
Abstract
Bone-marrow mesenchymal stem cells (BMSCs) have been used for systemic delivery of therapeutic genes to solid tumors. However, the optimal treatment time post-BMSC implantation and the assessment of the long-term fate of therapeutic BMSCs post-tumor treatment are critical if such promising therapies are to be translated into clinical practice. An efficient BMSC-based therapeutic strategy has been developed that simultaneously allows killing of tumor cells, inhibiting of tumor angiogenesis, and assessment and eradication of implanted BMSCs after treatment of glioblastoma. BMSCs were engineered to co-express the angiogenesis inhibitor kringle 5 (K5) of human plasminogen, under the control of the cytomegalovirus promoter (CMV) and the human sodium–iodide symporter (NIS), involved in uptake of radioisotopes, under the control of early growth response factor 1 (Egr1), a radiation-activated promoter. A significant decrease in tumor growth and tumor angiogenesis and a subsequent increase in survival were observed when mice bearing glioblastoma were treated with 188Re post-therapeutic intravenous BMSC implantation. Furthermore, the systemic administration of 188Re post-tumor treatment selectively eliminated therapeutic BMSCs expressing NIS, which was monitored in real time by 125I micro single photon emission computed tomography/computed tomography imaging. Meanwhile, the Egr1 promoter induced a 188Re radiation positive feedback effect absorbed by NIS. After intravenous BMSC implantation, BMSCs levels in the tumor and lung both peaked on day 10 and decreased to the lowest levels on days 24 and 17, respectively. These findings suggest that day 17 post-BMSC implantation could be an optimal time for 188Re treatment. These results provide a new adjuvant therapy mediated by BMSCs for glioblastoma treatment.
Introduction
G
Cell-mediated antitumor gene therapy, in which cells transport the therapeutic gene to the solid tumor, is a new approach for solid tumor targeting therapy. Bone-marrow mesenchymal stem cells (BMSCs) have unique advantages in local and disseminated malignant tumor targeting 5,6 and the ability to cross the blood–brain barrier and the blood–tumor barrier in glioblastoma. 7 Additionally, BMSCs have a reliable source, as they can be isolated and expanded with relative ease from bone marrow, are easy to obtain, lack immunogenicity, and can be used for autologous implantation with no ethical issues, suggesting that BMSCs may be an ideal cell carrier for gene therapy of glioblastoma. 8 In addition to these properties, the increase in vascular permeability and the release of various cytokines after tumor irradiation can also enhance BMSC migration to the tumor site. 9
However, in order to avoid continuous access of antitumor agents to normal tissue and to circumvent any malignant transformation of BMSCs, it is critical to develop and test BMSCs that simultaneously allow killing of tumor cells, follow the fate of therapeutic BMSCs with a clinically relevant noninvasive imaging method, and ultimately selectively eradicate BMSCs post-tumor treatment. The sodium–iodide symporter (NIS) is an intrinsic membrane glycoprotein responsible for iodide transport, which is expressed in thyroid follicular cells and others cells containing a lactoperoxidase system. 10,11 NIS is involved in the uptake of several isotopes, including 125I, 99mTc, 188Re, and 131I. 12 131I radioiodine therapy mediated by NIS has been shown to be a safe and effective approach for the imaging and treatment of thyroid disease. 13 The authors and other researchers have successfully introduced ectopic expression of NIS by virus for imaging and therapy in nonthyroidal carcinoma. 14 –18
To reduce immunogenicity and increase tumor tropism, virus intratumoral injection or stable therapeutic gene expression tumor cell lines were used in these studies, both of which, however, limit the clinical application, especially for patients with a disseminated malignant tumor or with tumors in deep tissues. Some studies 19,20 have reported that the early growth response factor 1 (Egr1), a radiation-sensitive promoter that can be activated by radiation, can initiate expression of downstream target genes efficiently and specifically. Based on this property, the Egr1 promoter is inserted upstream of the NIS gene. NIS absorbs radioisotopes, and the radiation emitted by the isotopes also activates the Egr1 promoter, inducing more expression of NIS. This forms a positive feedback loop to ensure tumor tissues have an adequate uptake of radionuclide. Among various radioisotopes, 188Re 21 can emit high energy γ and β rays, exerts strong biological effects, has a short half-life (16.9 h), and is easy to control, making it an ideal radionuclide for imaging and therapy.
Recent preclinical studies have suggested that radiotherapy in combination with antiangiogenic/vasculature targeting agents enhances the therapeutic ratio of ionizing radiation alone. 22 Previous work has shown that kringle 5 (K5) of human plasminogen or K5 combined with 131I absorbed by NIS can inhibit tumor angiogenesis. 15,16,23,24 However, these gene therapies were mediated by virus. In this study, an efficient BMSC-based therapeutic strategy was developed that simultaneously allows killing of tumor cells, inhibiting of tumor angiogenesis, and assessment and eradication of BMSCs after treatment of glioblastoma. BMSCs were engineered to co-express K5 under the control of the CMV promoter and NIS under the control of Egr1 promoter.
Then, 188Re combined with K5 expression was investigated for the treatment of glioblastoma, BMSC tumor targeting after intravenous (i.v.) implantation by dynamic micro single photon emission computed tomography/computed tomography (micro-SPECT/CT) imaging. The findings identify a novel therapeutic approach for glioblastoma.
Materials and Methods
Cell lines and animals
The U87 human glioblastoma cell line (ATCC, Manassas, VA), human umbilical vein endothelial cells (HUVECs), and the 293T cell line (Cell Bank of the Chinese Academy of Science, Shanghai, China) were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), and 1% penicillin and streptomycin. All cell lines were maintained at 37°C in a humidified 5% CO2 incubator.
BALB/c nude mice and Sprague–Dawley rats were purchased from Shanghai Slaccas Experiment Animal Corp. (Shanghai Institute for Biological Science, Shanghai, China). Animal studies were approved by the local Ethics Committee (Shanghai Jiaotong University School of Medicine) and performed according to ethical principles of animal experimentation.
