
Abstract
The failure to maintain the viability of ischemic myocardium is one of the mechanisms that causes ischemic heart dysfunction after revascularization. Hibernating myocardium is considered to be able to maintain long-term viability during chronic hypoperfusion. Pigment epithelium-derived factor (PEDF) decreases the contractility of hypoxic cardiomyocytes and protects cardiomyocytes against ischemic injury, which is strikingly similar to the pathophysiologic characteristics of hibernating myocardium. It was therefore postulated that PEDF may induce acute ischemic myocardium into a “hibernating-like” state to maintain its viability. Adult Sprague–Dawley rat models of acute myocardial infarction were surgically established. Lentiviral vectors carrying the PEDF gene (PEDF-LVs) were delivered into myocardium with infarction to overexpress PEDF locally. It was found that PEDF local overexpression significantly reduced myocardial infarct size and cardiomyocytes necrosis but did not improve cardiac function at rest. The contractile reserve assessed by low-dose dobutamine stress echocardiography and “perfusion-metabolism mismatch” assessed by positron emission tomography, which are the characteristics of viable myocardium in hibernation, were observed in the PEDF overexpressed ischemic heart. Ultrastructural changes observed by electron microscopy and glycogen deposition explored by Periodic acid–Schiff staining were similar to the histological characteristics of hibernating myocardium. Moreover, PEDF overexpression protected the cardiomyocytes against anoxic injury and retained their functional recovery potential after reoxygenation in vitro. PEDF local overexpression may induce acute ischemic myocardium into a “hibernating-like” state and maintain its viability. This novel effect of PEDF presents an important clinical approach to enhance functional recovery after revascularization therapy in acute myocardial infarction.
Introduction
Timely and sustained reperfusion is essential for myocardial salvage after acute myocardial infarction (AMI), 1 thus preserving left ventricular function and providing favorable long-term prognostic outcomes. 2 Nonetheless, there seems to be a limited time window in which myocardium can be salvaged. 3 Due to severe ischemia and anoxia, cardiomyocytes tend to lose their viability in a short period of time followed by scar formation. 4 Reperfusion therapy is not effective in the presence of necrotic or scarred tissue. 5 As a result, a feasible strategy for maintaining the viability of acute ischemic myocardium is highly needed to improve therapeutic effect of reperfusion and to achieve maximum left ventricular function recovery. 6
Hibernating myocardium, as originally described by Rahimtoola in 1984, 7 is a condition of myocardium that is thought to maintain long-term viability after chronic hypoperfusion. 8 During hibernation, the contractility and metabolism of cardiomyocytes decrease in order to adapt to the reduction in blood supply, thus preserving their viability and preserving their functional recovery potential. 9 Therefore, hibernating myocardium has been well described as one of the most important determinants of ischemic left ventricular functional recovery after revascularization. 10 However, studies have demonstrated that there are only a small number of viable cardiomyocytes at the edge of the infarction area after AMI. 11 Transforming more acute ischemic myocardium into a hibernating state after AMI may be an effective strategy to maintain its viability and enhance its functional recovery.
Pigment epithelium-derived factor (PEDF) is a 50 kDa secreted glycoprotein belonging to the SERPIN superfamily. It is commonly expressed in multiple tissues 12 and has been implicated in many cardiovascular diseases. 13 It has previously been suggested that PEDF treatment can protect cardiomyocytes against ischemia and hypoxia-induced cell apoptosis. 14 In addition, gene therapy has been reported as an attractive strategy for increasing PEDF local expression in myocardium with infarction, thereby protecting ischemia myocardium and reducing left ventricular infarct size. 15,16 However, the direct effects of PEDF overexpression on myocardial viability have not been evaluated. Recently, it was found that PEDF decreased the contractility of hypoxic cardiomyocytes and maintained the functional reserve of the ischemic heart. 17 The decreased contractility of cardiomyocytes observed in the study is not regarded as the consequence of a sustained energy deficit but instead as a regulatory process that serves to avoid an energy deficit. It is likely that PEDF reduces the contractility and energy requirements of ischemic cardiomyocytes so as to adapt to the anoxic environment. This acute adaptive mechanism induced by PEDF in AMI is strikingly similar to the pathophysiologic characteristics of chronic hibernating myocardium. 9 It was therefore hypothesized that PEDF may transform acute ischemic myocardium into a “hibernating-like” state and maintain its viability after AMI.
