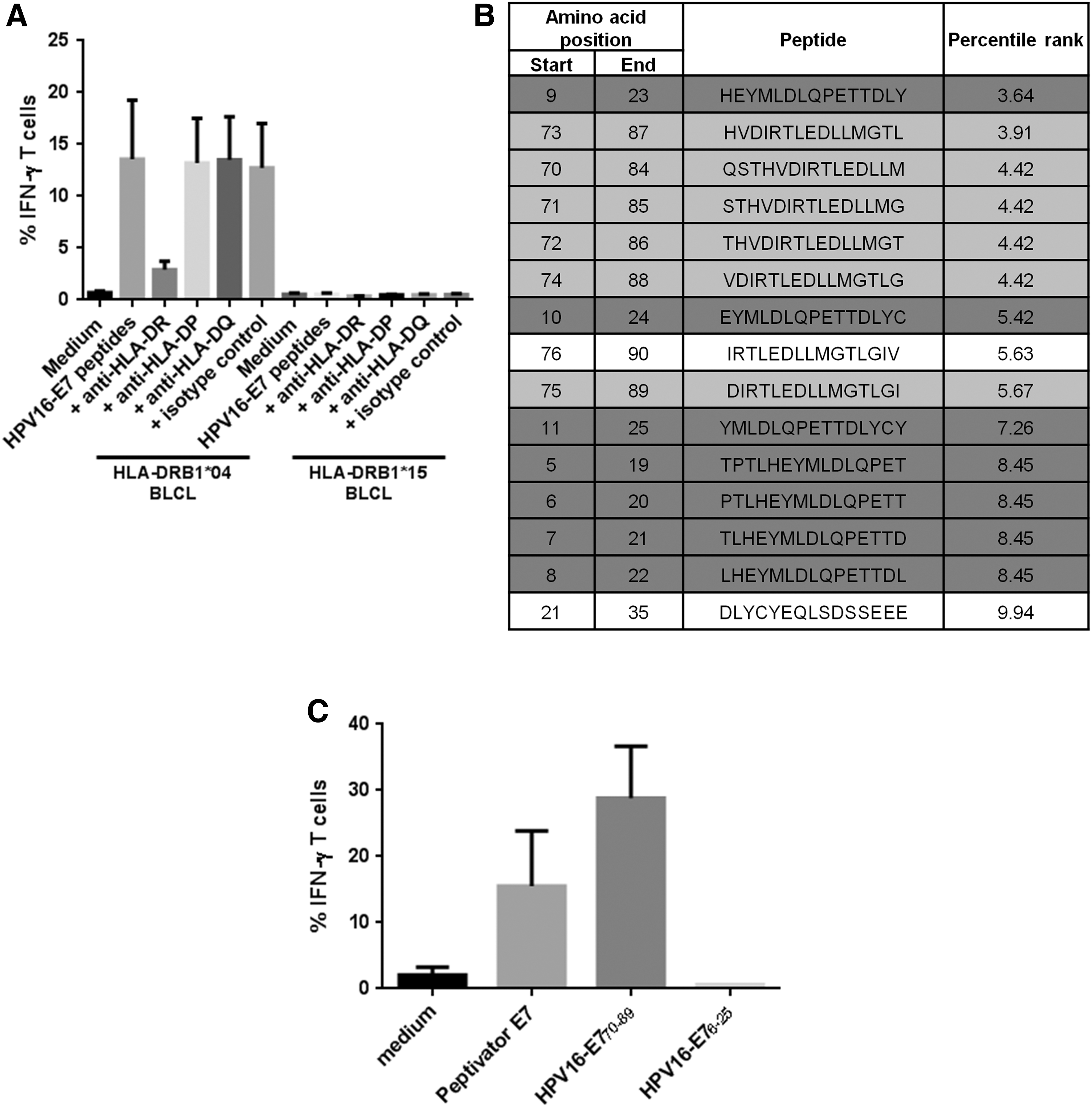
HLA-DRB1*04–restricted HPV16-E7–specific TCR transgenic T cell specificity.
HPV16-E7 is a 98–amino acid protein that contains two 20mer peptides (positions 6–25 and 70–89 on the protein sequence) encompassing 15mer peptides able to bind to the HLA-DRB1*04, with a high stability as predicted by the IEDB 22 (Fig. 3B). These two 20mer peptides were tested for their recognition by transduced T cells after pulsing onto HLA-DRB1*04 BLCLs. Only the sequence HPV16-E770-89 (QSTHVDIRTLEDLLMGTLGI) was recognized (Fig. 3C). TCR transgenic T cells were thus specific for the HPV16-E770-89/HLA-DRB1*04 complex.
