
Abstract
Recent marketing approval for genetically engineered hematopoietic stem and T cells bears witness to the substantial improvements in lentiviral vectors over the last two decades, but evaluations of the long-term efficacy and toxicity of gene and cell therapy products will, nevertheless, require further studies in nonhuman primate models. Macaca fascicularis monkeys from Mauritius have a low genetic diversity and are particularly useful for reproducible drug testing. In particular, they have a genetically homogeneous class I major histocompatibility complex system that probably mitigates the variability of the response to simian immunodeficiency virus infection. However, the transduction of simian cells with human immunodeficiency virus type 1 (HIV-1)-derived vectors is inefficient due to capsid-specific restriction factors, such as the tripartite motif-containing protein tripartite motif 5α, which prevent infection with non-host-adapted retroviruses. This study introduced the modified capsid of the macaque-trophic HIV-1 clone MN4/LSQD into the packaging system and compared transduction efficiencies between hematopoietic cells transduced with this construct and cells transduced with HIV-1 NL4-3-derived packaging constructs. Capsid modification increased transduction efficiency in all hematopoietic cells tested (by factors of up to 10), including hematopoietic progenitor cells, repopulating cells, and T cells from Mauritian Macaca fascicularis, regardless of vector structure or purification method. The study also established culture conditions similar to those used in clinical practice for the efficient transduction of hematopoietic stem and progenitor CD34+ cells. These results suggest that the procedure is suitable for use in Mauritian Macaca fascicularis, which can therefore be used as a model in preclinical studies for hematopoietic gene and cell therapy.
Introduction
Significant progress in the efficacy and safety of gene transfer have made it possible to use human immunodeficiency virus type 1 (HIV-1)-based lentiviral vectors (H-LVs) to treat patients with inherited 1,2 or acquired 3,4 diseases by ex vivo hematopoietic gene transfer. A few cell therapy products have entered Phase III pre-drug marketing clinical trials 5 or have recently been approved. 6 Nevertheless, a number of challenges remain, including the limited rate of human stem-cell transduction, the risk of insertional mutagenesis, and the possible loss of therapeutic benefit over time. Several strategies for improving the transduction efficiency of hematopoietic stem cells (HSCs) and lowering the risk of long-term aplasia have been developed. They include the optimization of culture media and cytokine combinations, the use of transduction enhancers acting on cellular entry or post-entry steps, the selection of transduced cells either ex vivo or in vivo, and the expansion of hematopoietic stem/progenitor cell (HSPC) populations. Important discoveries for both gene therapy and the immunotherapy of cancer and infectious diseases have been made in studies of immunodeficient mice, which are widely used as surrogate recipients for evaluating therapeutic approaches in human HSC and T-cell therapies. 7 However, despite the quality of the human hematopoietic myelolymphoid xenografts in these mouse models, several limitations render these mice unusable for long-term hematopoietic reconstitution studies in the context of humanized and autologous microenvironments, 8 and inappropriate for investigations of the pharmacological safety of genetically modified cells. 9 Furthermore, limited immune responses, a lack of lymphoid structures, and graft-versus-host responses 10 preclude non-clinical safety evaluations of cell and gene therapy products involving assessments of immune-mediated adverse events. 11
Nonhuman primates (NHPs) have similar HSC dynamics and engraftment properties to humans 12,13 ; they also have a similar immune system and display extensive physiological similarities. 14 However, gene transfer with H-LVs is hampered by low transduction efficiencies in Macaca fascicularis (cynomolgus macaques [CM]) and Macaca mulatta (rhesus monkeys [RM]), the two NHP species most frequently used for biomedical research in the United States and Europe. 15 –18 CMs from Mauritius (MCMs) originate from a small number of founders, 19 and their low genetic diversity 20 makes these animals particularly useful as a model for infectious diseases. Nevertheless, restriction factor polymorphisms 21,22 may act as confounding factors in gene therapy studies assessing the efficiency and toxicity of H-LV-mediated gene transfer in this subspecies.
The virus capsid (CA) plays an essential role in vector transduction by interacting with both factors supporting effective transduction 23 and the restriction factors produced by the cells to disrupt the retroviral life cycle. 24 The NHP innate immune factor tripartite motif (TRIM)5α protein and the TRIM5/cyclophilin A fusion protein (TRIMCyp) act as major restriction factors during the early post-entry step of HIV-1 infection. They restrict retroviral infection in a species-specific manner, 25 and differently between individuals within species, including RMs 26 –29 and CMs. 21,30 –32 Exon eight of the TRIM5α gene variant 33 is highly polymorphic and encodes the SPRY restriction domain, which binds to the capsid lattice, causing the species-specific restriction effect of TRIM5α. 34 –36 The TRIMCyp gene variant is translated into a protein in which the SPRY domain of TRIM5α is replaced by cyclophilin A, 37 –41 which interacts with the retroviral capsid 38,42,43 and acts like TRIM5α to promote HIV-1 restriction. 44,45
TRIM5 polymorphism and HIV-1 replication have been shown to be correlated in RM 25,38 and, more recently, CM cells. 21,22,30 –32,40,41,46 –48 As in RMs, 49 the allelic diversity of TRIM5α and TRIMCyp in CMs depends on geographic origin, 21,30 –32,47 and influences interindividual susceptibility to HIV-1. 22,31,40,41 The molecular clone SIVmac239 is resistant to all TRIM5 alleles and replicates in macaques. The replacement of wild-type CA with SIVmac239 CA in the context of chimeric HIV-1 50 or chimeric H-LV 51 –53 confers TRIM5 resistance. Capsid mutations leading to the substitution of one or a few amino acids 54 –62 have been shown to reduce restriction in RM cell lines and primary CD34+ cells. Amino-acid substitutions have also been generated in CM cell lines, 58,63,64 but the chimeric capsids were studied in the context of viral replication and not within a viral vector and in cell lines only. In this study, amino-acid modifications similar to those made in the capsid of the macaque-tropic HIV-1 clone MN4/LSQD 56,57 were introduced into H-LVs, and the transduction efficiency for this construct in hematopoietic cells from CM originating from Mauritius was compared to that achieved in the same cells with a wild-type CA.
Methods
Constructs, lentivector production, concentration, and purification
The chimeric (H/S) CA sequence (Supplementary Fig. S1) was synthesized by Genscript (Piscataway, NJ) and introduced into the HPV275 plasmid encoding GAG-POL 65 to yield HPV275-CAH/S (GAG-POL with the MN4/LSQD HIV-1-derived chimeric CA). The green fluorescent protein (GFP)-expressing LVs HPV57065 and LV3.4-BC 66 (Supplementary Fig. S2) were produced with a plasmid packaging system 65 consisting of HPV275 (gag-pol), ΨN15 (vsvG env), p633 (rev), and HPV601 (tat) to yield the wild-type capsid-containing vectors HPV570H-CA and LV3.4-BC1H-CA. HPV275 was replaced with HPV275-CAH/S to generate the H/S CA-containing vectors HPV570H/S-CA and LV3.4-BCH/S-CA. Vectors were either precipitated overnight at 4°C with Lentivirus Precipitation Solution (ALSTEM, Richmond, CA) or concentrated by ultracentrifugation. Where indicated, purification was performed with a Mustang Q anion exchange membrane with an Acrodisc unit (Pall, Saint Germain-en-Laye, France) connected to the AKTA PrimePlus system (GE Healthcare, Chicago, IL), under the control of PrimeView software (GE Healthcare). Desalting was performed with a 40K ZebaSpin desalting column (Thermo Fisher Scientific, Waltham, MA) before concentration. Infectious titers were determined in NIH3T3 cells, as previously described. 67
A sequence encoding an MCM TRIM5α protein with the variable amino acids most frequently identified in the animals (E4, H178, E209, D247, M330, Y387, and I435), established using the TRIM5α exon sequences of monkey C (see the Results section) was synthesized by Genscript (Supplementary Fig. S3), introduced into an orange fluorescent protein (OFP)-expressing vector, pLTGC/IRES/OFP (Supplementary Fig. S2), and sequenced. The resulting constructs, pLTGC/TRIM5α/IRES/OFP (Supplementary Fig. S2) and pLTGC/IRES/OFP, were used to generate wild-type capsid-containing recombinant lentiviruses LV-TRIM/I/O and LV-Mock/I/O, respectively.
