
Abstract
Myostatin, a negative modulator of muscle growth, has been considered as a potential target for the treatment of type 2 diabetes (T2D). In previous work, it was found that myostatin inhibition by adeno-associated virus (AAV)-mediated gene delivery of myostatin propeptide (MPRO) could improve muscle mass and achieve therapeutic effects on glucose regulation and lipid metabolism in db/db mice. This study investigated whether pre-intervention of rAAV-mediated expression of MPRO could lower the incidence of T2D. Three-week-old male C57BL/6 mice were randomly divided into saline control, rAAV-GFP, and rAAV-MPRO groups, all of which were fed on a high-fat diet. It was observed that pre-intervention of rAAV-MPRO prevented high-fat diet-induced hyperglycemia and hyperlipidemia. It also improved glucose tolerance, downregulated serum insulin levels, and facilitated the growth of skeletal muscle and fat redistribution, with no significant difference in serum free fatty acid levels and body weight, which ultimately reduced the incidence of T2D. In addition, pretreatment of rAAV-MPRO in C2C12 cells increased insulin-stimulated glucose uptake, as well as glycogen synthesis under insulin resistance conditions induced by free fatty acids, with no significant difference in insulin-stimulated glucose oxidation. Finally, the study demonstrated that improved glucose metabolism by rAAV-MPRO pretreatment might be due to the activation of the PI3K/Akt/GSK3β pathway and spurring Glut4 transposition from the cytoplasm to the cytomembrane in C2C12 cells. Based on these findings, MPRO is most likely to be a new method for the prevention of T2D.
Introduction
Type 2 diabetes (T2D) is a common and multifactorial metabolic disease worldwide with a gradually increasing mortality rate. 1 The main characteristic of this disease is disordered glucose homeostasis by pancreatic β cell insufficiency and insulin resistance. 2,3 Though previous studies have suggested some treatment methods for T2D, those approaches have been hampered by insulin resistance, which weakens the response to insulin signaling for glucose uptake and metabolism of its target tissues. 4,5 Therefore, enhancing insulin sensitivity can be an effective method to prevent T2D.
Myostatin, a member of the transforming growth factor beta (TGF-β) superfamily, is expressed mainly in skeletal muscle. 6 Its primary function is the negative regulation of muscle growth and development. 7 Evidence indicates that loss of function of myostatin by mutation can increase insulin sensitivity, and it has been well established that myostatin can regulate insulin sensitivity. 8 –10 Thus, inhibition of myostatin has been proposed as a therapeutic approach in T2D.
A previous study indicated that myostatin inhibition by adeno-associated virus (AAV)-mediated gene delivery of myostatin propeptide (MPRO) reversibly bound to myostatin and inhibited its activity. Further, it improved muscle mass and achieved therapeutic effects on glucose regulation and lipid metabolism in db/db mice. 11 Transgenic mice overexpressing MPRO were reported to be protected from high-fat diet-induced obesity by exhibiting lower blood glucose and insulin levels, increased muscle mass, and improved glucose tolerance. 10
Considering the potential effects of MPRO on the treatment of T2D, the current study further explored the physiological role of MPRO in vivo and in vitro. Here, the effect was investigated of pre-intervention of recombinant AAV (rAAV)-mediated expression of MPRO on the incidence of T2D in C57BL/6 mice fed a high-fat diet and on the C2C12 cells of insulin resistance induced by free fatty acids (FFA). The study sought to determine whether pretreatment of rAAV-mediated expression of MPRO could lower the incidence of T2D and the mechanisms involved in it.
Methods
Construction and preparation of rAAV vectors
AAV serotype 8 was used to construct a rAAV plasmid, rAAV-CAG-MPRO/Fc, which expresses MPRO protein. The promoter used in the constructed plasmid was specific to skeletal muscle and consisted of the chicken beta-actin promoter, cytomegalovirus enhancers, and a large synthetic intron, which is a robust transcription promoter in liver cells. Thus, MPRO was expressed mainly in skeletal muscle. The recombinant vectors containing MPRO were prepared by triple plasmid co-transfection in human embryonic kidney 293 cells, as described previously. 12,13 Plasmids for viral packaging were obtained from Dr. Xiao Xiao (University of North Carolina Eshelman School of Pharmacy, Chapel Hill, NC). 13 The construction method of the plasmid has been introduced previously, and MPRO expression in skeletal muscle cells was confirmed. 11 The titers of the purified viral stocks were determined by the DNA dot blot method, as reported previously. 14,15 The titer of the rAAV used in this study was about 1 × 1012 vg/mL. rAAV-GFP was used as a control vector.