During ischemia, viable myocardium in the hibernating state reveals some typical attributes that are used for its identification. 18,19 Functionally, it is characterized as being dysfunctional while retaining a contractile reserve, which can be demonstrated by low-dose dobutamine stress echocardiography (LDDE). Metabolically, it is characterized by decreased blood flow accompanied with sustained glucose uptake, which is usually assessed by positron emission tomography (PET). Morphologically, hibernating myocardium displays some typical structural changes, such as little evidence of ischemic damage, glycogen accumulation, and a mass of small, dense mitochondri. 20 The present study developed a rat model of AMI and delivered lentiviral vectors (LV) carrying the PEDF gene (PEDF-LVs) to overexpress PEDF locally in myocardium after infarction. The aims were to describe the special viable state induced by PEDF overexpression in acute ischemic myocardium and to investigate whether this viable state is consistent with the typical characteristics of hibernating myocardium. In addition, the study verified whether the contractile dysfunction of hypoxic cardiomyocytes after PEDF overexpression was recoverable using in vitro experiments.
Methods
Recombinant LV construction and viral production
Recombinant LVs carrying the PEDF gene (PEDF-LVs; Shanghai GeneChem Co. Ltd., Shanghai, P.R. China) were prepared, as described previously. 16 The PEDF gene was cloned into the lentiviral expression plasmid pGC287 using AgeI and BamHI restriction sites. The sequences of the PEDF primers used were as follows: forward 5'-CGACCGGTGCCACCATGCAGACCCTGG-3′, and reverse 5′-GGAATTCGGATCCTTAAGTGCTGCTGG-3′. LVs were produced by triple transient transduction. The expression plasmid, packaging plasmid, and envelope plasmid were mixed and were then packaged by 293T cells (American Type Culture Collection, Manassas, VA). LVs without PEDF gene were constructed as blank vector controls. All the LVs were constructed and labeled with green fluorescent protein (GFP). The concentrated titer of the virus suspension was 2 × 1012 TU/L.
Animal model and intra-myocardial gene delivery
Sprague–Dawley male rats (8–10 weeks old, weighing 210–250 g, 230 ± 20 g) were obtained from the Experimental Animal Center of Xuzhou Medical University and cared for in accordance with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health. Animal procedures were approved by the Xuzhou Medical University Committee on Animal Care. All experiments conformed to the international guidelines for the ethical use of animals. The AMI model was induced by ligation of the left-anterior descending coronary artery (LAD) in anesthetized rats, as described previously. 21 The animals were randomly divided into four groups: (1) sham rats that underwent the surgical procedure without LAD ligation; (2) control rats that were injected with 20 μL of enhanced infection solution (ENIS) as a solvent control; (3) vector the rats that injected with blank LVs that did not carry the PEDF gene as a blank vector control; and (4) PEDF rats that were injected with PEDF-LVs.
The rats were anesthetized with sodium pentobarbital (60 mg/kg) intraperitoneally (i.p.) and maintained under anesthesia using isoflurane (1.5–2.0%) mixed with air. The absence of the pedal reflex was considered as an indication that a surgical plane of anesthesia has been maintained. The rats were placed in a supine position, and a left thoracotomy was performed through the fourth intercostal space under sterile conditions. Then, PEDF-LVs (2 × 107 TU) in 20 μL ENIS (GeneChem, Shanghai, P.R. China) was delivered with a 20 μL syringe and 25-gauge needle into the myocardium along the LAD, and the chest cavity was then closed. Equivalent volumes of ENIS and blank LVs were delivered as solvent control and vector control, respectively. After reinstallation of spontaneous respiration, animals were extubated and allowed to recover from anesthesia, and postoperative analgesia was performed with buprenorphine administration at 0.5 mg/kg. A standard diet was maintained after surgery 5 days later. The LAD was ligated with 6-0 silk suture (Ethicon; Johnson & Johnson, New Brunswick, NJ) with the same surgery. Then, animals accepted the corresponding test after induction of AMI or were sacrificed with an overdose of sodium pentobarbital (100 mg/kg intravenously [i.v.]), and hearts were harvested for further analysis. Sham-operated rats underwent the same procedure, excluding LAD ligation.