Both CD4 and CD8 TCR transgenic T cells were HPV16-E770-89/HLA-DRB1*04 reactive
As CD4 and CD8 T cells were equivalently transduced (Fig. 4A), we assessed the contribution of the CD4 and CD8 co-receptors to the reactivity of transduced T cells with the HPV16-E770-89/HLA-DRB1*04-specific TCR. Controls included untransduced T cells. Both transduced CD8 and CD4 T-cell populations secreted IFN-γ and IL-2 when cultured with appropriate targets (Fig. 4B). However, CD8 T cells showed significantly reduced cytokine production compared to CD4 T cells. Nevertheless, transduced CD8 T cells recognized HPV16-E770-89–pulsed HLA-DRB1*04 BLCLs with an EC50 of 45 nM or 50 nM based on IFN-γ or IL-2 secretion, respectively (Fig. 4C). Based on IFN-γ or IL-2 secretion, the functional avidity of transduced CD4 T cells was slightly higher than transduced CD8 T cells (EC50 = 20 nM vs. 50 nM; Fig. 4C). Nonetheless, we did not formally assess the specific peptide naturally presented on HLA-DRB1*04.
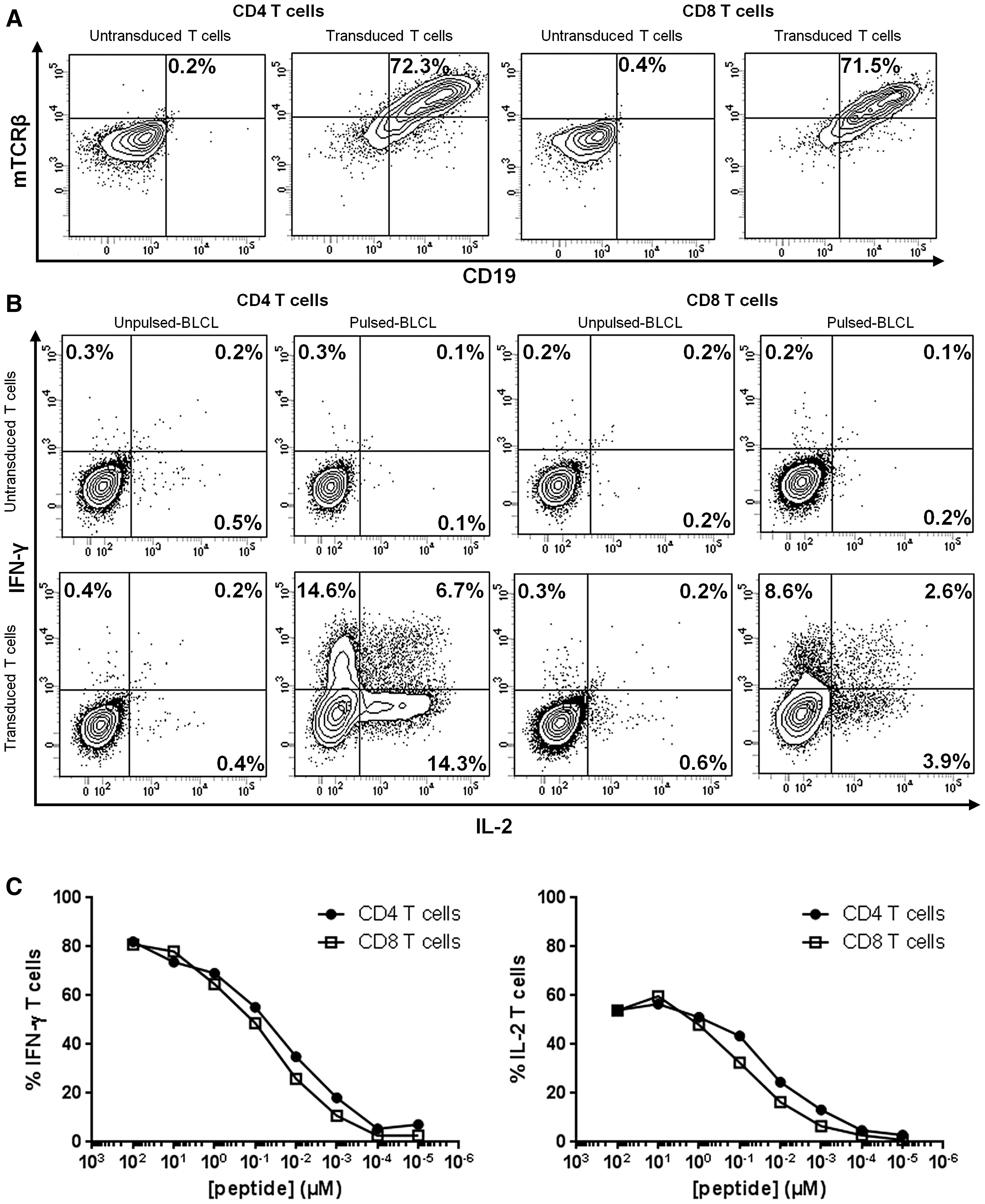
Functionality of TCR transgenic CD4 and CD8 T cells.