Ethics statement
Adult male cynomolgus macaques (Macaca fascicularis) imported from Mauritius were housed in the CEA animal facilities (Fontenay aux Roses, France). They were handled in accordance with national regulations (permit number D92-032-02) and the standards of the Office for Laboratory Animal Welfare (OLAW) under insurance number A5826-01 and European directive 2010/63. Interventions were performed by veterinary surgeons and staff from the animal core facility on animals sedated with ketamine hydrochloride. This project received government authorization under agreement number 05-025.
NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ (NOD-scid IL2rγnull ; NSG) mice were obtained from Charles River Laboratories (Saint Germain sur l'Arbresle, France), kept in ventilated cages, and supplied with sterile food and autoclaved water. Animal care and handling conformed to EU Directive 2010/63/EU concerning the protection of animals used for scientific purposes. Experimental protocols were approved by the ethics committee for animal experimentation of the CEA under notification number 12-033.
NHP CD34+ cell isolation, culture, and transduction
Bone-marrow cells were harvested from the head of the humerus, the femurs, and the iliac crest (in the absence of mobilization). Cells were diluted 1:5 in Iscove's Modified Dulbecco's Medium (GE Healthcare), and separated by centrifugation on Ficoll-Paque Plus (GE Healthcare) containing 5% phosphate-buffered saline (PBS) at 600 g for 40 min. Mononuclear cells were then washed in 2% fetal bovine serum (FBS) and 2 mM ethylenediaminetetraacetic acid in PBS, and CD34+ cells were sorted with a Dynabeads CD34-positive isolation kit (Thermo Fisher Scientific), as previously described. 68
Simian CD34+ cells were cultured in StemMACS HSC Expansion Medium (Miltenyi Biotec, Bergisch Gladbach, Germany), X-VIVO 20 (Lonza, Basel, Switzerland), or CellGro SCGM (CellGenix, Freiburg, Germany) in the presence of recombinant human stem cell factor (100 μg/mL), FMS-like tyrosine kinase 3 ligand (100 μg/mL), interleukin-3 (60 μg/mL), and thrombopoietin (100 μg/mL). All cytokines were obtained from Miltenyi Biotech. For cell concentration studies, cells were plated at the indicated concentration and re-plated at the same concentration every 2–3 days. For transduction, cells were grown in the same medium with the same cytokines and cell concentration for 2 days, and then transduced at a concentration of 2 million cells/mL in the presence of 4 μg/mL protamine sulfate (Sigma–Aldrich, St. Louis, MO) for 24 h. The cells were then diluted at the indicated concentration and re-plated every 2–3 days. Where indicated, 1,000 cells were plated in H4434 methylcellulose-based medium already supplemented with cytokines (STEMCELL Technologies, Vancouver, Canada).
T-cell isolation, activation, and transduction
Peripheral blood mononuclear cells were isolated in Ficoll-Paque Plus (containing 4% PBS) and activated with the T-cell activation/expansion kit for NHP from Miltenyi Biotec, according to the manufacturer's instructions, in X-VIVO 15 (Lonza) containing 10% fetal calf serum (GE Healthcare). After 2 days of activation, 100 IU rhIL2 (Roche, Basel, Switzerland) per milliliter was added, and the cells were transduced in the presence of 4 μg/mL protamine sulfate and expanded. T-cell activation was verified by flow cytometry with antibodies directed against CD4, CD8, CD25, and CD69. The transduction rate (GFP+ cells) was determined 2 days after cell transduction.
Cell lines
Human HEK293T (ATCC CRL-11268) and HeLa (ATCC CCL-2) cells were maintained in complete Dulbecco's modified Eagle's medium (high glucose) supplemented with 10% FBS, 2 mM L-glutamine (GLN), 100 IU/mL penicillin, and 100 μg/mL streptomycin. All components were from GE Healthcare. Cynomolgus CYNOM-K1 (ECACC 90071809) and rhesus LLCMK2 (ECACC 85062804) cells were maintained in Eagle's minimum essential medium (EMEM) supplemented with 2 mM GLN and 1% non-essential amino acids (NEAA) in the presence of 10% and 5% FBS, respectively. EMEM and NEAA were obtained from Sigma–Aldrich.
Determination of transduction rate
As the vectors used in this study encode the GFP reporter gene, transduction rates were evaluated by measuring the percentage of GFP+ cells by flow cytometry or microscopy. Transduction levels were also evaluated by determining the mean vector copy number (VCN) in cells harvested from methylcellulose. Genomic DNA was extracted with the NucleoSpin Blood kit (Macherey-Nagel, Düren, Germnay), and real-time polymerase chain reaction (PCR) was performed with specific primers and probes using the 7300 ABI Prism Detection system and a 2 × qPCR MasterMix containing Rox (Eurogentec, Liège, Belgium). The primers and probes (Eurogentec) used were directed against the LVs (forward: 5′-TGTGTGCCCGTCTGTTGTGT-3′; reverse: 5′-CGAGTCCTGCGTCGAGAGA-3′; probe: 5′-FAM-CAGTGGCGCCCGAACAGGGA-3′-BHQ1) and a sequence of the GAPDH gene common to human and simian cells (forward: 5′-GTATCGTGGAAGGACTCATGACC-3′; reverse: 5′-GTTCAGCTCAGGGATGACCTTG-3′; probe: 5′-Yakima Yellow-AGTCCACGCCATCACTGCCACCCA-3′-BHQ1) for normalization. Mean VCN were determined in comparison to the results obtained with a human cell line containing one copy of the integrated vector per haploid genome. 69
Expected transduction rate in unbiased conditions
If cells are transduced at a known mean VCN, if they are equally transducible, and if integration events occur independently of each other, the expected percentage of transduced cells is based on Poisson distribution and is: (1 – e−VCN) × 100. Thus, if the actual percentage (% GFP+) and expected percentages ([1 – e−VCN] × 100) of transduced cells are similar, the cells within a sample can be considered to be equally transducible. 69
Sequencing of TRIM5 exons
PCR amplification was performed in reaction mixtures containing DNA templates extracted from cultured cells and primers described elsewhere. 30 The PCR products were sequenced in both directions by Genewiz (South Plainfield, NJ), and variant nucleotides were analyzed with Chromas.
Simian cell infusion in immunodeficient mice
Two- to four-month-old female NSG mice were subjected to whole-body irradiation with a sublethal dose of 2.8 Gy (1 Gy/min) or treated with 30 mg/kg intraperitoneally injected busulfan (Busilvex; 6 mg/mL; Pierre Fabre, Paris, France). The following day, freshly isolated or cultured and transduced simian CD34+ cells were injected intravenously into the mice via the retro-orbital plexus route. Blood samples were obtained from the tail, and bone-marrow hematopoietic cells were harvested from the two femurs and tibias after the mice had been killed. Cells were analyzed by flow cytometry with a MACSQuant Analyzer flow cytometer (Miltenyi Biotech) and FlowJo cell analysis software (LLC).