GFP expression investigation
GFP expression was used to evaluate the transduction efficiency of LVs. After 5 days of LV transduction, the LAD was ligated to establish an AMI modal. The hearts were harvested before and 3 days after LAD ligation. All biopsy tissues were taken from the LAD distribution regions in the rat hearts. Frozen myocardial tissue was horizontally sliced into 8 μm sections and mounted on glass slides. Sections were fixed in 4% paraformaldehyde for 15 min. The sections were observed by a fluorescence microscope (Olympus IX73; Olympus Corp., Tokyo, Japan). GFP fluorescence was calculated by viewing four randomly selected fields for each section. Digitized images were analyzed using Image-Pro Plus v6.0 (Media Cybernetics, Inc., Bethesda, MD). The GFP intensity was expressed as the percent of transduced tissue area by GFP fluorescence. All investigators were blinded to the myocardial source in all analyses.
Measurement of plasma creatine kinase isoenzyme MB and lactate dehydrogenase
Myocardial cellular damage and necrosis were evaluated by measuring plasma levels of creatine kinase isoenzyme MB (CK-MB) and lactate dehydrogenase (LDH). Blood samples (1 mL) were drawn before the hearts were harvested and centrifuged to obtain plasma (14,000 g for 10 min at 4°C). LDH and CK-MB levels were measured using a MindrayBS-120 Chemistry Analyzer (Mindray Medical Corp., Shenzhen, P.R. China). All samples were measured in duplicate.
Determination of myocardial infarct size
One, three, and seven days after LAD ligation, 2% Evans Blue dye (30 mg/kg; Sigma–Aldrich, St. Louis, MO) was injected i.v. for 10 min, and selected rats were euthanized. Then, the hearts were removed for myocardial infarct size analyses using 2,3,5-triphenyltetrazolium (TTC) staining. Hearts were frozen at −20°C and cut into 4 mm slices perpendicular to the axis of the LAD. Slices were immediately immersed in 1% TTC (Sigma–Aldrich) in phosphate buffer (pH 7.4) at 37°C for 10 min to discriminate infarcted tissue from viable myocardium. All slices were scanned from both sides by a color CCD camera (FV-10; Fujifilm, Tokyo, Japan). Morphometric measurements of infarct area (INF) were performed by digital planimetry software (Image-Pro Plus 6.0; Media Cybernetics, Inc.) Myocardial infarct size was expressed as percentage of the infarct area (INF) over total left ventricular (INF/left ventricular × 100%).
Animal echocardiography test at rest and low-dose dobutamine stress
Echocardiography was conducted under sedation by sodium pentobarbital (30 mg/kg i.p.), as described previously. 17 It was performed at rest and during low-dose dobutamine stress at the end of 1, 3, and 7 days after induction of AMI. Two dimensional–guided M-mode echocardiography was used to determine left ventricular chamber volume at systole and diastole and contractile parameters, such as left ventricular end-diastolic dimension (LVEDD), left ventricular end-systolic dimension (LVESD), left ventricular end-diastolic volume (LVEDV), left ventricular end-systolic volume (LVESV), and left ventricular anterior wall thickness. Left ventricular fractional shortening (LVFS) was calculated as follows: FS (%) = (LVEDDLVESD)/LVEDD × 100%. The ejection fraction (EF) was then derived as EF (%) = (EDVESV)/EDV × 100%. All measurements were based on the average of at least three cardiac cycles. Dobutamine (1 μg/g body weight; Sigma–Aldrich) was given i.p. Cardiac reserve was investigated 10 min after dobutamine injection.
PET
PET was performed by MITRO Biotech Co. Ltd. (NanJing, P.R. China). In brief, randomly selected animals were persistently anesthetized with a rodent anesthesia machine using isoflurane (1.5–2.0%) mixed with air 3 days after AMI. The tail vein was cannulated for injection of 13 N-NH3 (300 ± 150 μCi). A micro PET (Siemens, Munich, Germany) dynamic scan was performed immediately after 13 N-NH3 injection for rest myocardial perfusion imaging, and the scanning time was set to 10 min. Then, after at least 40 min of 13 N-NH3 washout period, 18 F-FDG (300 ± 150 μCi) was injected in the tail vein of the same rat. A micro PET static scan was performed again 1 h after 18 F-FDG injection for myocardial metabolic imaging. Then, images were reconstructed and polar maps were generated using FlowQuant software. The reconstruction algorithm was OSEM 3D, and the number of iterations was 2. The image and data were processed after reconstruction. The normal myocardium and ischemia area were delineated as areas of interest. Then, the standard uptake value (SUV) of the area of interest and the volume of the ischemic area and the entire heart were calculated. All the PET data were interpreted in a blinded manner both quantitatively and qualitatively by a physician (R.E.C.) experienced in reading cardiac PET scans.