Plasmid construction and lentiviral preparation
The NIS, K5 (His-tagged), and Egr1 genes were present in the laboratory. 15 The pLVX-CMV-Puro (puromycin) expression vector was purchased from Clontech (Takara, Dalian, China). The NIS, Egr1, and K5 genes were polymerase chain reaction amplified separately and then cloned into the pLVX-CMV-Puro vector to construct pLVX-CMV-K5-Egr1-NIS, pLVX-CMV-K5-Egr1-GFP, and pLVX-CMV-GFP-Egr1-NIS plasmids. The lentiviral particles were produced and amplified in 293T cells.
Stable transfection of BMSCs and U87 cells
BMSCs were isolated from bone-marrow aspirates of Sprague–Dawley rats (5 weeks old), propagated and characterized, as previously described, 25 and then infected with the above lentivirus. The stably transfected cell lines were termed BMSC-K5-NIS, BMSC-K5-GFP, and BMSC-GFP-NIS after puromycin selection.
To monitor the established glioma tumor by an in vivo optical imaging system, U87 cells were infected with GV260 lentivirus containing a luciferase reporter gene. The stably transfected cell lines were termed U87-Luc after puromycin selection.
Western blot analysis
BMSC-K5-NIS, BMSC-K5-GFP, BMSC-GFP-NIS, and BMSC cell lysates were prepared using standard methods. 25 To detect NIS and K5 expression, Western blot was performed using antibodies against the following proteins: NIS (Millipore, Boston, MA), His-tag (Abgent, Suzhou, China), and β-actin (Abgent).
125I uptake assay
Following transfection, iodide uptake of BMSC-K5-NIS, BMSC-K5-GFP, BMSC-GFP-NIS, and BMSC cells was determined under steady-state conditions, as described previously. 16 The day before the experiment, BMSC-K5-NIS, BMSC-K5-GFP, BMSC-GFP-NIS, and BMSC cells were plated (2 × 10 5 cells/well) onto 24-well plates. After 24 h, 500 μL of Hanks' balanced salt solution (HBSS) containing 3.7 kBq of 125I and 10 μmol/L of sodium iodide (NaI) was added. The cells were incubated at 37°C for 5–180 min, washed twice with ice-cold HBSS, and lysed using 0.5 mol/L of NaOH. The radioactivity (counts/min, cpm) of the cell lysates was measured using an automatic γ-counter (Shanghai Rihuan Company, Shanghai, China).
Cell irradiation
To study the influence of 188Re irradiation on the NIS expression (Egr1 promoted), BMSC-K5-NIS cells were irradiated with 188Re (Shanghai Kexing Pharmaceutical Company, Shanghai, China) at varying doses (0.4625–7.4 MBq/mL) for different time periods (0–48 h). NIS expression was detected by Western blot.
In vitro migration assay
To examine the effect of K5 on HUVECs, a Boyden Chamber-based assay was performed. HUVECs (5 × 103) were plated into the upper chamber of a 24-well transwell (Corning, Corning, NY) in DMEM without FBS and incubated for 6 h. The medium was removed, and the medium from different stable BMSC cell lines was loaded into the upper chambers. DMEM containing 10% FBS was added into the lower chambers. Following incubation for 12 h, the upper chamber was dismantled. The non-migrated cells were scraped from the upper surface of the membrane with cotton swabs. Then, the membrane was stained with crystal violet (Beyotime Institute of Biotechnology, Shanghai, China). Each sample was tested in quadruplicate, and a representative field in each well was counted at 10 × magnification to determine the number of migrated cells.
Orthotopic glioblastoma tumor model
BALB/c nude mice, aged 5–6 weeks, were used for orthotopic glioblastoma experiments, as previously described. 26 To establish glioblastoma xenografts, 5 × 10 5 U87-Luc cells were stereotactically injected in a 1.5 μL volume into the left striatum over a 10 min period based on the following coordinates: 1 mm anterior, 1 mm lateral from the bregma, and 3 mm deep from the cortical surface. Mice were injected intraperitoneally with D-luciferin (150 mg/kg body weight; Sciencelight, Shanghai, China) on day 14 after implantation, and the average luminescence (photons/s/cm2) of tumors was monitored by optical imaging (Xenogen IVIS, Alameda, CA). The tumor take rate was 100%.
Tumor tropism of BMSCs and 188Re irradiation
To examine tumor tropism, survival time, and distribution of BMSCs after i.v. implantation, 5 × 10 5 BMSC-GFP-NIS were injected i.v. into orthotopic U87-Luc tumor-bearing nude mice (n = 3) every 3 days a total of three times. 125I micro-SPECT/CT imaging was performed to monitor the migration and survival time of BMSC-GFP-NIS. Three-dimensional lung structures were constructed using CT images, and three-dimensional structures of U87-Luc tumors were constructed using 125I tumor uptake. U87-Luc tumor-bearing nude mice injected with phosphate-buffered saline (PBS) were used as controls (n = 3).
To detect iodide uptake by NIS promoted by the radiosensitive Egr1 promoter after 188Re irradiation, U87-Luc tumor-bearing mice were injected i.v. with 7.4 MBq of 125I 3 days after the last round of BMSC-GFP-NIS implantation, and 30 min later, micro-SPECT/CT imaging was performed. Twenty-four hours after micro-SPECT/CT imaging, 18.5 MBq 188Re was injected i.v. into the nude mice for Egr1 irradiation. Two days after 188Re injection, 7.4 MBq 125I micro-SPECT/CT imaging was performed again.
All of the images were processed and reconstructed using Nuclear v1.02 (Bioscan, Poway, CA). HiSPECT v1.4.2 (Bioscan) was used for image acquisition, and InVivoScope v1.44 (Bioscan) was used for image analysis. Regions of interest (ROIs) were drawn around the U87-Luc tumor and lung, the radioactivity per volume unit (Conc) in the ROIs was measured using InVivoScope v1.44 (Bioscan).