Antibodies for flow cytometry
The antibodies for flow cytometry are detailed in Table 1.
Antibodies for flow cytometry
Statistics
Comparisons between two or more groups were performed with Student's t-test or two-way analysis of variance. p-Values of <0.05, <0.01, and <0.005 were deemed to be of statistical significance. Linear regression was performed, and slopes were compared with GraphPad Prism v6 (GraphPad Software, Inc., La Jolla, CA). The error bars indicate the standard deviation.
Results
Study animals reproduce TRIM5α diversity observed in MCMs
The CMs housed at the authors' research center (IDMIT, Fontenay aux Roses, France) are imported from Mauritius. A PCR-based strategy 30,46,70 was used to check that they had no CypA insertion 3′ to TRIM5α exon 8, as previously reported for animals originating from Mauritius. 21,22 No PCR product larger than the expected size for the TRIM5α PCR product was detected in the six animals tested (Supplementary Fig. S4A). Then, the intron 6/exon 7 junction was sequenced, nucleotide polymorphisms of which have been shown to segregate with CypA insertion in both CMs and RMs. 21,22 Alleles from all animals displayed the canonical AG splice acceptor dinucleotide and the three associated nucleotide polymorphisms previously identified around the intron 6/exon 7 junction of TRIM5α but not of TRIMCyp (Supplementary Fig. S4B). Finally, the deduced amino-acid sequences encoded by exons 7 and 8 were found to be identical to those of TRIM5α 32 (Supplementary Fig. S4C). These results confirm previous observations 21,22 and attest to the absence of the TRIMCyp variant in MCMs housed at the research center.
In the six animals for which TRIM5α exons were amplified (Fig. 1A) and sequenced, seven divergent amino acids were observed (Fig. 1B) previously identified as polymorphic in CM, including two in the coiled-coil domain (H178K, E209K) and three in the B30.2/SPRY region (M330V, C387Y, and I435V). Two research groups investigating TRIM5 polymorphism in MCMs have published sequences for TRIM5α exon 8. 21,22 They observed divergence at three positions of the B30.2/SPRY domain (M330V, C387Y, and I435V) and distinguished three haplotypes on the basis of the amino-acid combinations present: Mafa4 (M330Y387I435), Mafa8 (V330C387I435), and Mafa9 (M330Y387V435), corresponding to 74.5%, 12.7%, and 12.7% of the chromosomes, respectively. The genotype combinations for the six study animals were determined: Mafa4/Mafa4 for monkeys B, C, M, and Z; Mafa8/Mafa9 for monkey A; and Mafa4/Mafa8 for monkey Y (Fig. 1C). The Mafa4, Mafa8, and Mafa9 alleles were therefore detected on 75%, 17%, and 8% of the chromosomes, suggesting that the animals housed in the animal center were representative of MCM TRIM5α diversity in Mauritius. 21,22

Macaca fascicularis monkeys at the authors' research center are representative of the TRIM5α diversity observed in Mauritius.
Cell concentration affects MCM CD34+ cell growth and transduction efficiency
This study investigated whether the serum-free medium, CellGrow SCGM (CG) or X-VIVO 20 (X20), used for the transduction of human CD34+ cells in gene therapy trials for hemoglobin disorders 1,71 was compatible with MCM CD34+ cell growth. Simian hematopoietic CD34+ cells could not survive in X20 (data not shown). Then, the effect of CG with another serum-free medium, StemMACS HSC (SM), was compared. The expected numbers of myeloid and erythroid progenitors were observed on day 3 with both media, with only minor differences between them, whereas the proportion of myeloid progenitors on day 7 was significantly higher in CG (Supplementary Fig. S5). It is concluded that CG medium supplemented with a cytokine cocktail previously used in a Phase I/II lentiviral/CD34+ clinical trial is compatible with the growth of bone-marrow CD34+ cells from MCMs.
An automated strategy for decreasing the levels of inhibitory paracrine signaling factors via a fed-batch medium dilution strategy has been shown to increase the abundance of hematopoietic cord blood cells with and without repopulating activity. 72 This study investigated whether a straightforward limitation of cell concentration during cell culture had a similar effect on the number of proliferating simian hematopoietic cells (Fig. 2 and Supplementary Fig. S6). Bone-marrow CD34+ cells were plated at different concentrations, and cell numbers were determined. For cells plated and transferred to fresh medium at a concentration ≤0.5 × 106 cells/mL, cell numbers 6 days after plating were five to seven times higher than those after plating and transfer at a concentration of 1 × 106/mL (Supplementary Fig. S6A). The effect on early progenitor cells was visible as soon as 2 days after plating (Supplementary Fig. S6B). It was stronger on day 6 (Supplementary Fig. S6C) and less marked thereafter (Supplementary Fig. S6D).
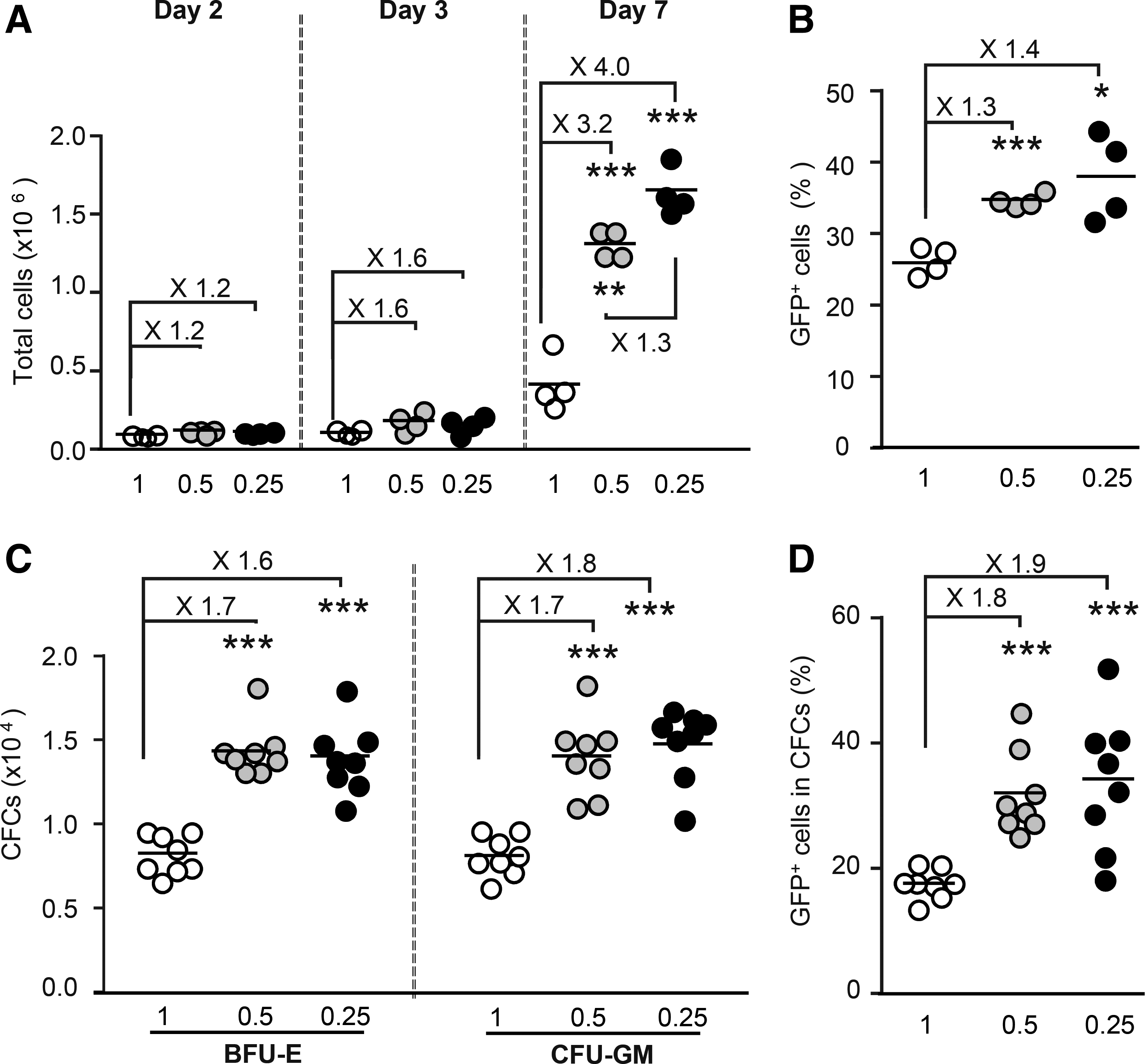
Cell concentration <0.5 × 106 cells/mL stimulates cell proliferation and lentiviral transduction. CD34+ cells were grown in liquid culture at a density of 0.25 × 106 cells/mL (n = 4), 0.5 × 106 cells/mL (n = 4), or 1.0 × 106 cells/mL (n = 4). Two days later (day 2), they were counted and transduced at a density of 2 × 106 cells/mL with unpurified HPV570H-CA. On day 3, a few cells were plated in methylcellulose medium (two wells per sample).