Plasma glucose and insulin levels testing
Plasma glucose and insulin levels were tested before PET. Blood (100 μL) was drawn from an orbital vein of the rat using a capillary micropipette (Ringcaps; Hirschmann Laborgerate, Eberstadt, Germany) after anesthesia. Plasma glucose levels were determined by placing the whole blood (1 μL) on a glucose test strip, which was immediately analyzed using a blood glucose analyzer (Ascensia Breeze; Bayer, Mishawaka, IN). Plasma insulin levels were measured using an Ultra High Sensitive Rat Insulin Measurement Kit (Morinaga Institute of Biological Science, Tokyo, Japan) according to the manufacturer's protocol.
Electron microscopy imaging
For electron microscopy observation, small samples of heart tissue were fixed in 2.5% glutaraldehyde overnight and then incubated in 1% osmium tetroxide for 2 h with light-proofing. After washing in distilled water, specimens were incubated in 2% uranyl acetate for 2 h at room temperature and then dehydrated in graded ethanol concentrations. Finally, samples were embedded in molds with fresh resin. Ultrathin sections were cut with an EM UC7 (Leica Microsystems GmbH, Wetzlar, Germany), stained with lead citrate, and examined with a Tecnai G2 T12 (FEI, Hillsboro, OR).
Rat neonatal ventricular cardiomyocytes isolation, culture, and transduction
Neonatal cardiomyocytes were isolated from 1-day-old newborn Sprague–Dawley rats. 22 Briefly, neonatal rats were anesthetized with sodium pentobarbital and decapitated. Hearts were rapidly removed into dishes on ice, and the vessels and atria were discarded. The ventricles were then dissected and minced into 1 mm 3 pieces and transferred to a sterile tube, washed in cold phosphate-buffered saline (PBS) solution (136.9 mmol/L NaCl, 2.7 mmol/L KCl, 8.1 mmol/L Na2HPO4, 1.5 mmol/L KH2PO4, pH 7.3) to remove blood clots. Minced tissue was digested in a PBS solution supplemented with 1 mg/mL trypsin, 1 mg/mL collagenase type II, and 0.2 mg/mL glucose for 5 min at 37°C and incubated with 0.1 mmol/L BrdU to enrich for cardiomyocytes selectively by inhibiting the growth of cardiac fibroblasts. Cardiomyocytes were then purified using differential adhesion method. Hypoxia was achieved by culturing cells in D-Hank's liquid with glucose deprivation in a tri-gas incubator (Heal Force, Shanghai, P.R. China) saturated with 5% CO2/1% O2 at 37°C for the indicated time periods. To regulate the PEDF expression, PEDF-LVs were transfected following the manufacturer's protocol at the desired multiplicity of infection (MOI = 20). PEDF-LVs were diluted to 2 × 109 TU/L in growth medium and added to cardiomyocytes. After a transduction period of 8 h, the medium was removed, and fresh medium was added. After an additional 24 h, GFP co-expression on the construct was used to determine the efficiency of viral transduction, and the expression of PEDF in neonatal cardiomyocytes was confirmed by Western blot analysis.
Periodic acid–Schiff glycogen staining and glycogen content assay
Glycogen staining was performed using a Periodic acid–Schiff (PAS) kit (395B; Sigma–Aldrich). Semithin sections or cardiomyocytes that were fixed with 4% paraformaldehyde were oxidized with periodic acid for 5 min at room temperature, washed, and incubated in Schiff's reagent for 15 min at room temperature. Next, they were washed and counterstained with hematoxylin solution for 90 s, washed, and dehydrated, and the cover slips were mounted on slides using 50% glycerin. Images were acquired using a color CCD camera (FV-10; Fujifilm). Glycogen contents were measured using two enzymes, colorimetric glucose assay by Glycogen Assay Kit (MAK016; Sigma–Aldrich) following the manufacturer's instructions. Relative quantitative analysis of the change in glycogen levels was normalized to normal groups.