BMSC-mediated dual gene therapy in vivo
After establishing orthotopic U87-Luc tumors, the average luminescence (photons/s/cm2) of the tumors was measured by optical imaging. The mice were randomly assigned to five groups (n = 6 in each group), and the average tumor radiance in each group was roughly equivalent.
BMSC-K5-NIS, BMSC-GFP-NIS, BMSC-K5-GFP, BMSCs, or PBS (5 × 105) were injected i.v. into nude mice (three times every 3 days). Seventeen days after the final injection, 37 MBq of 188Re (twice every other day) was applied for radiation therapy. Tumor growth monitored by optical imaging and survival time of the animals was used to assess the therapeutic effects.
Survival of BMSC-GFP-NIS after 188Re radiation therapy
BMSC-GFP-NIS cells were implanted i.v. into U87-Luc tumor-bearing mice (n = 3) as above. 125I micro-SPECT/CT imaging and immunofluorescence were used to monitor the survival of BMSC-GFP-NIS cells in the orthotopic U87-Luc tumor before and after 188Re treatment according to the method above. The tumors from mice without 188Re radiation therapy were used as controls (n = 3).
Histology and immunofluorescence
Three days after the final round of BMSC-GFP-NIS implantation, U87-Luc tumor-bearing nude mice were sacrificed. Serial coronal cryosections (6 μm) from frozen brains were processed. The entire brain sections were subjected to hematoxylin and eosin (H&E) staining for visualization of the tumor mass and immunohistochemical analysis using mouse anti-NIS (Millipore) for visualization of the implanted BMSC-GFP-NIS cells. U87-Luc tumor sections were subjected to immunofluorescence analysis using mouse anti-NIS (Millipore) and mouse anti-CD31 (Dako, Glostrup, Denmark) combined with DyLight594 rabbit anti-mouse IgG (MultiSciences Biotech Co., Shanghai, China). Nuclei were labeled with 4′,6-diamidino-2-phenylindole (Beyotime Institute of Biotechnology), and images were acquired with a fluorescence microscope (Olympus, Tokyo, Japan).
Capillary density was defined as the number of CD31+ ECs/high-power field (hpf; 200 × ). Five high-power fields were counted per section from six sections/tumor tissue in three animals/group.
Statistical analysis