As the proliferation status of human cells affects the efficiency of transduction with LVs, 73,74 this study investigated whether cell concentration also affected the transduction of simian hematopoietic cells. CD34+ cells were plated at several concentrations in CG medium, and they were transduced 2 days later at a fixed concentration of 2 × 106 cells/mL with HPV570H-CA. The cells were re-plated at their initial concentrations on day 3. Total cell and progenitor cell numbers were greater and transduction rates were 1.5–2.0 times higher when cells were grown at a concentration of ≤0.5 × 106/mL (Fig. 2).
Therefore, in subsequent experiments, cells were plated at a concentration of 0.5 × 106/mL in CG medium and transduced at a concentration of 2 × 106 cells/mL.
A HIV/SIV chimeric capsid increases LV titer in NIH3T3 cells
HIV/SIV chimeric capsids (H/S-CAs), with substitutions of various lengths, ranging from single amino acids to the entire CA, were evaluated to identify the residues involved in TRIM5-mediated HIV-1 restriction in simian cells and/or to increase the sensitivity of monkey cells to HIV-1-derived vectors. However, the impacts of H/S-CAs on the transduction of hematopoietic CD34+ and T-lymphocyte cells have not been evaluated simultaneously in any species, or in hematopoietic CD34+ or T lymphocytes from MCM. Based on studies identifying the capsid surface amino acids involved in TRIM5-mediated anti-HIV1 restriction factors in macaques, the following amino acids from SIVmac239 were incorporated into the HIVNL4.3 CA (Fig. 3A): (1) the QPAPQQ motif between α-helixes 4 and 5, 54,55,57,58,63,64 (2) the RQQN stretch between α-helixes 6 and 7, 57,63 and the LSDQ amino acids from α-helixes 5 and 6. 57,58
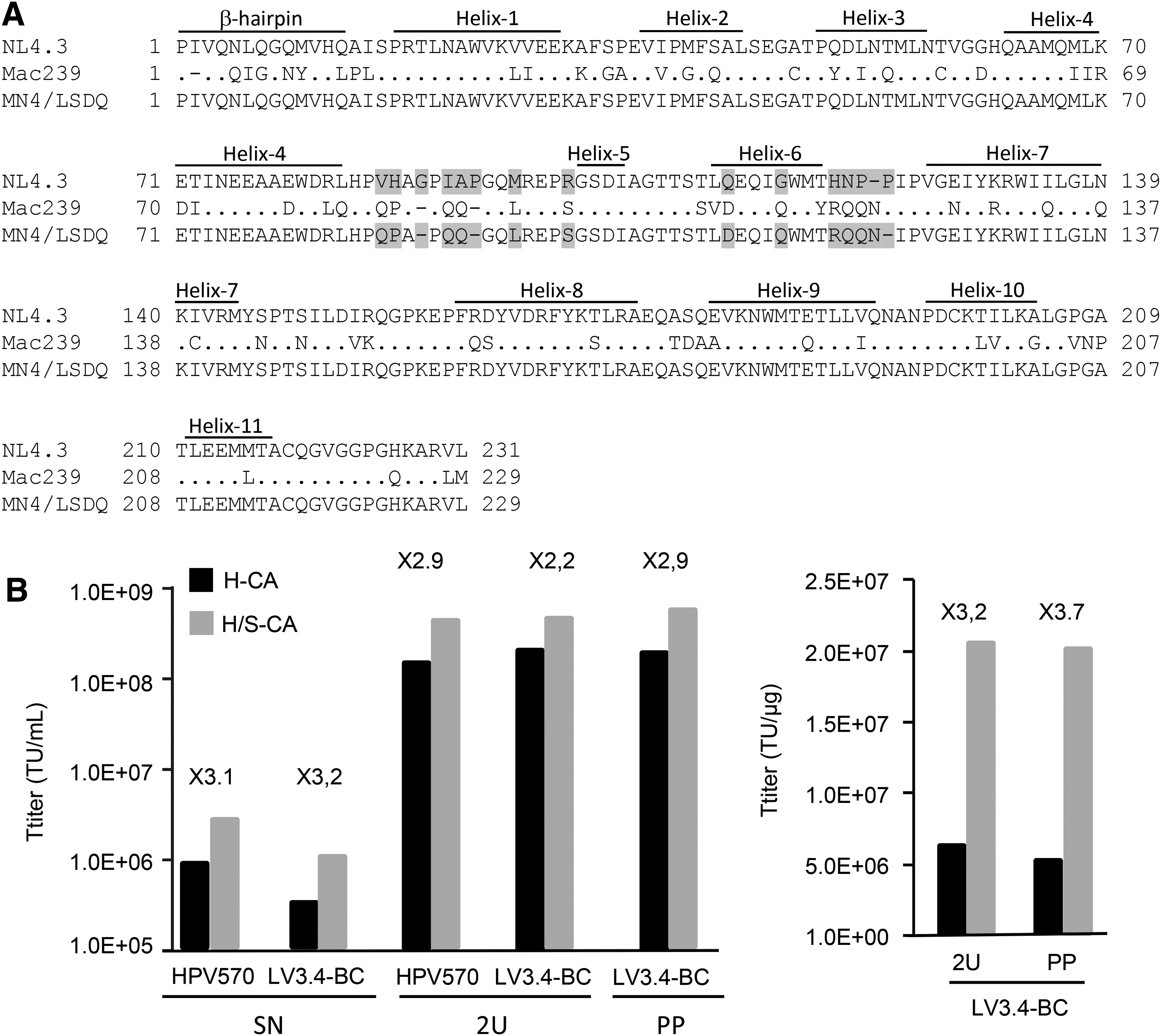
Capsid amino-acid substitution improves titers in NIH3T3 cells.