TCR transgenic T cells recognized HPV16-E770-89/HLA-DRB1*04 complex presented by DCs
One potential concern with such transgenic TCR was whether it could directly recognize endogenous HPV16-E7–expressing target cells, rather than exogenously peptide-pulsed cells. To assess the natural tumor cell recognition by HPV16-E770-89/HLA-DRB1*04–specific T cells, we generated immature MoDCs from HLA-DRB1*04 PBMCs and pulsed them with cell lysates of HPV16-E7 negative and positive cell lines, respectively, C33A and SiHa (Fig. 5A). After their maturation, DCs were co-cultured with transduced or untransduced T cells and IFN-γ secretion was evaluated (Fig. 5B). Transduced T cells co-cultured with C33A or SiHa lysate-pulsed MoDCs secreted 9.54 ± 5.87 μg/mL and 32.1 ± 19.6 μg/mL of IFN-γ, respectively. To assess if the functional reactivity of transduced T cells was due to the transgenic TCR rather than endogenous HPV-specific activities of donor T cells or alloreactivity, we verified that untransduced T cells reacted at the same level to C33A- and SiHa-pulsed MoDCs (data not shown). The results indicated that HPV16-E770-89/HLA-DRB1*04 complexes naturally presented by DCs are recognized by TCR transgenic T cells.
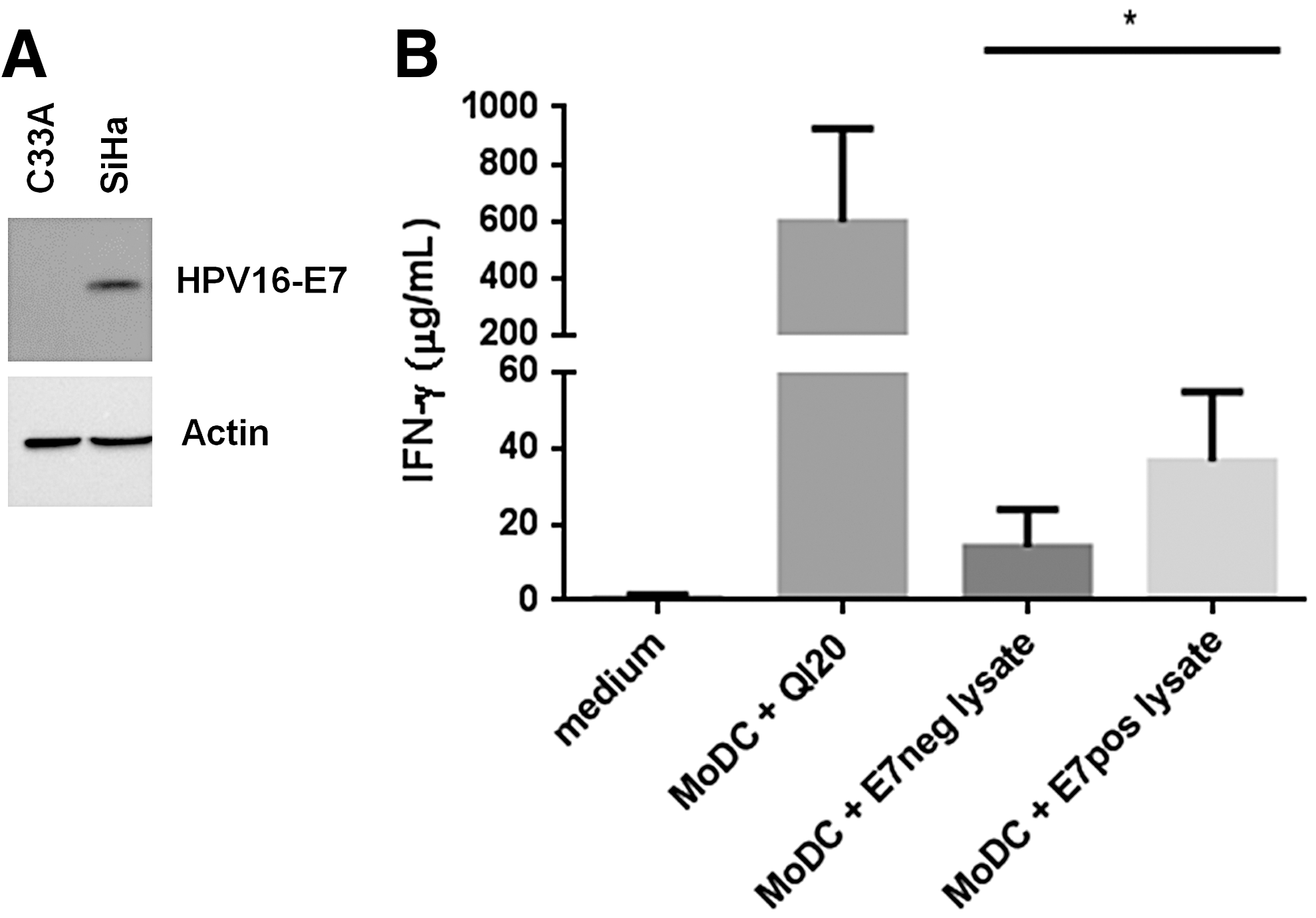
TCR transgenic T-cell recognition of HPV16-E770-89/HLA-DRB1*04 complex presented by dendritic cells.