Protein extraction
For protein extraction, rat heart tissue from the left ventricular myocardium or rat neonatal ventricular cardiomyocytes were solubilized using a lysis buffer (100 mmol/L Tris-HCl, 4% sodium dodecyl sulfate, 20% glycerine, 200 mmol/L DTT, phosphatase and protease inhibitors, pH 6.8). Protein concentrations were measured using a bicinchoninic acid (BCA) assay.
Western blot analysis
An equivalent amount of protein was prepared and separated by 8–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and electro-transferred to nitrocellulose membranes (Millipore, Darmstadt, Germany). This was then probed with PEDF antibodies (1:1,000, DF6547; Affinity Biosciences, Cincinnati, OH), RIP3 (1:1,000, 17563-1-AP; Proteintech, Rosemont, MN), or Tubulin (1:1,000, 2146; Cell Signaling Technology, Beverly, CA) at 4°C overnight, and incubated with secondary antibody for 2 h at room temperature. Finally, signals were detected by Odyssey Infrared Imaging System (Li-Cor Biosciences, Lincoln, NE). Digitized images were analyzed using ImageJ (NIH, Bethesda, MD). For all Western blot analyses, other than when specifically noted, protein levels were calculated from the ratio of corresponding protein/Tubulin.
Cell viability assay
Cardiomyocytes were seeded onto 96-well plates (Corning, New York, NY) at a concentration of 1 × 104 cells/mL. Cell viability was detected using a cell counting kit-8 (CCK-8; DOJINDO, Tokyo, Japan). Absorbance at 450 nm was measured with a microplate reader (BioTek Synergy 2, Winooski, VT). The means of the optical density (OD) measurements from six wells of the indicated groups were used to calculate the percentage of cell viability.
Measurement of single-cell contractility
Cardiomyocytes were cultured for 24 h in standard growth medium in Plexiglas superfusion chambers, in which the bottom was formed by a collagen-coated glass coverslip. The chamber was then placed on the stage of an inverted microscope (Nikon Diaphot; Nikon Corp., Tokyo, Japan), and cells were cultured in D-Hank's liquid with glucose deprivation in a tri-gas incubator saturated with 5% CO2/1% O2 at 37°C for 24 h. Cell contraction was achieved at a voltage of 30 V and frequency of l Hz of electrical stimulation. Cell shortening was measured using a video edge detection system (Crescent Electronics, Sandy, UT). The signal was acquired using a Data Q DI-200 board interfaced to a personal computer and stored using WinDaq Software (DATAQ Instruments, Inc., Akron, OH). The cell image was also recorded on videotape for additional off-line analysis. Amplitude of contraction under hypoxia condition was expressed relative to control values. Contractile amplitude under normoxic condition was set to 100%. Then, glucose and serum concentrations were restored by supplementing the culture medium with glucose and fetal bovine serum and incubated under normoxic conditions at 37°C for 72 h (recovery) after 16 h of hypoxia. Cell shortening was measured using a video edge detection system (Crescent Electronics) again.
Statistical analysis
Data are expressed as the mean ± standard deviation. Data between two groups were compared using two-tailed Student's t test, and multiple comparisons utilized one-way analysis of variance followed by a Student–Newman–Keuls test. p-Values of <0.05 were considered significant.
Results
Intra-myocardial injection of PEDF-LVs induces PEDF local overexpression
First, the PEDF expression level was measured in the rat hearts by Western blot. Results showed that the basal protein level of PEDF was high in normal myocardium, which then gradually decreased after AMI in the control group (Fig. 1A and D). The PEDF expression trends in the vector group were similar to the control group (Fig. 1B and D). However, Western blot in the PEDF group showed that PEDF-LV transduction maintained a continuous high level of PEDF expression after AMI for 7 days (Fig. 1C and D). These findings demonstrate that PEDF-LV transfection successfully produces stable local PEDF overexpression.

Validity of PEDF-LV transduction.
Both the blank LVs and PEDF-LVs used in this study were constructed and labeled with GFP. To evaluate the transduction efficiency of lentivirus 5 days after injection, GFP expression in myocardium was investigated before or 3 days after AMI (Fig. 1E). Results showed that there was no significant GFP expression in the sham and control groups either before or 3 days after AMI. However, a great deal of GFP expression was observed in the vector and PEDF groups before AMI. GFP expression in the vector group was significantly decreased 3 days after AMI, which may be attributed to a large number of cardiomyocytes dying after LAD ligation. There were still large quantities of GFP expression in the PEDF group 3 days after AMI, but the intensity was lower than transduced normal myocardium (Fig. 1F). These results suggest that lentivirus transduction using intra-myocardial injections can achieve high transduction efficiency.