Crude LV extracts obtained with packaging plasmids encoding either the HIV-1 wild-type (H-LVs) or chimeric (H/S-LVs) capsid were produced in parallel. In one experiment, supernatants of HPV570H-CA and HPV570H/S-CA preparations were harvested and concentrated by two rounds of ultracentrifugation. In another experiment, LV3.4-BCH-CA and LV3.4-BCH/S-CA supernatants were split in half, and one half was purified and concentrated by precipitation, whereas the other half was concentrated by ultracentrifugation. Vectors were titered simultaneously in NIH3T3 cells. Functional titers of H/S-LVs were 2.2–3.2 times higher than those of H-LVs in NIH3T3 cells (Fig. 3B), whereas the CA (P24) concentrations measured by enzyme-linked immunosorbent assay were similar for H-LVs and H/S-LVs (not shown). Thus, when normalized relative to vector particle concentrations, the functional titer of concentrated H/S-LVs was also higher than that of H-LVs (Fig. 3B).
Chimeric capsid LVs outperform HIV-1-derived LVs in MCM CD34+ hematopoietic progenitor cells
Vector preparations concentrated by ultracentrifugation were used to transduce bone-marrow CD34+ cells at identical multiplicities of infection (MOIs). As NIH3T3 titers were higher for H/S-LV than for H-LV, vector volume for H/S-LV was one half to one third that for H-LV. Nevertheless, transduction rates for hematopoietic cells harvested from pooled myeloid and erythroid colonies were 1.7–9.5 times higher with H/S-CA than with the wild type (Fig. 4A). As the ultracentrifugation of LVs prepared from crude extracts increases the concentration of products decreasing cell transduction efficiency, 75 it was wondered whether the observed differences might be due to inhibitory compounds in the vector preparations. Thus, the vectors were purified before concentration and the comparisons were repeated. The results obtained were similar to those obtained with crude extracts (Fig. 4C). Transduction rates were also evaluated under the microscope for myeloid and erythroid progenitors from monkey Z. The transduction rates of erythroid and myeloid progenitors were different (Fig. 4B and D), but the effect of amino-acid substitution on transduction was similar to that observed in cells harvested and analyzed by fluorescence-activated cell sorting (FACS; Fig. 4A and C). The greater transduction efficiency in total cells and myeloid progenitors was confirmed with another vector backbone and cells from another animal (Supplementary Fig. S7A).

Capsid amino-acid substitution enhances hematopoietic stem/progenitor cell transduction. Simian CD34
For comparison of the level of transduction enhancement independently of basal transduction levels, the odds ratios of transduction efficiencies between the capsid groups were calculated, and the means were compared (Fig. 4E). Odds ratios differed between samples, but their means (∼5) were independent of MOI and preparation method (p = 0.78), indicating that the higher transduction rates observed were related to the chimeric capsid rather than other parameters. As expected, the mean odds ratios for transduction efficiencies between MOIs (Fig. 4F) were independent of vector capsid. However, they were higher with the purified vectors than with crude extracts (p = 0.04), indicating that lentiviral vector purification made it possible to transduce simian hematopoietic cells such that the transduction rate plateau was reached more slowly than with crude extract.
Unsurprisingly, higher percentages of transduced cells were obtained with a higher mean VCN (Fig. 5A and B). Expected transduction rates, calculated from mean VCN, assuming a Poisson distribution, were compared with the actual percentages of transduced cells (GFP+ cells). The slopes of the two lines obtained with H-CA and H/S-CA vectors were not significantly different, and the data for these vectors were therefore pooled (Fig. 5C). The slope value of 0.997 (±0.031) indicates that LVs transduce cells in a manner similar to that expected if the transduction distribution follows a Poisson distribution. Thus, the higher VCN obtained with H/S-CA does not result from higher rates of transduction in a cell subpopulation more susceptible to transduction but from the higher overall transduction efficiency of MCM CD34+ cells.
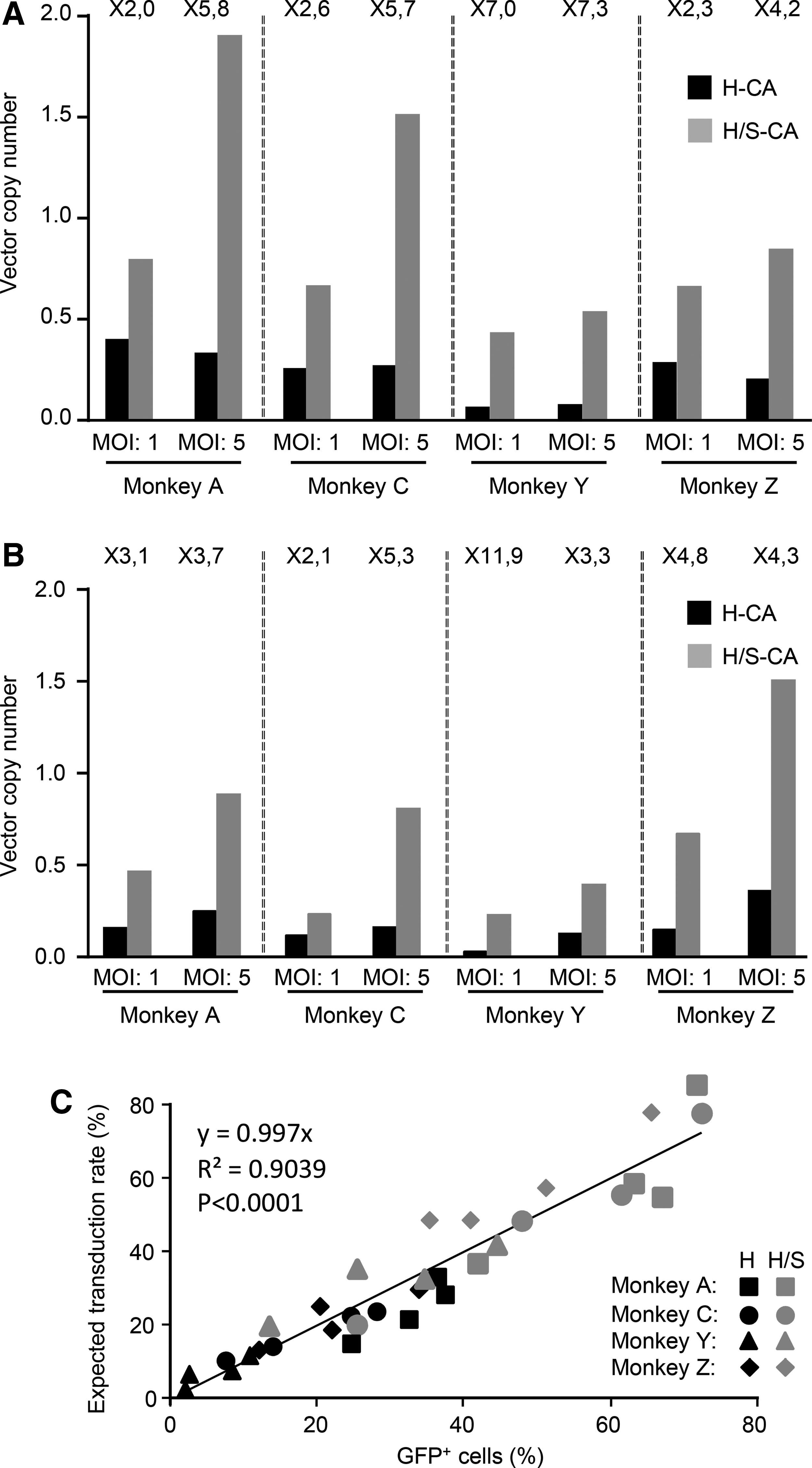
Capsid amino-acid substitution increases mean vector copy numbers (VCN) to the expected level. Transduction rates in cells harvested from methylcellulose (Fig. 4A and B) were evaluated by determining VCN.