Experimental animals
Three-week-old male C57BL/6 mice, purchased from the Experimental Animal Center of Changsha (Changsha, P.R. China), were used in the experiments. The study was approved by the Institutional Animal Research Committee of Tongji Medical College. All animal experimental protocols complied with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The mice were housed five per cage in a room with a 12 h light/dark cycle, a temperature of 20°C, and a humidity of 55 ± 5%. The animals had free access to normal mice chow and water throughout the experiment. Following 1 week of acclimation, the mice were randomized into three groups, feeding on a high-fat diet (D12492, 60% energy from fat; Beijing HFK Bio-Technology, Beijing, P.R. China), and treated with tail-vein injections of saline, rAAV-GFP, and rAAV-MPRO, respectively. Body weight, blood glucose, and other physical parameters were determined at the start of the experiment and every week up to 24 weeks. At sacrifice, mice were anesthetized with an intraperitoneal injection of 2.5% avertin (37.5 g/kg). Blood was drawn from the tail vein, and the skeletal muscles were removed. These samples were then stored at −80°C until analysis was performed.
Blood glucose and glucose tolerance test
Blood glucose level was tested at least twice a week from the snipped tail-vein blood at about 9:00am by a handheld glucometer (One-Touch; Lifescan, Inc., Milpitas, CA). A glucose tolerance test (GTT) was performed at 14 weeks after saline and rAAV vector injection. In this experiment, mice received an intraperitoneal injection of 10% dextrose (1 g/kg) after an overnight fast (about 8–12 h). Two drops of blood were obtained from the tail vein at the indicated time points (0, 30, 60, 90, and 120 min) after the dextrose injection to measure the blood glucose level using a One-Touch glucose monitor (Lifescan, Inc.). Both of the tests were expressed as the actual blood glucose value.
Skeletal muscle and fat pad weight analysis
The mice were sacrificed at the end of the experiment. Quadriceps, gastrocnemius, tibialis anterior, and triceps muscle were dissected and weighed. The subcutaneous, epididymal, and retroperitoneal adipose tissue fat pads were also dissected and weighed. The method was followed according to the procedures established by Johnson et al. 16
Serum insulin, FFA, total cholesterol, and triglyceride assay
Serum insulin, FFA, total cholesterol (TC), and triglycerides (TG) were measured using enzyme-linked immunosorbent assay kits from Zen-Bio, Inc. (Research Triangle Park, NC) and Crystal Chem (St. Louis, MO), as per the manufacturers' instructions.
Cell culture and treatments
C2C12 cells, a mouse myoblast cell line, were obtained from American Type Culture Collection (CRL-1446; ATCC, Manassas, VA) and cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and penicillin-streptomycin (100 IU/mL) in a humidified atmosphere of 95% air and 5% CO2 at 37°C.
Cells were planted in six-well plates, transfected with rAAV vectors, followed by FFA (0.4 mM) and/or insulin (100 nM) treatment, and further incubated for 24 h before being collected for Western blotting.
Glucose uptake analysis
The glucose uptake rate was assessed, as described previously. 17 Briefly, C2C12 cells were homogenized in 0.5% perchloric acid and centrifuged at 2000 g for 15 min at 4°C. The supernatants were neutralized with potassium hydroxide. An aliquot of the homogenate was quantified by liquid scintillation counting to determine total values for the sum of [ 14 C]2-deoxyglucose ([ 14 C]2DG) and [ 14 C]2-deoxyglucose phosphate ([ 14 C]2DGP). Another aliquot was deproteinized with zinc sulfate and barium hydroxide to precipitate [ 14 C]2-deoxyglucose-6-phosphate ([ 14 C]2DG6P) and quantify [ 14 C]2DG in the supernatant. The value for the [ 14 C]2DG in the supernatant was subtracted from the total [ 14 C]2DG and [ 14 C]2DGP to calculate the glucose uptake rate.
Glycogen assay
C2C12 cells were homogenized in 0.03 N HCl, mixed with 1.25 N HCl, and heated for 1 h at 100°C. Samples were centrifuged at 21756 × g, and 10 mL supernatant was mixed with 1 mL glucose oxidase reagent (Sigma–Aldrich, St. Louis, MO). After incubation at 37°C for 10 min, the absorbance was read at 505 nm. A standard curve was simultaneously analyzed using glycogen type III obtained from rabbit (Sigma–Aldrich) to determine the final glycogen concentrations.
CO2 production assay
CO2 production assay was carried out, as described previously. 18 Briefly, C2C12 cells were cultured in 12.5 cm 2 flasks, which were made airtight by using stopper tops. After 4 h, 300 μL phenylethylamine methanol was added with a syringe to a center well containing a folded filter paper. Subsequently, 300 μL 1 mol/L perchloric acid was added to the cells through the stopper tops using a syringe. The flasks were placed at room temperature for at least 1 h to trap the labeled CO2.
Cytosolic and plasma membrane fraction of C2C12 cell preparation
The plasma membrane fraction of C2C12 cells was prepared by differential ultracentrifugation, as described by Miura et al. 19 Briefly, the cells were placed in buffer (5 mM sodium azide, 0.25 M sucrose, 0.1 mM phenylmethylsulfonyl fluoride, 10 mM NaHCO3, pH 7.0) at 4°C, homogenized, and centrifuged at 1,000 g for 10 min, and the supernatant was saved as cytosolic fractions. The resulting pellet was re-suspended in the buffer and re-homogenized in a glass homogenization tube. The supernatant was then centrifuged at 190,000 g for 60 min. The pellet was collected as the plasma membrane pellet and re-suspended in buffer containing a protease inhibitor for Glut4 analysis.