Data were analyzed by Student's t-test when comparing two groups and by analysis of variance when comparing more than two groups. Data are expressed as mean ± SD. p-Values <0.05 were considered significant. Survival times of the five mouse groups were compared using the log-rank test. All analyses were performed using GraphPad Prism v5 (GraphPad Software, La Jolla, CA).
Results
Construction of lentiviral vector and BMSC stable cell lines
The pLVX-CMV-K5-Egr1-NIS, pLVX-CMV-K5-Egr1-GFP, and pLVX-CMV-GFP-Egr1-NIS constructs were successfully generated and packaged. BMSCs were successfully isolated and cultured as per a previous study. 25 After lentivirus infection and puromycin selection, the stable cell lines BMSC-K5-NIS, BMSC-GFP-NIS, and BMSC-K5-GFP were generated. BMSCs were transfected with NIS linked to Egr1 promoter for positive radioisotope uptake of the construct. The treatment regimen consisted of systemic stem cell (carrying therapeutic gene K5) administration followed by systemic 188Re injections (Fig. 1a). Western blot analysis showed K5 protein expressed in BMSC-K5-NIS and BMSC-K5-GFP cells but not BMSC-GFP-NIS and BMSCs, and NIS protein expressed in BMSC-K5-NIS and BMSC-GFP-NIS cells but not BMSC-K5-GFP and BMSC cells (Fig. 1b). BMSC-K5-NIS and BMSC-GFP-NIS cells showed a significant increase in 125I uptake in vitro that peaked at 120 min and decreased only slightly at 180 min. There was no 125I uptake observed in BMSC-K5-GFP cells or BMSCs (Fig. 1c). These results revealed that BMSCs could coexpress K5 and NIS simultaneously and absorb radioisotope by NIS.
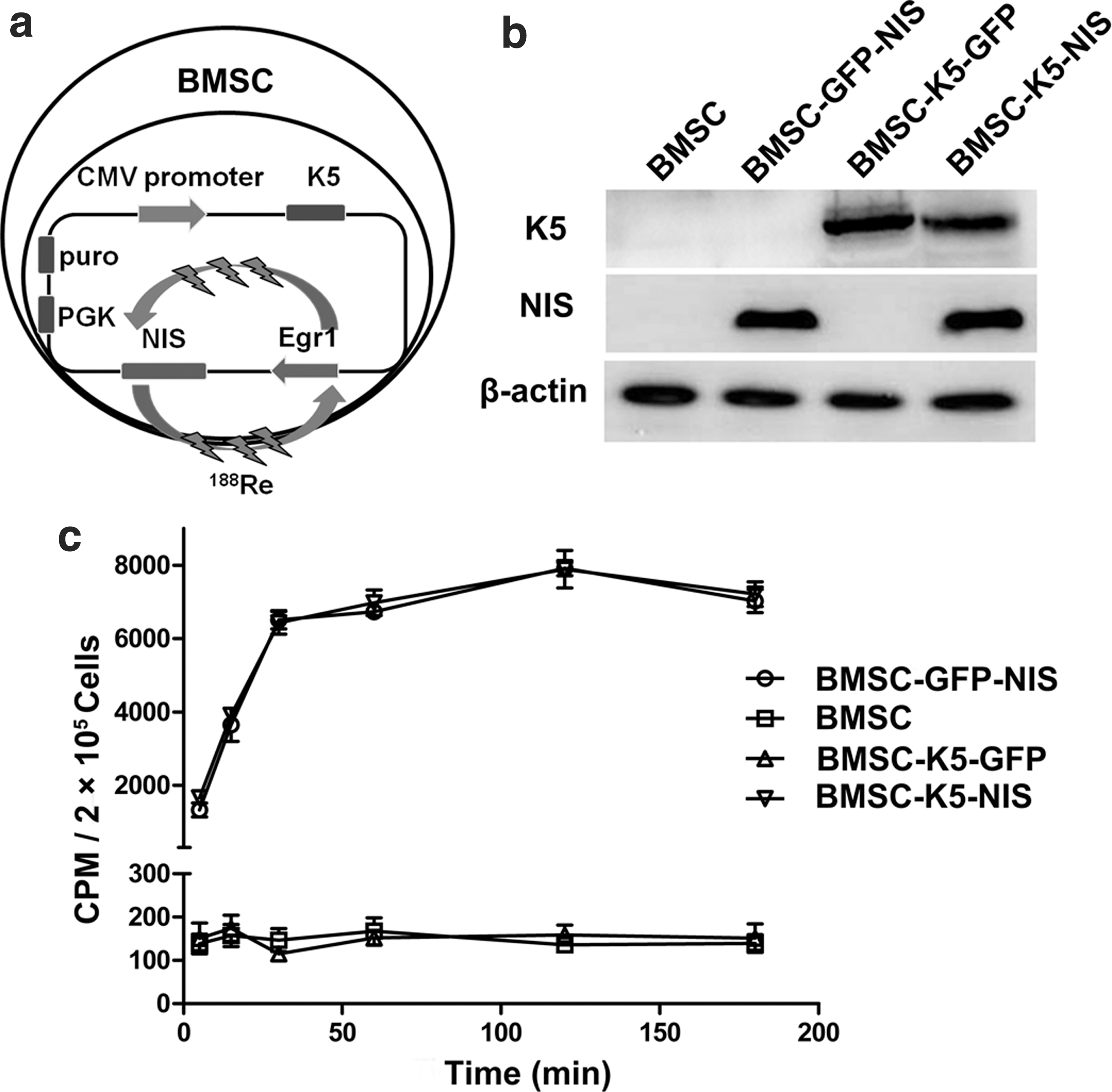
Schematic depiction of the CMV-K5-Egr1-NIS bone marrow mesenchymal stem cell (BMSC) construct, kringle 5 (K5) and sodium–iodide symporter (NIS) expression, and iodide uptake in vitro.
Migration assay in vitro, specific function of Egr1 promoter in vitro
Migration assays showed that using medium from BMSC-K5-NIS and BMSC-K5-GFP to culture HUVECs for 12 h resulted in approximately 50% of HUVECs migrating to the opposite membrane (Fig. 2a and b). In contrast, BMSC-GFP-NIS and BMSCs culture medium had no influence on HUVECs migration, with approximately 100% of the cells migrating to the opposite membrane (Fig. 2a and b). These results show that culture medium from BMSC-K5-NIS and BMSC-K5-GFP could significantly inhibit the migration of HUVECs.
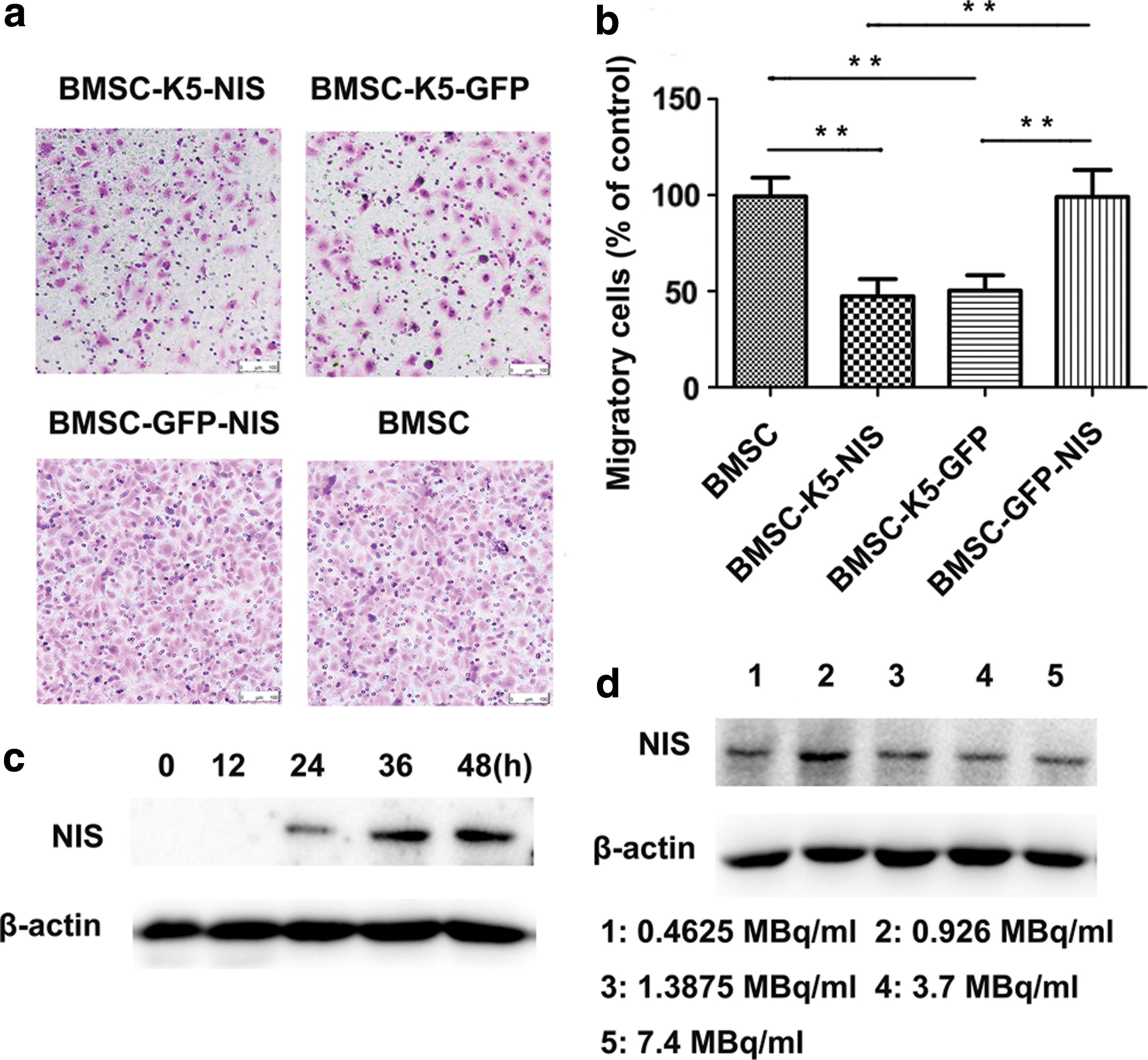
Migration assay in vitro, the function of Egr1 promoter in vitro.