Chimeric capsid LVs and HIV-1-derived LVs give similar transduction efficiencies in human cells
CA modification slightly decreased transduction rates in human cord blood cells (Supplementary Fig. S7A). However, an assessment of transduction efficiency on a per volume basis (vector preparation fractions of identical volume) rather than according to vector MOI showed transduction efficiencies to be similar in human cells (Supplementary Fig. S7B). In an independent experiment, two human cell lines (293T and HeLa cells), human CD34+ CB cells, two simian cell lines (cynomolgus Cynom-K1 and rhesus LLCMK2), and MCM CD34+ cells from animal A were transduced, and transduction efficiencies were determined. The absence of a difference in transduction rates in human cells was confirmed (Supplementary Fig. S8), despite the approximately six and ten times higher rates of transduction observed with H/S-CA in MCM primary cells and simian cell lines, respectively.
Chimeric LVs transduce short-term repopulating cells efficiently
A decision was made to evaluate transduction in less mature CD34+ cells from MCMs by analyzing cells after hematopoietic reconstitution in immunodeficient mice. Reconstitution with CM CD34+ cells is weak and transient in NSG mice. 76 Therefore, first, the most appropriate preparative conditioning and time window was determined. The highest proportion of monkey cells was obtained 6 weeks post infusion, following busulfan conditioning (Supplementary Fig. S9). Then, CD34+ cells were transduced with purified H-LV or H/S-LV, the transduced cells were infused into NSG mice, and the proportion of cells expressing GFP in vitro and after infusion was analyzed (Fig. 6). In this case, transduction levels were eight times higher with H/S-LV than with H-LV in vitro (Fig. 6A). GFP-expressing cells were scarce among the hematopoietic progeny of transplanted simian cells in the bone marrow and blood of animals into which H-LV-transduced CD34+ cells were injected (Fig. 6B and C). However, in animals receiving H/S-LV-transduced cells, the proportions of transduced simian CD45+ cells in mouse bone-marrow and blood samples (Fig. 6B) and of transduced simian CD34+ cells (Fig. 6C) were higher. It was not possible to calculate the fold increase, as most of the mice receiving H-LV-transduced cells had numbers of GFP cells below the limit of detection. As expected, simian cell engraftment levels were independent of the vector capsid (Supplementary Fig. S10).

Capsid amino-acid substitution enhances the transduction of short-term repopulating cells and activated T cells. The progeny of two million CD34+ cells transduced with purified LV3.4-BC1H-CA (H) or purified LV3.4-BC1H/S-CA (H/S) at a MOI of 5 was transplanted into 10 NSG mice treated with busulfan (5 mice per vector).
Enhancement of simian activated T-cell transduction
The ability of the chimeric LV to transduce T cells was evaluated. Peripheral blood cells from five MCMs (different from those described above) were isolated, activated, transduced, and analyzed by FACS (Fig. 6). Transduction efficiencies with H/S-LV were 6.9 and 8.4 times higher than those with H-LV in CD4 and CD8 T lymphocytes, respectively (Fig. 6D). The improvement in transduction rates was similar for activated (CD69+CD25+) and total CD4 or CD8 T cells (Supplementary Fig. S11A), and the proportion of lymphoid T cells changed little upon cell transduction (Supplementary Fig. S11B). For three of the five samples, FACS analyses were performed after an additional 3 days. The differences in transduction efficiency were even larger in CD8 T cells (Fig. 6E and F). Higher rates of transduction were also observed with H/S-LV for cells transduced before T-cell activation, although overall transduction rates were markedly lower (Supplementary Fig. S12) than those for activated T cells (Supplementary Fig. S11). The study also evaluated the performance of the chimeric H/S vector in rhesus primary T cells. As for MCM cells, transduction rate was higher with H/S-LV than with HIV-1-derived vector (Supplementary Fig. S13).
Chimeric capsid overcomes MCM TRIM5α restriction
This study showed that the enhanced rate of transduction in MCM cells resulted from the reversal of TRIM5α restriction by producing bicistronic LVs encoding either OFP (LV-Mock/I/O) or MCM TRIM5α and OFP (LV-TRIM5α/I/O) and using them to modify 293T cells. The resulting 293T-Mock/I/O cells, and 293T-TRIM5α/I/O cells, the OFP+ cells of which should express TRIM5α, were then transduced with identical volumes of LV3.4-BCH-CA and LV3.4-BCH/S-CA vector preparations. The OFP− and OFP+ cell fractions of 293T-Mock/I/O cells were transduced at similar rates—∼10%, ∼20%, and ∼35%—with the two vectors, as expected, and were therefore insensitive to the vector capsid (Supplementary Fig. S14A1, B1, C1, D1, and E1). As expected, total transduction levels were lower in 293T-TRIM5α/I/O cells. Surprisingly, however, they were also lower (∼1.6-fold) in the OFP− cell fraction of 293T-TRIM5α/I/O cells transduced with LV3.4-BCH-CA than in 293T-Mock/I/O cells, even though this fraction is thought not to express TRIM5α. The findings that (1) OFP fluorescence followed a continuous rather than discrete gradient in 293T-TRIM5α/I/O cells and (2) the mean orange fluorescence intensity in 293T-TRIM5α/I/O cells (Supplementary Fig. S14D2 and E2) was about half that in 293T-Mock/I/O cells (Supplementary Fig. S14D1 and E1) indicate that OFP is expressed at a lower level in 293T-TRIM5α/I/O cells than in 293T-Mock/I/O cells. As cap-dependent gene expression is significantly higher than IRES-dependent second gene expression in bicistronic constructs, 77 this observation suggests that some 293T-TRIM/I/O cells in which OFP is undetectable may actually express TRIM5α. The presence of such cells in the OFP− fraction of 293T-TRIM5α/I/O cells may explain the 1.6-fold decrease in transduction efficiency. Nevertheless, despite this anomaly, the ratio of transduction rates between vectors prepared with H/S-CA and H-CA in OFP+ cells (ratio = 3.3) was about double that in OFP− cells (ratio = 1.6), indicating that H/S-CA had the greatest potential for enhancing transduction in the cell fraction with the highest percentage of TRIM5α-expressing cells.
OFP− 293T-TRIM5α/I/O cells may not all be negative for TRIM5α expression, but OFP+ 293T-TRIM/I/O cells should all express TRIM5α. Thus, rather than comparing transduction rates between the OFP− and OFP+ fractions of 293T-TRIM5α/I/O cells, transduction rates were compared between the OFP+ fraction of 293T-Mock/I/O cells and the OFP+ fraction of 293T-TRIM5α/I/O cells (Fig. 7). With the wild-type HIV capsid, the transduction rate in TRIM5α-expressing cells was one tenth that in 293T-Mock/I/O cells. However, with the chimeric capsid, the transduction rate was reduced by only a factor of three. Thus, although the H/S capsid may not completely abolish MCM TRIM5α-mediated restriction, the observed transduction enhancement with the chimeric capsid in MCM cells is due to weaker TRIM5α-mediated restriction.