Western blot analysis
Western blotting was performed, as described previously. 20 Briefly, both the membrane and cytosolic fractions were assayed for total protein using the Bradford method, as described by the manufacturer (Bio-Rad, Hercules, CA). About 20 μg protein was suspended in 1% sodium dodecyl sulfate and 50 mM dithiothreitol, subjected to electrophoresis, and transferred onto a polyvinylidene fluoride membrane (Bio-Rad, Burlington, Canada), which was incubated at room temperature for 2 h with a blocking solution, 5% nonfat milk, then probed with anti-total IR, anti-total IRS, anti-Glut4 (Santa Cruz Biotechnology, Sant Cruz, CA), anti-PI3K, anti-total Akt, and anti-total GSK3β and phosphor-specific antibodies to IR, IRS, Akt, and GSK3β (Cell Signaling Technology, Beverly, MA) overnight at 4°C. Next, these membranes were treated with a secondary antibody conjugated to horseradish peroxidase (Sigma–Aldrich, St. Louis, MO) at room temperature for about 2 h. After incubation with each antibody, the membranes were washed five times with TBST buffer containing 10 mM Tris-Cl (pH 7.5), 100 mM NaCl, and 0.1% Tween 20 at room temperature. Immunoreactive bands were examined with ECL solution (Thermo Fisher Scientific, Waltham, MA), quantified by densitometry, and normalized to their respective controls.
Statistical analysis
All in vitro experiments were repeated at least three times with the same intervention. Results are expressed as the mean ± standard error of the mean. Differences between groups were evaluated using an unpaired Student's t-test or one-way analysis of variance and Bonferroni post test. p-Values of <0.05 was considered statistically significant. SPSS 22.0 Statistics for Windows was used in this study.
Results
Pretreatment of rAAV-MPRO prevents hyperglycemia and the impairment of glucose tolerance, decreasing the incidence rate of T2D mellitus of mice on a high-fat diet
To determine whether rAAV-MPRO pretreatment could prevent hyperglycemia in mice on a high-fat diet, blood glucose was tested in different groups. It was found that the initial blood glucose concentration of the three groups did not show any difference before feeding on a high-fat diet (saline [n = 5], 5.03 ± 0.65 mmol/L; rAAV-GFP [n = 5], 5.10 ± 0.58 mmol/L; rAAV-MPRO [n = 5], 4.97 ± 0.48 mmol/L). Blood glucose levels of saline- and rAAV-GFP-injected mice increased gradually and remained at a high level (>7.0 mmol/L) during the study period. However, there was no increase above 7.0 mmol/L in the rAAV-MPRO pretreated group from the third month on a high-fat diet. The mean blood glucose values of rAAV-MPRO pretreated mice were significantly lower than those of the saline and rAAV-GFP pretreated groups from that time (p < 0.05; Fig. 1A).