After culturing with medium containing 1.85 MBq/mL of 188Re, Western blot analysis showed that NIS protein expression in BMSC-K5-NIS increased gradually with time, with the highest levels expressed at 36 h, and levels did not decrease by 48 h (Fig. 2c). The 36 h time point was selected as the optimum time for 188Re radiation to activate the Egr1 promoter. BMSC-K5-NIS cells were cultured with medium containing different concentrations of 188Re (0.4625–7.4 MBq/mL) for 36 h. Western blot showed that the highest expression of NIS protein was induced by 0.925 MBq/mL of 188Re (Fig. 2d).
Micro-SPECT/CT imaging in vivo and immunofluorescence analysis
The U87-Luc stable cell line was established successfully after infection with GV260 lentivirus (Fig. 3a). The orthotopic U87-Luc tumor in the brain could be monitored by optical imaging after D-luciferin administration (Fig. 3b). To detect the function of radiosensitive Egr1 promoter in vivo, 125I micro-SPECT/CT imaging was performed before and after 18.5 MBq of 188Re irradiation.
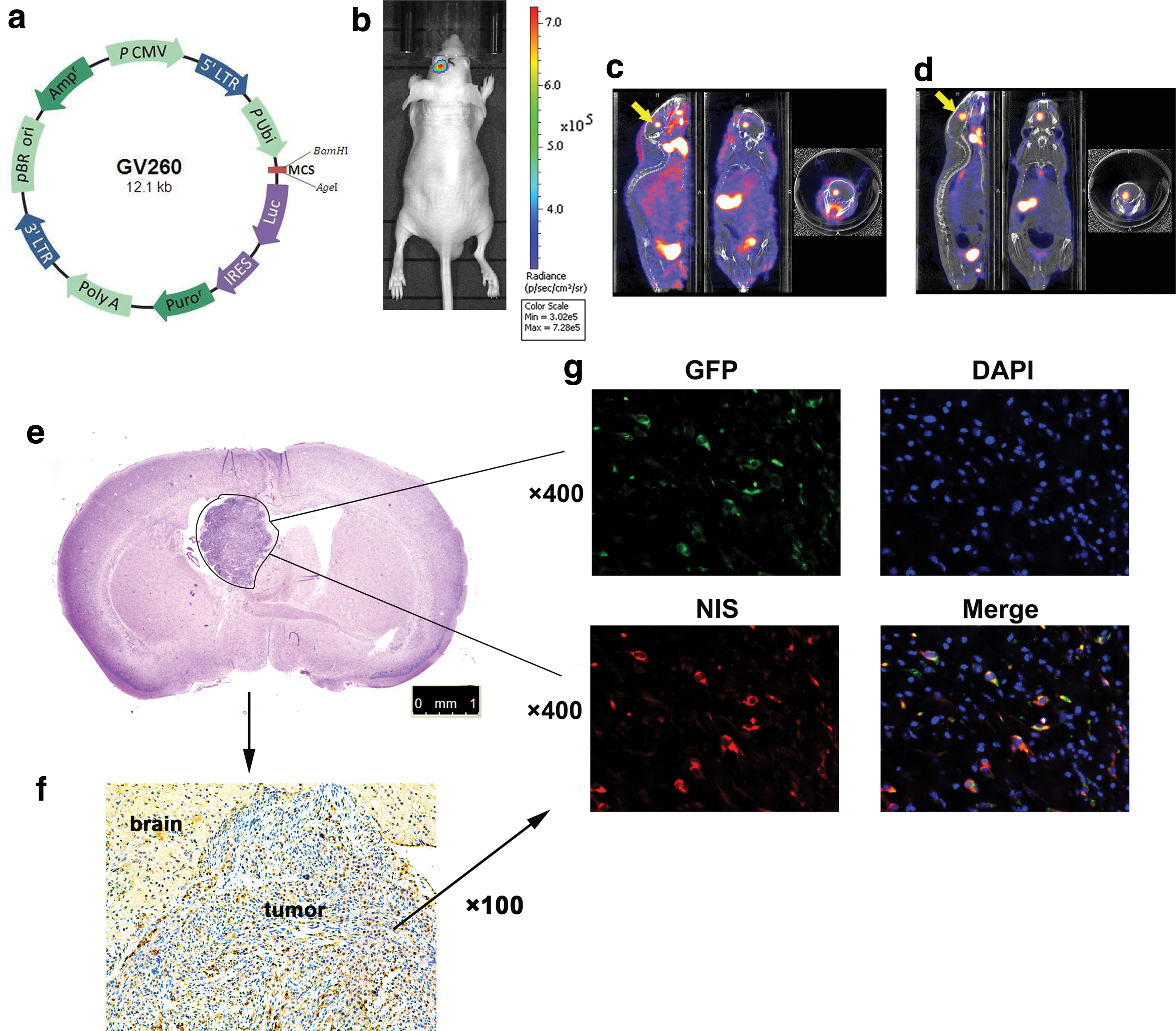
Optical and micro single photon emission computed tomography/computed tomography (micro-SPECT/CT) imaging in vivo, immunohistochemical and immunofluorescence analysis.