PEDF overexpression reduces myocardial infarct size and protects myocardium against ischemia-induced necrosis after AMI
To clarify the cardioprotective effects of PEDF, Evans blue/TTC double staining was performed to measure left ventricular infarct size 1, 3, and 7 days after AMI. The representative images are shown in Fig. 2A. The results indicated that LAD ligation resulted in a significant ischemic region. However, PEDF overexpression significantly reduced the myocardial infarct size, and the gap was largest 3 days after AMI (Fig. 2B). In addition, the serological levels of CK-MB and LDH (markers of myocardial injury) were significantly decreased in the PEDF group compared to the control group or vehicle group (Fig. 2C and D). Necrosis is one of the main causes of cardiomyocyte death and heart failure after AMI. 23 To determine whether PEDF overexpression could attenuate ischemia-induced myocardial necrosis, protein expression of RIP3 (a necrosis-associated protein) was tested in ischemic myocardium. 24 Results showed that the RIP3 protein level increased 3 days after AMI, but PEDF local overexpression significantly diminished the increase in RIP3 (Fig. 2E and F). Taken together, these results suggest that PEDF overexpression confers cardio-protection against ischemic injury after AMI in a rat model.
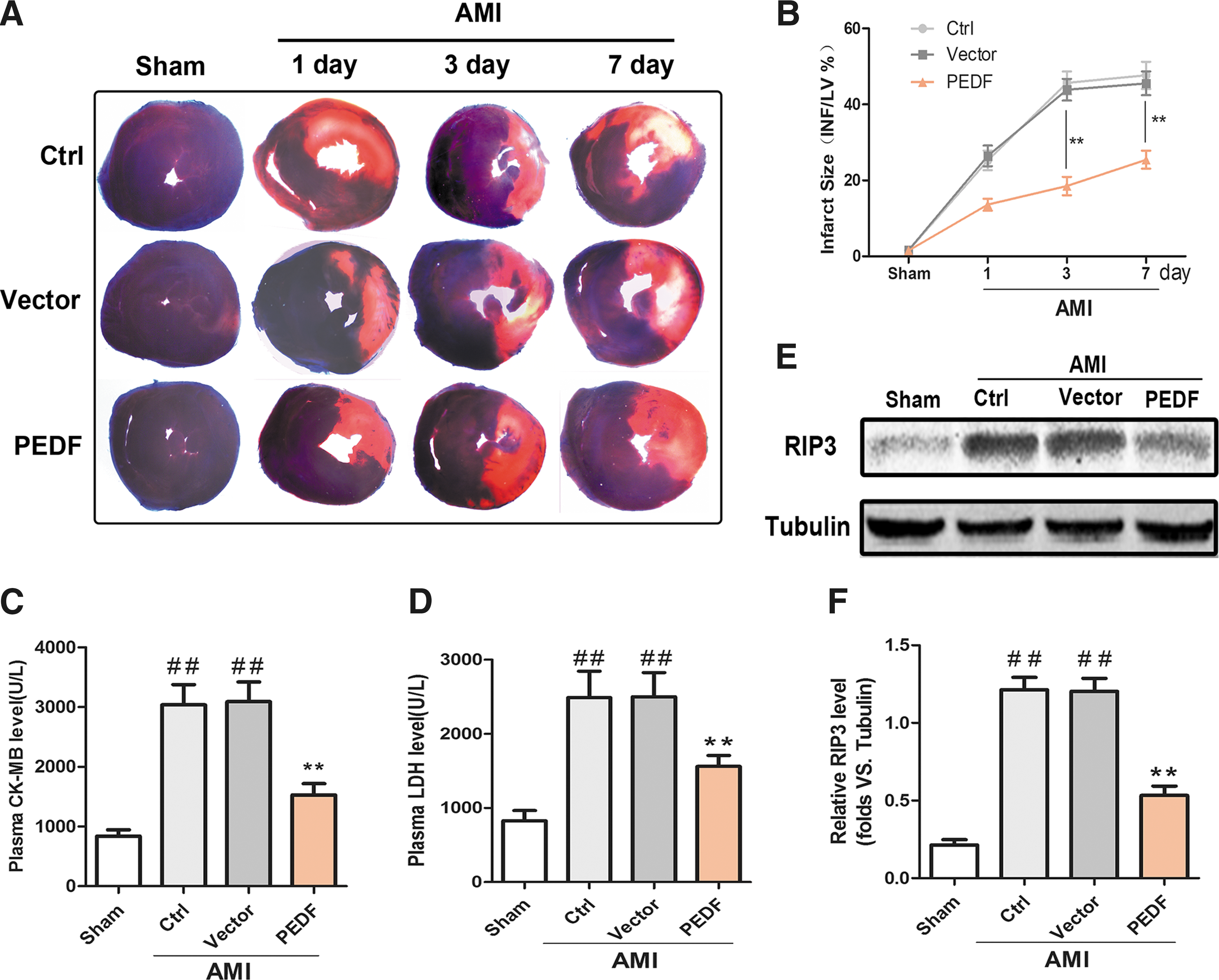
Myocardial infarct size and necrosis evaluation after AMI.