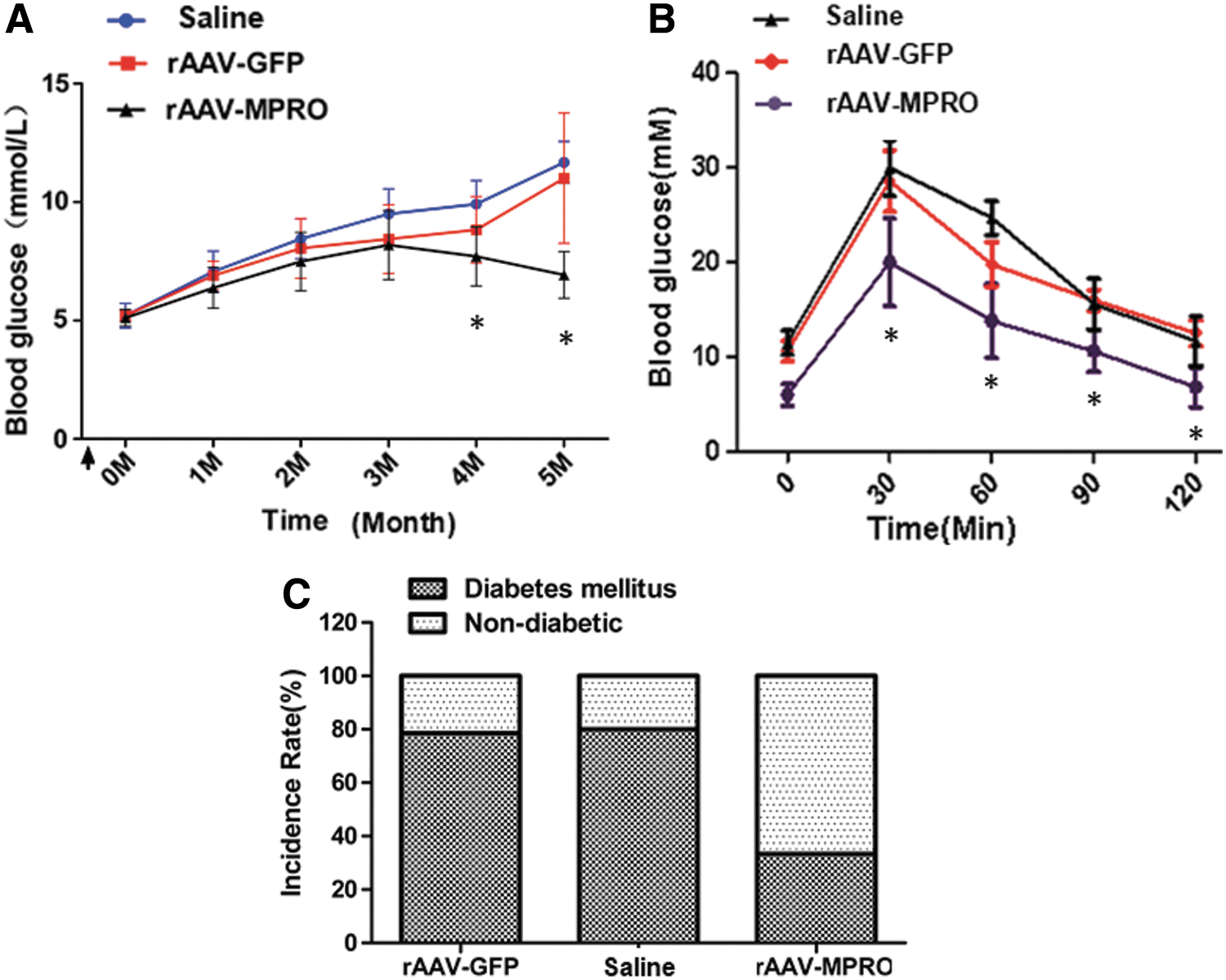
Pretreatment of recombinant adeno-associated virus (rAAV)-myostatin propeptide (MPRO) prevents the increment of blood glucose and the impairment of glucose tolerance, decreasing the incidence rate of type 2 diabetes (T2D) mellitus of mice on a high-fat diet.
Moreover, to assay the effect of increased MPRO expression on glucose tolerance, an intraperitoneal GTT was performed at 14 weeks after saline and rAAV vector injection. Saline and rAAV-GFP pretreated mice had significantly higher fasting blood glucose levels and an abnormal response to the glucose injection (Fig. 1B). However, rAAV-MPRO pretreatment resulted in an improvement in glucose tolerance compared to the other two groups. At 2 h after the glucose injection, the mean blood glucose levels in rAAV-MPRO pretreated mice decreased significantly to 9.02 ± 0.82 mmol/L, which was lower than those of the saline (13.4 ± 1.02 mmol/L) and rAAV-GFP pretreated animals (14.3 ± 0.83 mmol/L; p < 0.05; Fig. 1B).
Further, we calculated the incidence rate of T2D at 20 weeks after pretreatment. Mice with a high blood glucose level (>250 mg/dL) were diagnosed as T2D mice, and there were fewer suffering from T2D in the rAAV-MPRO pretreatment group than in the saline and rAAV-GFP groups (Fig. 1C).
These results indicate that pretreatment with rAAV-MPRO prevents hyperglycemia and the impairment of glucose tolerance, and further decreases the incidence of T2D mellitus of mice on a high-fat diet.
rAAV-MPRO pretreatment facilitates the growth of skeletal muscle and fat redistribution, with no significant difference in body weight of mice on a high-fat diet
Because most T2D patients tend to be obese, the body weight of mice in each group was evaluated, and no significant changes were found (Fig. 2A). To investigate the composition of body weight further, the main skeletal muscle and fat pad weights were compared. As shown in Fig. 2B, the gastrocnemius and tibialis anterior muscles in rAAV-MPRO pretreated mice were heavier than those in the other groups (p < 0.05). Meanwhile, the subcutaneous fat of rAAV-MPRO pretreated mice was lighter compared to mice pretreated with saline and rAAV-GFP (Fig. 2C and D). In addition, the epididymal fat of rAAV-MPRO pretreated mice was heavier than that of other two groups (Fig. 2D).