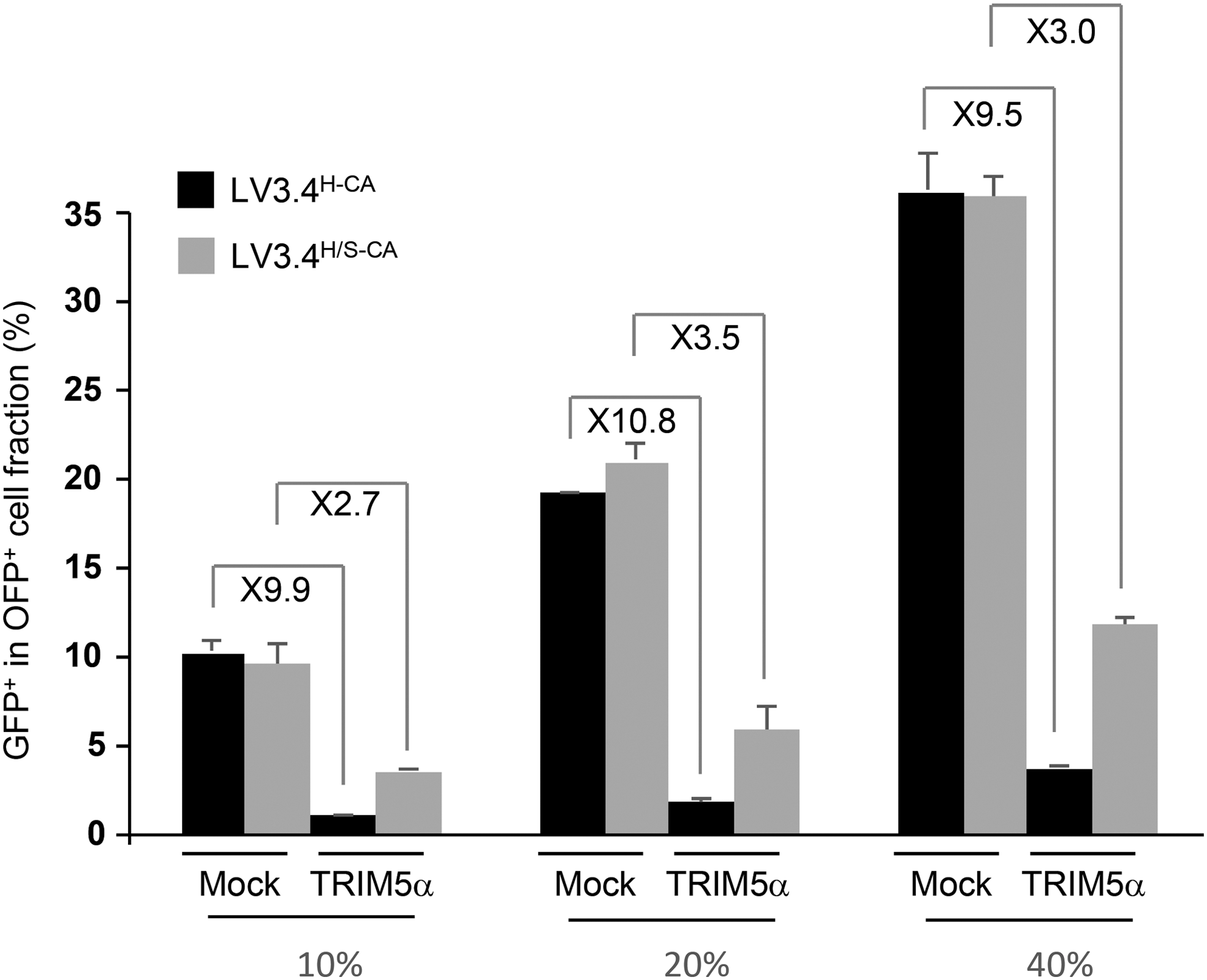
H/S capsid overcomes TRIM5α resistance. 293T-TRIM5α/I/O (TRIM5α) and 239T-Mock/I/O (Mock) cells were transduced with twofold dilution series of GFP-expressing vectors, LV3.4-BCH-CA and LV3.4-BCH/S-CA, such that transduction rates reached 10%, 20%, and 40% of 293T cells. The proportions of GFP+ cells were determined 4 days later in the orange fluorescent protein (OFP)+ cell fractions. The numbers indicated at the top are the ratios between transduction rates in 293T-Mock/I/O and 293T-TRIM5α/I/O cells transduced with LV3.4-BCH-CA and LV3.4-BCH/S-CA. Error bars are the standard deviations of cells transduced independently (n = 3).
Discussion
Preclinical animal models with a hematopoietic system similar to that in humans are required to reproduce clinical toxicities and for evaluations of the therapeutic potential of gene and cell therapy products during long-term follow-up. In this context, NHPs are the most relevant models of hematopoietic gene transfer for preclinical safety testing and long-term evaluations of gene and cell therapy products. Macaques are the most commonly used NHPs in biomedical research on infectious diseases 78,79 and in organ transplant research, 80 but their origin affects their susceptibility to immunodeficiency virus for several reasons, including the role of major histocompatibility complex (MHC) class I molecules in the immune control of virus replication 81 and host-encoded restriction factors. 82 Gene transfer to autologous HSCs and lymphoid T cells with HIV-1-derived LVs is an effective potential tool for the treatment of inherited hematopoietic diseases and for cancer immunotherapy. However, HIV-1 has a narrow host range. The key restriction factor, TRIM5α, can induce capsid disassembly, prevent the completion of reverse transcription, and alter HIV-1 replication in a species-specific manner. 25,83,84 Polymorphisms in the C-terminal (B30.2/SPRY) domain of TRIM5α orthologs act as major determinants of the spectrum of viruses recognized by a given ortholog, 85 –87 and affect immunodeficiency virus infections in individuals of the same species. 32,88,89 In some macaques, a CypA cDNA has retrotransposed into the TRIM5 locus, resulting in the replacement of the B30.2 domain with CypA. 38,41 Polymorphisms of the CypA domain of the fusion protein TRIMCyp affect its affinity for the HIV-1 core and its anti-lentiviral activity. 22,47
CMs were introduced into Mauritius about 400 years ago 90 from a very small founding population. This introduction event has resulted in lower levels of genetic variability than in other CMs 91 in the MHC class I region 92 –94 and lower TRIM5 allele diversity. 21,22 These macaques are thus a valuable model, relevant to humans, for therapeutic approaches, and are increasingly being used to test innovative therapeutic approaches, 95 –98 including gene transfer strategies. 99 –101 This study sequenced the TRIM5 exons of CMs imported from Mauritius, and it showed that they had no TRIM5Cyp allele, as previously reported for animals originating from this island. 21,22 The three missense mutations identified in MCM TRIM5α exon 8 to date 21,102 were detected at the expected frequency. The other MCM TRIM5α exons had not been sequenced before, but the polymorphisms observed in the N-terminal region (E4G), the coiled-coil domain (H178Y, K209E), and the second linker (E247D) have also been found in the TRIM5α 47 and TRIMCyp 22 sequences of CMs from other geographic locations. Their distribution in monkeys B, C, M, and Z indicate that the major Mafa4 haplotype, defined from sequences of the B30.2/SPRY domain of MCM, can be further divided into several subcategories. As exons were sequenced separately, it was not possible to reconstitute the amino-acid organization of each allele fully, but these results nevertheless highlight a previously unsuspected variability of TRIM5α in MCMs.