PEDF overexpression does not increase cardiac systolic function at rest but increases cardiac contractile reserve after AMI
Next, the effect of PEDF overexpression on cardiac function after AMI was investigated. Echocardiographic examination was performed in the rat models 1, 3, and 7 days after AMI (Fig. 3A). First, the cardiac systolic function at rest was examined. As shown in Fig. 3B and C, the cardiac systolic function was significantly decreased after myocardial infarction. Although the values of left ventricular EF % and FS % in the PEDF group were slightly lower 1 day and slightly higher 7 days after AMI compared to the control group or vector group, no significant difference was found between groups. Even so, the left ventricular ESV was significantly lower and the left ventricular anterior wall thickness was significantly higher in the PEDF group 7 days after AMI compared to the control group or vector group (Fig 3D and E). It is noteworthy that the lower ESV and thicker left ventricular wall assessed by echocardiography at rest are considered as indicators of myocardial viability. 25 When low-dose dobutamine was injected, there was a significant improvement in cardiac contractile function in the PEDF-LV transduced hearts. The increased value of EF % and FS % (ΔEF% and ΔFS%) after dobutamine injection were both higher in the PEDF group than in the control group or vector group, and the gaps were largest 3 days after AMI (Fig. 3F and G). These results indicate that PEDF overexpression in ischemic myocardium does not increase cardiac function at rest but maintains the responsiveness of ischemic myocardium to low-dose dobutamine, which is an indicator of functional reserve. Notably, these functional features of acute ischemic myocardium induced by PEDF overexpression are similar to the functional characteristics of hibernating myocardium. 26
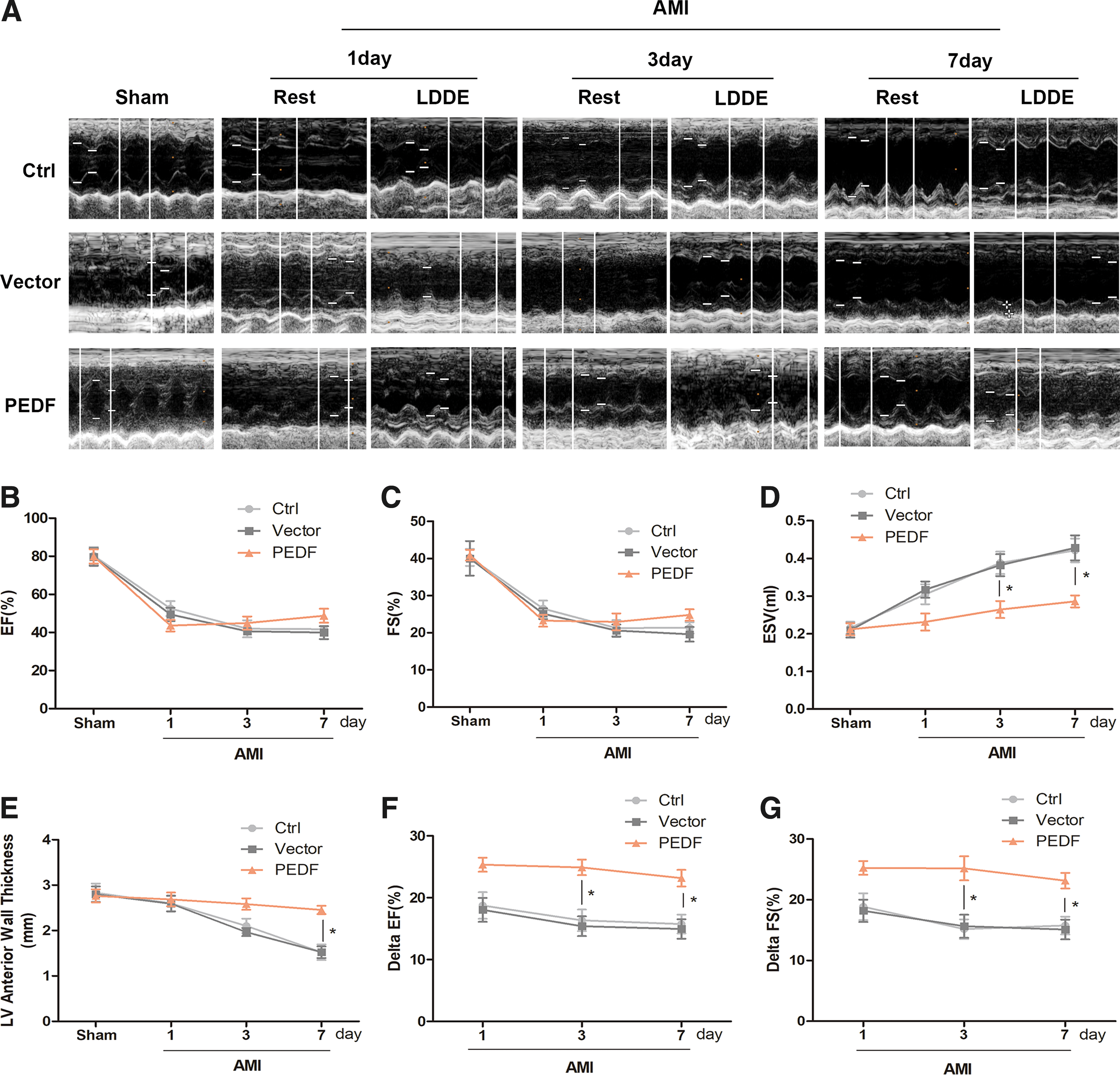
Echocardiography test at rest and dobutamine stress. Echocardiography was performed at rest and 10 min after injection of dobutamine (1 lg/g) in rats at 1, 3, and 7 days after AMI.