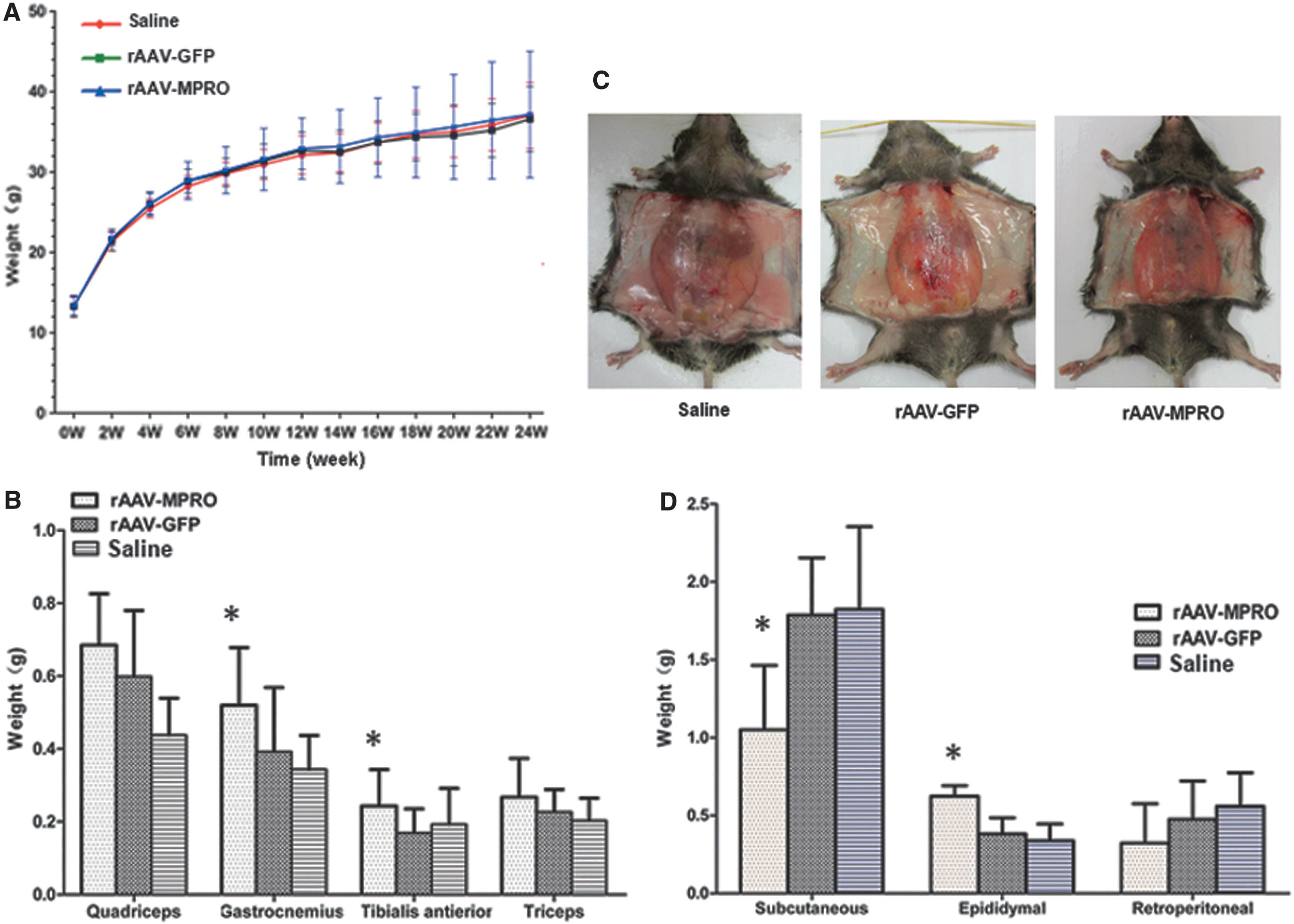
The effect of rAAV-MPRO pretreatment on body weight, different skeletal muscle mass, and fat redistribution in mice on a high-fat diet.
The above results suggest that rAAV-MPRO pretreatment facilitates the growth of skeletal muscle and fat redistribution, with no significant difference in body weight of mice on a high-fat diet.
Pretreatment of rAAV-MPRO prevents hyperlipidemia and hyperinsulinemia, with no significant difference in FFA in mice on a high-fat diet
Since no changes in body weight were observed, the serum lipid profile was analyzed further. The results showed that TG and TC of rAAV-MPRO pretreated mice were significantly reduced compared to the rAAV-GFP and saline pretreated groups (Fig. 3A and B), while the FFA content exhibited no significant changes (p < 0.05; Fig. 3C).