Discussion
High-risk HPV infection is a causal factor in oropharyngeal and gynecological malignancies, which allows treating these cancers by HPV-targeting immunotherapy. Several successful prophylactic vaccines have been developed to prevent diseases associated with HPV, however, they are not effective in therapeutic settings, leaving unvaccinated individuals unprotected. 30 In 2016, vaccination coverage among adolescents in the United States was 60.4% for at least one dose of HPV vaccine. 31 In France, the proportion is less than 20%, which is among the lowest in Europe. 32 While a gender-neutral approach has been adopted by some countries and shows declines in infections with HPVs covered by vaccines, 33 heterosexual men are not systematically targeted by immunization campaigns, although oral HPV infection leads to 22-fold increased risk of developing head and neck squamous cell carcinoma. 34 Altogether, there is still a need to develop treatments for HPV-associated malignancies. One major hurdle is that CTL frequency in the blood is extremely low in the context of HPV16-induced malignancies, requiring multiples rounds of in vitro stimulation to obtain highly differentiated HPV16-specific CTLs. 9,10 Furthermore, MHC class I surface expression has been found to be impaired in HPV-positive cancer cells, limiting the clinical efficacy of MHC class I–restricted approaches. 19,35 Emerging evidence suggests that Th activity plays a major role in effective anticancer responses. 36,37 Although it has not yet reached a clinical setting, 16,38 adoptive transfer of in vitro redirected Th cells could confer clinical benefits through multiple mechanisms, such as inflammatory changes in the microenvironment and epitope spreading. 37,39 Altogether, TCR gene transfer through generation of high-avidity MHC class II–restricted T cells represents an attractive tool for the treatment of HPV-positive cancers.
In our study, HPV16-E7-specific CD4 T cell clones were obtained from a patient in complete remission from a metastatic anal cancer. These clones shared the same TCR and were used to design an HPV16-E7-specific transgenic TCR. The data herein addressed safety and efficacy issues by transducing T cells with this TCR. The TCR was modified to express the murine TCRα/β constant regions, which avoided the mispairing of transgenic TCR chains with endogenous ones and increased its surface expression. 40 The TCR α and β chains were linked by viral 2A peptides in order to ensure an equal molar ratio, and consequently an optimal pairing between them. 41 Furthermore, the iCasp9 was incorporated as a suicide gene to treat potential autoimmunity events or an endogenous TCR-induced graft-versus-host disease in an allogeneic setting. Indeed, the iCasp9/AP1903 suicide system was able to kill transduced T cells in a few hours and was not likely to be immunogenic because of its human origin. 29 Transduction efficiency was not above 30%, which is on average the result of one retroviral insertion, preventing lymphomagenesis. 42 Indeed, higher efficiencies would lead to multiple insertions, raising the possibility of insertional mutagenesis and oncogene activation. 42 Moreover, many studies have confirmed the need for high-affinity TCRs for cancer elimination, whereas others have suggested that an affinity plateau should not be bypassed, in order to avoid autoimmune side effects. Here, the functional avidity of transgenic CD4 T cells (20 nM) was similar to natural antitumor T cell clones. 43,44 We sought to exclude potential cross-reactivity for the peptide used in our study, allowing the generation of high-affinity TCRs without leading to autoimmune side effects. Using BLASTP programs, 45 we were not able to find a corresponding cross-reactivity with naturally expressed proteins in human tissues (data not shown).