Increased radionuclide uptake was observed after 188Re radiation with 0.4212 ± 0.0102 × 10–3 MBq/mm 3 to 0.9863 ± 0.0093 × 10–3 MBq/mm 3 (Fig. 3c and d; c: before 188Re irradiation; d: after 188Re irradiation; n = 3). H&E staining showed the U87-Luc tumor in the brain (Fig. 3e). Immunohistochemical analysis showed NIS-expressed BMSC-GFP-NIS cells existed in the U87-Luc tumor but not in normal brain tissues (Fig. 3f). Fluorescence confocal microscopy pictures confirmed the existence of BMSC-GFP-NIS cells in the orthotopic U87-Luc tumor after their i.v. implantation (Fig. 3g).
BMSC-GFP-NIS cells' survival time after implantation
To follow BMSCs fate after implantation, micro-SPECT/CT imaging was performed using 125I, an optimal radiotracer to monitor NIS activity that has high uptake rate and low background activity. Micro-SPECT/CT imaging allowed specific detection of i.v. implanted BMSC-GFP-NIS cells on day 10, 17, and 24 in tumor-bearing nude mice (Fig. 4a). As shown in representative images, 125I uptake in the tumor and lung reached the highest levels on day 10 (conc: 0.401 ± 0.011 × 10–3 MBq/mm3 and 0.312 ± 0.026 × 10–3 MBq/mm3, respectively), then decreased over time, reaching the lowest levels on day 24 (tumor conc: 0.230 ± 0.017 × 10–3 MBq/mm3) and day 17 (lung conc: 0.227 ± 0.022 × 10–3 MBq/mm3), respectively. However, levels in the tumor on day 17 (conc: 0.303 ± 0.014 × 10–3 MBq/mm3) were higher than those in the lung. Concentrations in the lung on day 17 were similar to the control group (i.v. injection of PBS; conc: 0.247 ± 0.048 × 10–3 MBq/mm3; Fig. 4a and c). There was almost no change in 125I uptake in the tumor and lung in the control group over time (Fig. 4b and c). Together, these results showed that BMSCs fate can be detected by clinically relevant micro-SPECT/CT imaging, and day 17 post implantation may be an optimal time for radiation treatment, while day 10 treatment could possibly lead to severe lung injury.

BMSC-GFP-NIS cell survival after implantation.
Dual gene therapy in vivo