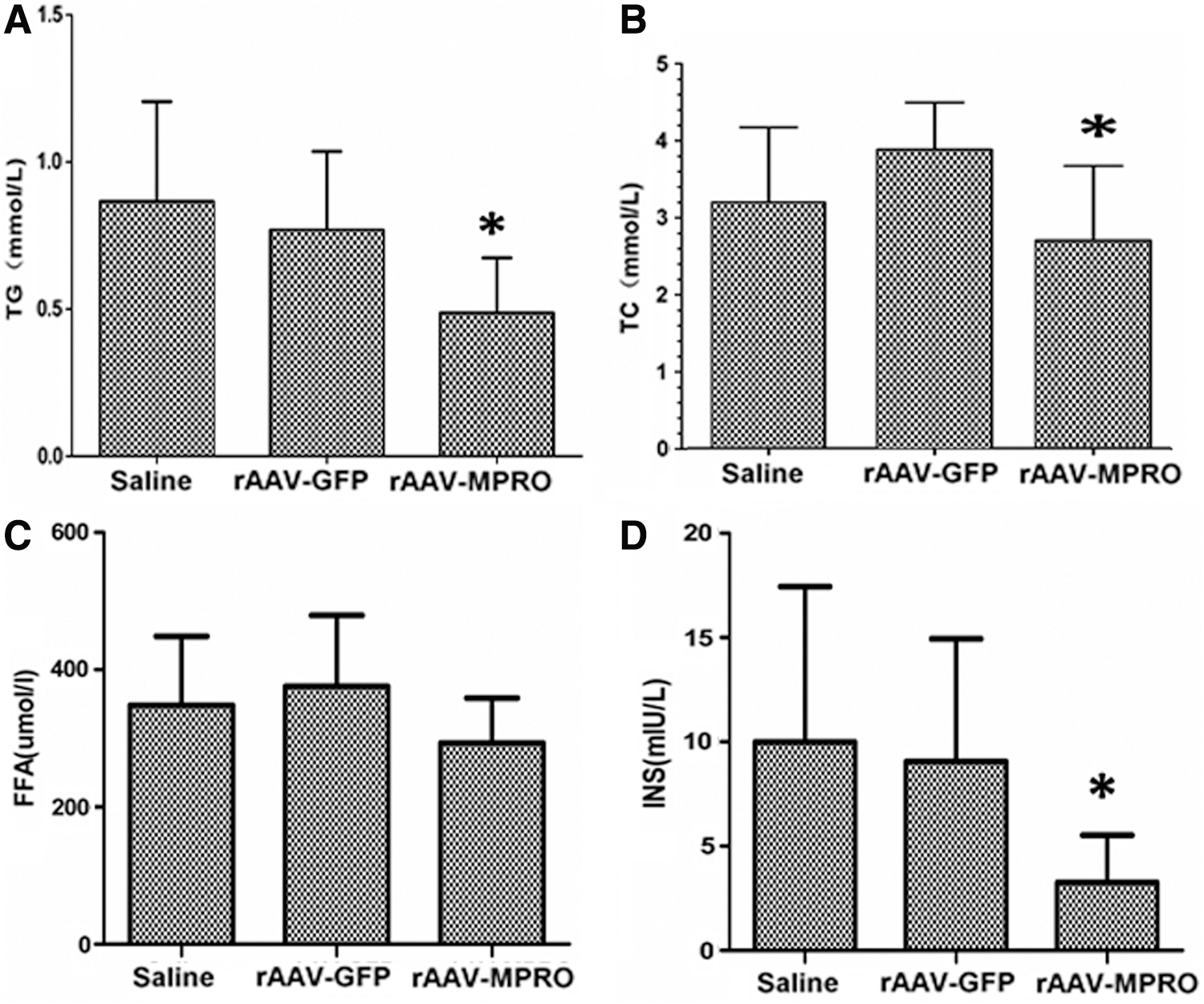
The effect of rAAV-MPRO pretreatment on blood lipoids and insulin levels in mice on a high-fat diet.
To determine whether rAAV-MPRO pretreatment would affect the insulin secretion of mice on a high-fat diet, serum insulin levels were measured at 20 weeks after pretreatment. It was observed that rAAV-MPRO pretreated mice displayed significantly lower insulin levels than those of saline and rAAV-GFP pretreated mice (p < 0.05; Fig. 3D).
These results indicate that pretreatment of rAAV-MPRO prevents hyperlipidemia and hyperinsulinemia, with no significant difference in free fatty acid in mice on a high-fat diet.
Pretreatment of rAAV-MPRO increases insulin sensitivity and facilitates glucose metabolism in C2C12 cells of insulin resistance induced by FFA
The study further investigated the impact of pretreatment of rAAV-MPRO on C2C12 cells. It was observed that pretreatment with rAAV-MPRO increased the uptake of 2-DG and facilitated the synthesis of glycogen in C2C12 cells of insulin resistance induced by FFA, with no significant difference in the aerobic oxidation of glucose (p < 0.05; Fig. 4A–C).

The effect of rAAV-MPRO on glucose metabolism in C2C12 cells of insulin resistance induced by FFA.
These data indicate that pretreatment with rAAV-MPRO increases insulin sensitivity and facilitates glucose metabolism in C2C12 cells of insulin resistance induced by FFA.
rAAV-MPRO activates IR/IRS phosphorylation and improves Glut4 translocation from the cytoplasm to the cytomembrane in C2C12 cells of insulin resistance induced by FFA
To analyze further the mechanism associated with rAAV-MPRO in ameliorating insulin resistance and the impairment of glucose tolerance, the insulin signaling status was examined in C2C12 cells of insulin resistance induced by FFA. It was found that IR and IRS phosphorylation was significantly increased in the rAAV-MPRO group compared to the saline and rAAV-GFP groups (p < 0.05; Fig. 5A–D). In addition, the alteration in insulin-stimulated Glut4 translocation in C2C12 cells of insulin resistance induced by FFA was tested by Western blot. It was found that insulin significantly increased the Glut4 translocation from the intercellular pool to the plasma membrane in the rAAV-MPRO group (p < 0.05; Fig. 5E and F).