The HPV16-E7/HLA-DRB1*04–specific TCR is equivalently expressed by CD4 and CD8 T cells, and both populations show reactivity against HPV16-E770-89–pulsed HLA-DRB1*04 BLCLs. The acquisition of MHC class II–restricted activity by CD8 T cells has already been described by others. 16 We observed that redirected CD4 T cells produced higher levels of IFN-γ and IL-2 than CD8 T cells. Nonetheless, redirected CD8 T cells were functional as well, even though they were slightly less avid than their CD4 counterparts, suggesting that the TCR affinity is strong enough to stimulate T cells without CD4 co-receptor requirement. It would be interesting to evaluate the functionalities of these MHC class II–restricted CD8 T cells because they may lead to positive as well as detrimental effects.
An additional concern about TCR transgenic T cells is their HLA allele restriction. Nonetheless, several TCRs have degenerated recognition capacity. 43 The HPV16-E770-89 peptide has already been predicted to bind several HLA class II molecules, such as DRB1*01, DRB1*03, DRB1*04, DRB1*07, and DRB1*15. 46 Our transgenic T cells recognized HPV16-E770-89–pulsed HLA-DRB1*04 cells and not HLA-DRB1*15 cells, however, we have not evaluated other MHC II alleles. HLA-DRB1*04 type is the most frequent in the white American population (30.1%). 47 Moreover, the HLA-DRB1*04 allele family is significantly associated with cervical squamous cell carcinoma in white women. 48 Altogether, these findings highlight the interest for HLA-DRB1*04 targeting in the context of transgenic TCR-based immunotherapy.
Th1 subsets that produce IFN-γ, IL-2, and tumor necrosis factor (TNF)-α play a critical anti-tumor role through the coordination of cell-mediated immunity against cancer cells, whereas Th2 or Treg profiles may lead to tumor tolerance. 49 We have noted that our redirected T cells produced high levels of IFN-γ and IL-2, which is potentially indicative of a Th1 profile. Nonetheless, the use of unselected CD4 T cells may lead to TCR transfer into regulatory T cells inducing unwanted suppressive T-cell reactivity. Moreover, even if cells are transduced with the same TCR, the cytokine profile is likely to vary. Adoptive cell therapy using Th cells with Th2 or Treg profiles might strengthen tumor tolerance. It may be useful to more precisely characterize the profile of redirected T cells by analyzing a broader range of cytokines. Another approach would be to generate redirected Th1 cells by sorting naive T cells before transduction and by stimulating these cells with IL-2 and IL-12 cytokines and IL-4–blocking antibodies. Furthermore, polyfunctional T cells may represent the more effective cell population both in infection 50 and cancer immunity. 51 Nevertheless, Th cell plasticity should be kept in mind while conducting Th-based immunotherapies.
In summary, we have isolated from a patient in complete remission from a metastatic HPV-related anal cancer a CD4 T-cell clonotype targeting the HPV16-E7 oncoprotein, and we have characterized redirected T cells with its HLA-DRB1*04-restricted HPV16-E7 TCR. The transgenic TCR is highly expressed in redirected T cells and confers the expected MHC class II specificity. Of note, the TCR was functional in both CD4 and CD8 T cells, and TCR transgenic T cells were able to recognize the endogenously processed HPV16-E770-89 peptide. We speculate that a tumor-targeting Th cell strategy may be a promising approach to mobilize patient immune system while overcoming tumor tolerance in the context of HPV16-induced malignancies. The favorable properties of these redirected T cells in vitro support the fact that they may confer clinical benefits, but this strategy has not been demonstrated yet. The efficient expression of the suicide gene and the incorporation of the murine TCRα/β constant regions into the construct, in addition to the TCR being derived from a long-term survivor, strengthen the safety profile of this approach. In the future, a Th cell strategy may be investigated alone or in combination with other therapeutic procedures in order to treat patients with HPV-related malignancies.
Footnotes
Acknowledgments
We thank Dr. Negar Seyed, Julie Frejaville, and Dr. Charline Marotel for technical assistance. We also thank the patient and healthy volunteers who participated in this study. This work was supported by the Ligue Nationale contre le Cancer, and by the MiMedi project funded by BPI France (grant No. DOS0060162/00) and the European Union through the European Regional Development Fund of the Region Bourgogne-Franche-Comte (grant No. FC0013440). C.M. has benefited from a fellowship from Nancy Regional University Hospital.
Author Disclosure
No competing financial interests exist.