Mice were randomly assigned to one of five groups (n = 6 in each group) based on average luminescence (1–2 × 107 photons/s/cm2). After 37 MBq of 188Re treatment (twice every other day), the tumor growth in the BMSCs group and sham group increased quickly (monitored by optical imaging), and mice died in a short time (Fig. 5a and b). However, tumor growth in the BMSC-K5-NIS, BMSC-GFP-NIS, and BMSC-K5-GFP groups was inhibited, and mice survived longer, especially those in the BMSC-K5-NIS group (Fig. 5a and b). Kaplan–Meier survival curves showed that the median survival time for the BMSC-K5-NIS, BMSC-GFP-NIS, BMSC-K5-GFP, BMSC, and sham group was 54.5, 42, 41.5, 31.5, and 31.5 days, respectively (Fig. 5c).

Dual gene therapy in vivo.
Survival of implanted BMSC-GFP-NIS cells after 188Re radiation therapy
Post BMSC-GFP-NIS i.v. implantation, 125I micro-SPECT/CT imaging was performed. Radionuclide uptake was observed before 188Re treatment (Fig. 6a1 and b1) but not 5 days after 188Re treatment (Fig. 6a2) in the orthotopic U87-Luc tumor. However, 125I uptake was observed by micro-SPECT/CT imaging on day 5 in the absence of 188Re treatment (Fig. 6b2). BMSC-GFP-NIS cells were found in the U87-Luc tumor before 188Re treatment but not after treatment, as determined by immunofluorescence (Fig. 6c1, 2). These results confirmed that the implanted BMSCs expressing NIS can be eliminated selectively with 188Re in vivo.
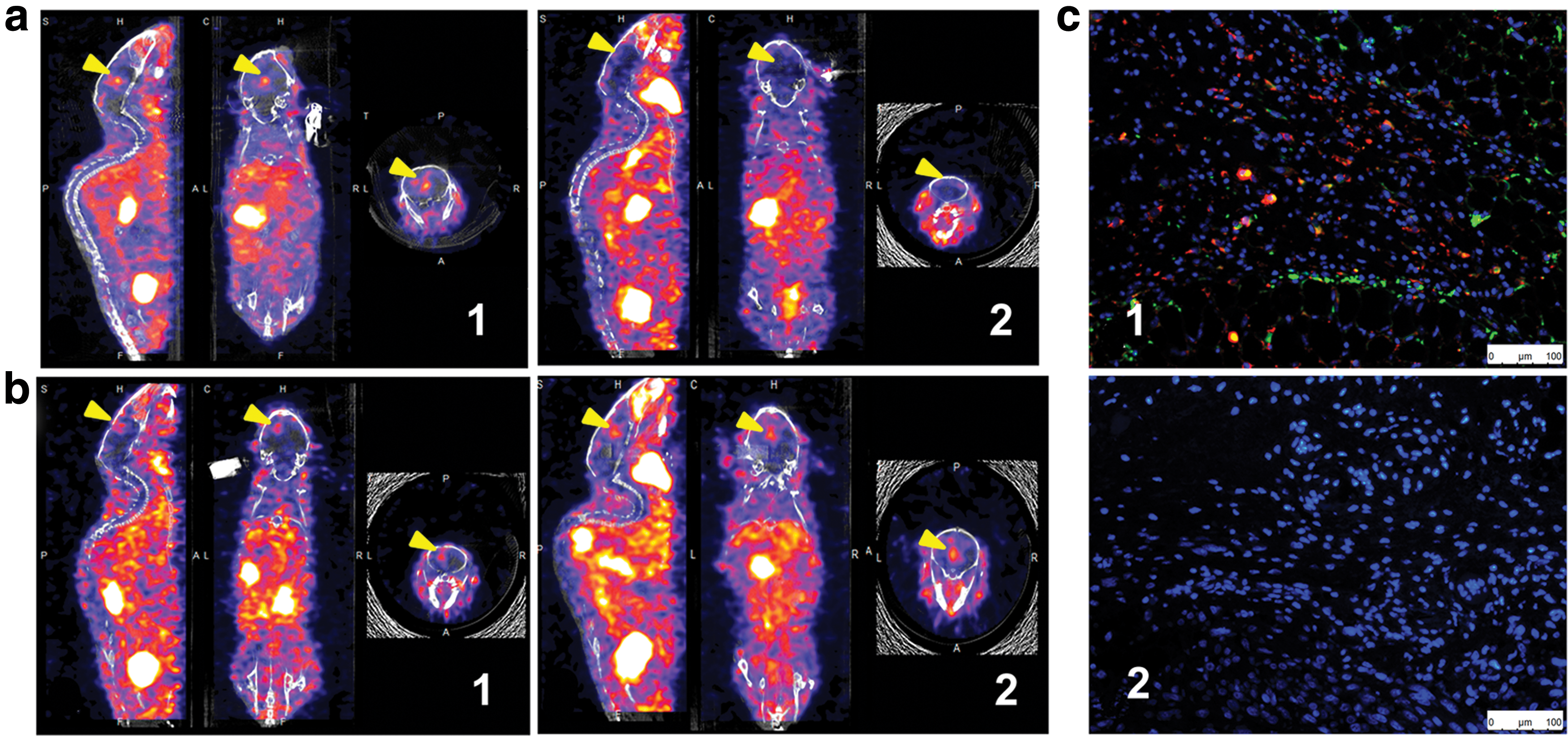
125I micro-SPECT/CT imaging and immunofluorescence.
Immunofluorescence analysis
Figure 7 depicts representative images of the indirect tumor xenograft immunofluorescence staining. Immunofluorescence revealed fewer CD31+ capillaries in the BMSC-K5-NIS group compared to the BMSC-K5-GFP (p < 0.05), BMSC-GFP-NIS (p < 0.01), BMSC (p < 0.01), and PBS (p < 0.001) groups. There were fewer CD31+ capillaries in the BMSC-K5-GFP group compared to the BMSC-GFP-NIS (p < 0.05), BMSC (p < 0.05), and PBS (p < 0.01) groups (Fig. 7a–f).
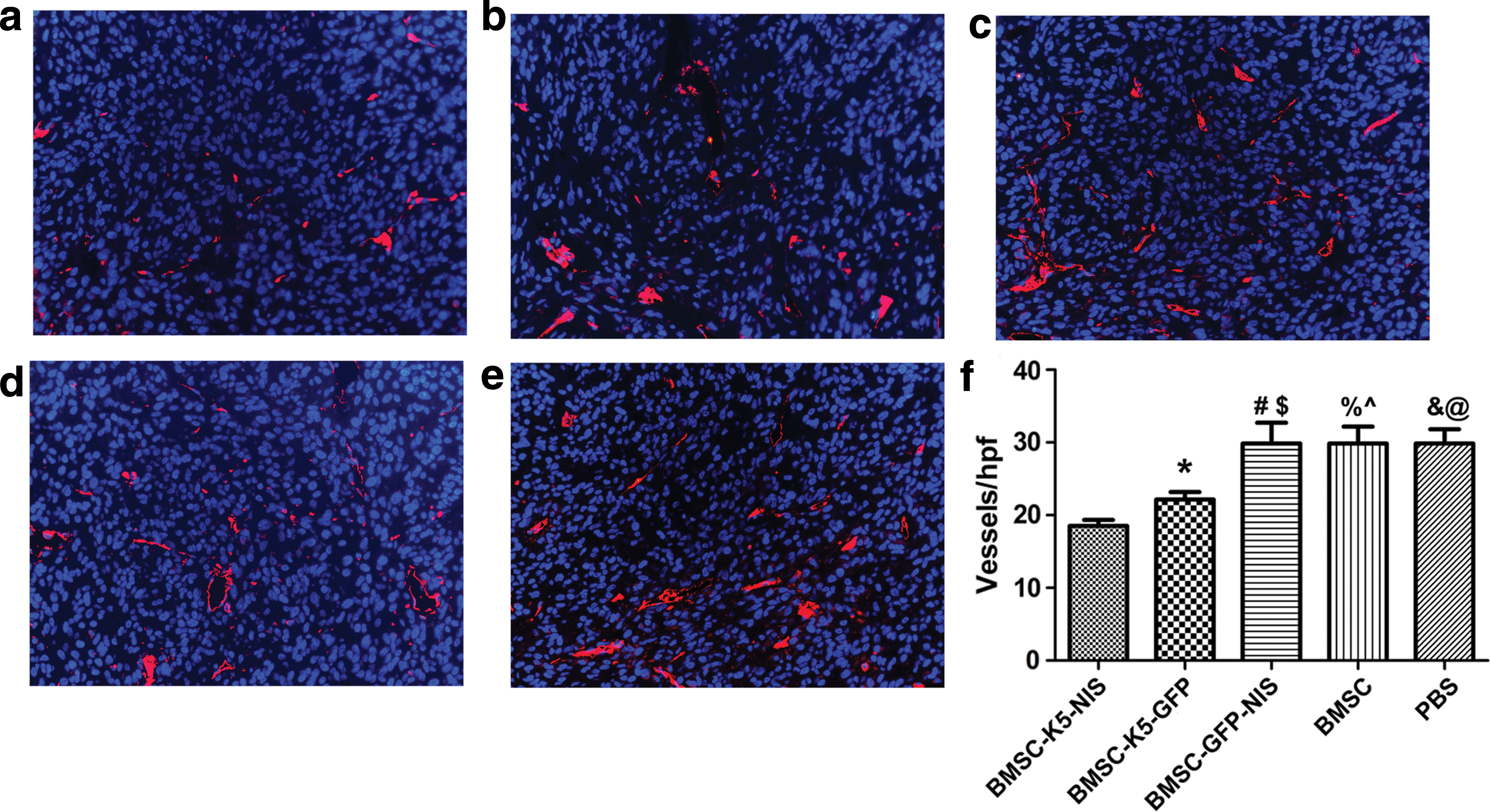
Immunofluorescence analysis.