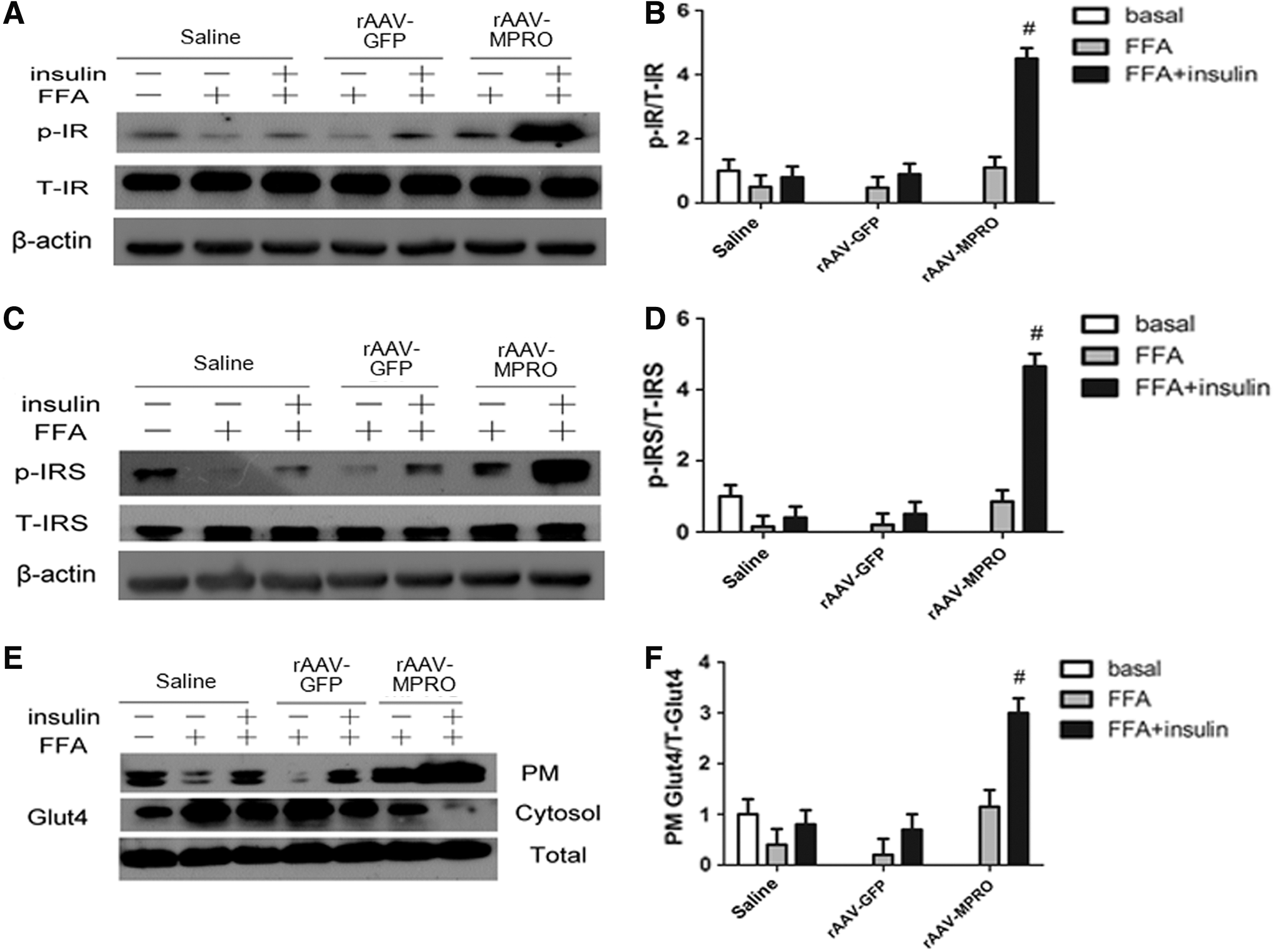
rAAV-MPRO activates IR/IRS phosphorylation and improves Glut4 translocation from the cytoplasm to the cytomembrane in C2C12 cells of insulin resistance induced by FFA.
rAAV-MPRO activates the PI3K/Akt/GSK3β pathway in C2C12 cells of insulin resistance induced by FFA
Next, the active status of the PI3K/Akt/GSK3β pathway was examined, which mediates many metabolic effects of insulin, including the activation of Glut4 translocation to the plasma membrane. 21 –23 Immunoblot analysis showed that PI3K and Akt/GSK3β phosphorylation was significantly increased in the rAAV-MPRO group compared to the saline and rAAV-GFP groups (p < 0.05; Fig. 6A–F).

rAAV-MPRO activates the PI3K/Akt/GSK3β pathway in C2C12 cells of insulin resistance induced by FFA.
These findings indicate that inhibition of myostatin with rAAV-MPRO might improve glucose metabolism through activating the PI3K/Akt/GSK3β pathway.