Whatever the relative level of permissiveness associated with intra-species polymorphisms, TRIM5α is an innate immune sensor for the retroviral capsid 103 that limits transduction with HIV-1 LVs in macaque hematopoietic cells. 28,53,61 Indeed, it was found that expression of the prototypical MCM TRIM5α in human 293T cells strongly affected the transduction of H-LVs. Therefore, the study investigated whether capsid modifications shown to relieve the TRIM5-mediated restriction of HIV-1 replication in CM and/or RM cells 55,57,58,63 also increased the efficiency of MCM hematopoietic cell transduction with HIV-1-derived LVs. Four series of missense mutations were incorporated into the capsid coding region of the HIV packaging construct to incorporate amino-acid substitutions shown to reduce early post-entry restriction in simian cells. CypA makes contact with residues from the loop between α-helixes 4 and 5 (L4/5) of HIV-1 CA and renders HIV-1 sensitive to restriction by TRIM5α. 104,105 Accordingly, conversion of the CypA binding loop-spanning residues 85–93 of HIV-1 CA (PVHAGPIAP) into the corresponding segment from SIV (PQPAPQQ) suppresses restriction in RM kidney cells. 55 In addition to L4/5 amino-acid substitutions, the replacement of amino acids located in a loop between α-helixes 6 ad 7 (L6/7) has been shown to enhance HIV-1 replication in a CM T-cell line, probably by abolishing interaction with TRIM5α. 63 The additional replacement of HIV-1 CA-Q112 with aspartic acid (D) further overcomes the replication block in macaque T cells but independently of TRIM5 protein restriction via an undefined mechanism. 58 Finally, the replacement of three additional amino acids involved in the interaction with TRIM5α by their SIVmac39 CA counterpart (M96, R100, and G116 from HIV-1 CA to L, S, and Q, respectively) is accompanied by an increase in resistance to macaque TRIM5 restriction in RM cells bearing homozygous restrictive TRIM5 alleles. 57 Therefore, L4/5, L6/7 and M96, R100, Q112, and G116 in HIV-1 CA were replaced with the corresponding amino acids from SIVmac239 in the packaging construct. The rate of CD34+ cell transduction was markedly improved by 2- to 15-fold for all samples. The observed increases were largest for samples and/or conditions for which transduction rates with the HIV capsid were lowest, suggesting that restriction levels may vary between samples and indicating that the impact of capsid modification was greatest when restriction was strongest. Maximal transduction efficiencies differed between samples, suggesting that the CA modifications introduced here did not result in the complete evasion of TRIM5α restriction, consistent with the observations for 293T cells expressing MCM TRIM5α and with previous observations reported by Nomaguchi et al. 57 who were unable to abolish the TRIM5-mediated restriction of HIV infection completely in human cells expressing macaque TRIM5α. Alternatively, this finding may highlight the differential expression of other restriction factors 106 and a need for complementary strategies to optimize the transduction of simian MCM cells further.
In preclinical studies for gene therapy, hematopoietic CD34+ cells from macaques have been cultured in several serum-containing and serum-free media before ex vivo transduction. 28,107 –109 This study showed that culture conditions known to sustain human stem-cell survival 71 promoted the expansion of MCM CD34 progenitor cell populations, although the optimal culture conditions have yet to be determined. The study investigated the effects of cell concentration on cell expansion and transduction, and found that both cell proliferation rates and transduction efficiencies were higher when CD34+ cells were grown at a density of <0.5 million cells/mL.
Other studies have investigated ways of bypassing restriction factors and improving the HIV-1-based LV transduction of simian hematopoietic cells. A chimeric LV (χHIV) in which the vector genome is packaged within the entire SIV capsid, coupled to a high MOI, has been shown to result in RM CD34+ cell transduction rates similar to those obtained with SIV-derived vectors. 51 Modifications of small regions have made it possible to identify key groups of amino acids, the substitution of which improves the HIV-1-based LV transduction of monkey cell lines, 110,111 baboon CD34+ cells, 111 RM peripheral blood cells, 62 and RM primary T cells. 61 The LNEIE HIV-1 quintuple mutant is resistant to the disruptive effect of RM TRIM5α and replicates in rhesus primary lymphocytes. 60 In the context of LVs, the LNEIE-packaged HIV-1 vector had a titer half that of the wild type in human cells, but yielded four times higher rates of rhesus primary T-cell transduction. 61 The MN4/LSDQ HIV-1 derivative is also resistant to RM TRIM5α, 57 and a side-by-side comparison with the LNEIE HIV-1 clone showed it to be less susceptible to various macaque TRIM5 proteins. 56 Improvements in T-cell transduction efficiency are vital because studies in NHP models are required to investigate the mechanisms driving chimeric antigen receptor T-cell toxicities and to evaluate treatments for preventing or abolishing them. 112 CD34+ hematopoietic cells are another important target for gene therapy studies in NHPs. This study compared NL4.3- and MN4/LSDQ-derived capsids in LVs, and assessed transduction efficiencies for both T cells and CD34+ cells in a model frequently used for biomedical research. It showed that the chimeric MN4/LSDQ-derived capsid, embedded in an HIV-1-derived packaging system, gave higher transduction rates with both MCM hematopoietic CD34+ cells (in vitro and in xenografted NSG mice) and MCM lymphoid (CD4+ and CD8+) T cells, and with MCM TRIM5α-expressing 293T cells, than a vector prepared with an HIV-1-derived capsid. It also transduced the rhesus TRIM5α sensitive cell line LLCMK263 10 times more efficiently, yielding results similar to those obtained with SIV-derived vectors. 113,114 Transduction enhancement in simian cell lines and primary MCM CD34+ cells yielded results similar to those obtained with the chimeric χHIV vector 53 . Therefore, the modification of key amino acids, such as those introduced into the HIV-1 clone MN4/LSDQ, circumvents MCM TRIM5α-mediated restriction, and resistance seems to be as efficient as with SIV-based LVs and chimeric HIV-1 vectors, including the complete simian capsid.
By contrast to the sustained high-level hematopoietic reconstitution achieved by injecting human HSPCs into immunodeficient mice preconditioned by total body irradiation or with busulfan, 115 the reconstitution of hematopoiesis with simian cells is inefficient and only transient. 76 Thus, even though capsid modification increased transduction rates in short-term repopulating cells, it was not possible to demonstrate an increase in transduction rates in long-term stem cells. This aspect requires further study in transplantation assays in monkeys.
The phenomenon underlying the higher rates of transduction in mouse NIH3T3 cells is currently unknown. The mouse Fv1 capsid binding factor has no restriction activity against HIV-1 and SIVmac. 116 However, HIV-1-derived vectors harboring single amino-acid substitutions in the CypA binding loop have been shown to yield higher transduction rates in mouse induced pluripotent stem cells. 117 A similar restrictive phenotype in NIH3T3 cells may thus be relieved by the use of the MN4/LSQD capsid.
In conclusion, transduction was enhanced by a factor of 2–10 in simian HSPCs and by a factor of 6–15 in T cells. As with the replacement of the entire HIV capsid with the SIV capsid, 53 the modifications made did not affect transduction efficiency in human hematopoietic CD34+ cells. CYPA-binding mutants are known to use nuclear entry routes different from those used by viruses with wild-type capsids and to integrate preferentially into genomic regions with a higher density of transcription units than wild-type viruses in human cells. 118 It will therefore be important in the context of a gene transfer model for long-term toxicity studies to analyze integration site preferences in human and simian cells transduced with the chimeric capsid described here and to compare integration profiles with those of other chimeric vectors and with those observed in human cells transduced with H-LV-derived vectors.
Footnotes
Acknowledgments
This work was supported by an Industrial Chair (HemGenTher) from France's “Agence Nationale pour la Recherche” (ANR) awarded to P.L. and by the “Programme Investissements d'Avenir” (PIA) managed by the ANR under reference ANR-11-INBS-0008, funding the Infectious Disease Models and Innovative Therapies (IDMIT) infrastructure (Fontenay-aux-Roses, France). We thank IDMIT staff for providing us with access to the animal science core facility and the veterinary surgeon Dr. Christophe Joubert (iDMIT) for his expertise with NHPs, as well as Simon Rascalou (bluebird bio France, Fontenay aux Roses, France) for technical assistance.
Author Disclosure
The authors have no competing financial interests to declare.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Figure S11
Supplementary Figure S12
Supplementary Figure S13
Supplementary Figure S14
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.