Discussion
This study evaluated the role of engineered BMSCs as a therapeutic gene vehicle for the selective delivery of an antiangiogenesis gene combined with a gene involved in absorbing radionuclides in the context of the glioblastoma microenvironment. 125I micro-SPECT/CT imaging was used to monitor the migration of intravenous implanted BMSCs.
The ability of mesenchymal stem cells (MSCs) to migrate specifically to tumors has led to the use of these cells as a delivery vector for anticancer agents in different tumor models. They have been engineered to express and provide targeted delivery of immunomodulatory cytokine interleukin-12, 27 interferons (IFN) such as IFN-α and IFN-β, 28 –30 the prodrug converting enzyme such as herpes simplex virus thymidine kinase, 31 tumor necrosis factor apoptosis-inducing ligand, 32 or NIS 33 to many types of tumors. The systemic MSC-mediated NIS gene delivery followed by 131I application resulted in a significant delay in hepatocellular tumor growth. 33 In this study, K5 and NIS were chosen as the therapeutic agent because of their potent synergistic antitumor effect. In addition, NIS has been shown to be a safe and effective approach not only for the treatment of tumors but also for following the fate of therapeutic MSCs with a noninvasive imaging method.
The approach showed efficacy in an orthotopic glioblastoma tumor model. The antiangiogenesis K5 protein combined with 188Re uptake mediated by NIS gene inhibited tumor angiogenesis and growth prolonged the survival time of tumor-bearing nude mice. K5 is an antiangiogenesis factor that targets endothelial cells and is cytotoxic. The in vitro results showed that K5 inhibited migration of HUVEC cells and treatment study in vivo showed fewer CD31+ capillaries in the BMSC-K5-NIS group compared to the BMSC-K5-GFP (p < 0.05), BMSC-GFP-NIS (p < 0.01), BMSC (p < 0.01), and PBS (p < 0.001) groups. As angiogenesis is largely absent in healthy adult cells and present in solid tumors, especially in glioblastoma, which is one of the most vascularized cancers, 34 only the tumor environment is targeted using this approach. BMSCs can be obtained directly from glioblastoma patients. Therefore, this therapy would not only reduce the side effects of residual implanted BMSCs in the tumor after treatment, but also permit the specific delivery of the therapeutic gene through easy i.v. administration without the risk of allogeneic implantation rejection.
Several studies have reported that BMSCs could increase tumor growth and metastasis. 35 –37 However, these effects were not observed in this study. No difference in tumor growth were observed between the sham group (i.v. injected PBS) and BMSC-implanted group (without therapeutic gene). Bexell et al. 38 also reported that BMSC grafting had no effect on tumor microvessel density or survival of glioma-bearing animals. It is possible that the effects of BMSCs on tumor growth and metastasis in vivo are not universal in solid tumors, especially glioblastoma. In addition, no BMSC-GFP-NIS cells were observed in the tumor by 125I micro-SPECT/CT imaging or immunofluorescence 5 days after 188Re radiation therapy in this study, indicating that NIS may act as a safety mechanism in BMSC implantation. Bexell et al. 39 found no BMSCs in gliomas at 2 and 7 days following BMSCs injection. The authors suggested that BMSCs should be administered by intratumoral implantations rather than by i.v. injection. However, in this study, after three rounds of i.v. BMSC implantation, 125I micro-SPECT/CT imaging showed that there were BMSCs within the glioblastoma tumor on days 10 and 17 but not on day 24. The quantity of grafting BMSCs in the tumor tissue peaked on day 10, decreased by day 17, reaching the lowest levels on day 24, as determined by 125I micro-SPECT/CT imaging. The quantity of BMSCs in the lung also peaked on day 10, and then decreased almost to the same levels as the control group on day 17. These results demonstrate that BMSCs could be administered by i.v. injection. However, the migration time is longer than by intratumoral implantation, so 188Re or other therapies relying on BMSCs should be used only once the BMSCs have had sufficient time to migrate to the tumor. Furthermore, 188Re or other treatment drug may lead to severe lung injury on day 10 after BMSC i.v. implantation due to the high levels of BMSCs in lung tissue on at that time. Therefore, day 17 may be the most suitable time for treatment.
Using a glioma surgical treatment model, Bexell et al. 39 found that intratumorally grafted BMSCs could migrate efficiently within glioma remnants after partial surgical resection. Klopp et al. 9 also showed that tumor irradiation increased the recruitment of circulating MSCs into the tumor microenvironment. In glioblastoma clinical treatment, the majority of patients have undergone tumor resection and irradiation, and most of them experience high tumor recurrence. Therefore, BMSC-mediated gene therapy is more suitable for these patients.
Future studies should focus on BMSCs-mediated gene therapy for glioma remnants and the tumor after irradiation.
Footnotes
Acknowledgments
We are indebted to the staff of the Department of Nuclear Medicine, Fudan University Shanghai Cancer Center, for their technological support with micro-SPECT/CT imaging. This work was supported by grants from the National Natural Science Foundation of China (grant number 81471686) and the Discipline Leaders Climbing Project of Ruijin Hospital and Shanghai Health Bureau Youth Foundation (grant number 2010Y021).
Author Disclosure
The authors declare that they have no competing interests.