Discussion
A previous study found that AAV-mediated gene delivery of MPRO could improve muscle mass and achieved therapeutic effects on glucose regulation and lipid metabolism in db/db mice. 11 This study shows that pretreatment with rAAV-MPRO could attenuate hyperglycemia and the impairment of glucose tolerance, decreasing the incidence rate of T2D, facilitate the growth of skeletal muscle and fat redistribution, and improve insulin sensitivity. These results suggest that MPRO is likely to be a potential method for the prevention of T2D.
The in vivo effects of rAAV-MPRO pretreatment on blood glucose levels were examined in C57BL/6 mice. Results showed that attenuated hyperglycemia lasted up to 5 months while on a high-fat diet. It is widely known that diabetes is a lifelong disease, and long-term blood glucose control is necessary to prevent chronic complications. 24 Thus, rAAV-MPRO pretreatment could be an effective prevention tool for T2D. Interestingly, mice receiving rAAV-MPRO pretreatment exhibited a decrease in fasting blood glucose and had sharply reduced insulin levels, while the saline and rAAV-GFP pretreated groups had significantly higher fasting blood levels and an abnormal response to the glucose injection. Furthermore, the rAAV-MPRO group showed improved glucose tolerance and insulin resistance. All these results indicate that decreased insulin sensitivity rather than decreased insulin secretion contributes to the impairment of glucose tolerance and hyperglycemia. Hyperglycemia itself can cause insulin resistance. 25 Thus, correction of blood glucose levels should have positive effects on human insulin sensitivity.
Additionally, no significant effect of rAAV-MPRO pretreatment on body weight and FFA was observed in mice on a high-fat diet. This conclusion is not consistent with previous reports that myostatin knockout or MPRO therapy can increase body weight and decrease FFA. 26,27 The causes of these results have not yet been entirely understood, although it is probably because of the different genetic backgrounds or the different timings for giving the MPRO intervention. 28,29 Previous studies have indicated that MPRO may affect food uptake, body weight, and fat metabolism by regulating the concentration of leptin in serum. 10,26 This study showed that rAAV-MPRO pretreatment increased skeletal muscle mass and facilitated fat redistribution. These findings further support the notion that inhibition of myostatin is sufficient to trigger the growth of skeletal muscle. 10,26,27,30,31 A large proportion of insulin stimulating glucose uptake accounts for skeletal muscle, which represents the most important tissue for the maintenance of a balanced postprandial glucose homeostasis. 32 Therefore, the hypertrophy muscles would demand and utilize more energy nutrients, including glucose, and skeletal muscle would become a dominant storage site for glucose utilization. As for the fat redistribution in rAAV-MPRO pretreated mice, this might result from the different function of tissues and the different roles of fat played in these tissues.
To investigate the mechanism through which pretreatment of rAAV-MPRO enhances insulin sensitivity, GLUT4 and the representative insulin signaling pathway were examined in C2C12 cells. The acute action of insulin leads to the stimulation of glucose uptake and glycogen synthesis. 33,34 Insulin exerts its biological effects through sequential activation of a cascade of signaling molecules. 35 Insulin binds to its receptor, which in turn results in the receptor auto-phosphorylation and the activation of several docking proteins, such as IRS. Phosphorylation of IRS allows activation of PI3K, which subsequently recruits the serine-threonine kinase, Akt, to the membrane to initiate activation of Akt. 36 –39 Then Akt activation results in the phosphorylation of serine-threonine kinase, GSK-3β, further leading to glucose uptake and Glut4 transposition from the cytoplasm to cell surface membranes. PI3K/Akt/GSK-3β activation is a required and important signaling pathway in glycometabolism. 21 –23 Interestingly, it was found that phosphorylation of Akt/GSK-3β and translocation of Glut4 were significantly increased after insulin stimulation in rAAV-MPRO pretreated C2C12 cells of FFA-induced insulin resistance, and 2-DG uptake and glycogen content were also increased in this group. These results suggested that increased glucose transport, increased glycogen synthesis, and lowered blood glucose—all of which were induced by increased insulin sensitivity—ultimately led to increased glycogen accumulation. This observation, together with others from previous studies, shows that MPRO could increase glucose oxidation in vitro and in vivo, 26 providing strong evidence that MPRO plays a vital role in glucose metabolism in muscle tissues.
In conclusion, the present study indicates that pre-intervention of rAAV-mediated expression of MPRO could attenuate hyperglycemia and the impairment of glucose tolerance, lowering the incidence of T2D, facilitate the growth of skeletal muscle and fat redistribution, and improve insulin sensitivity. MPRO may function through the activation of the PI3K/Akt/GSK3β pathway and spurring Glut4 transposition. These findings may provide important information regarding the potential clinical applications of MPRO pretreatment to prevent T2D.
Footnotes
Acknowledgments
This work was supported by the National Nature Science Foundation Committee of China (grant no. 30900694) and the Project Sponsored by the Scientific Research Foundation for the Returned Overseas Chinese Scholars, State Education Ministry (grant no. JYBHG201005).
Author Disclosure
No competing financial interests exist.