“Perfusion-metabolism mismatch” induced by PEDF overexpression in ischemic myocardium is revealed by PET
PET imaging has been considered as the gold standard for diagnosing the viability of myocardium under ischemic conditions. 27 The “perfusion-metabolism mismatch” imaged by PET has been clinically described as the hallmark of hibernating myocardium. 28 In this study, small-animal PET imaging of myocardial perfusion (traced using 13 N-NH3) and metabolic activity (traced using 18 F-FDG) was administered 3 days after AMI. Representative vertical long-axis images are shown in Fig. 4A. Relative perfusion defect volume (indicative of ischemia) and relative metabolic defect volume (indicative of loss viability) were calculated from polar map images. Quantification results showed that there was no significant difference in relative perfusion defect volume among all AMI rats. The relative metabolic defect volume was similar to that of perfusion defect in control group and vector group hearts, which is known as “perfusion-metabolic match.” However, the relative metabolic defect volume was dramatically lower than that of perfusion defect in PEDF group hearts, which is known as “perfusion-metabolic mismatch” (Fig. 4B). The relative volume of the mismatch (indicative of ischemia yet viable myocardium) was markedly increased in PEDF group hearts compared to control group or vector group hearts (Fig. 4C). In addition, plasma glucose and insulin levels were determined before PET imaging because they can influence myocardial glucose uptake and PET metabolic imaging. 29 The results showed that there was no significant difference among any of the groups (Fig. 4D and E). Thus, the interference of systemic serum glucose and insulin on glucose uptake was excluded. These results suggest that PEDF overexpression can maintain the viability of myocardium under acute ischemic conditions. Importantly, the “perfusion-metabolic mismatch” observed by PET imaging exhibited similar metabolic characteristics as those observed in hibernating myocardium.
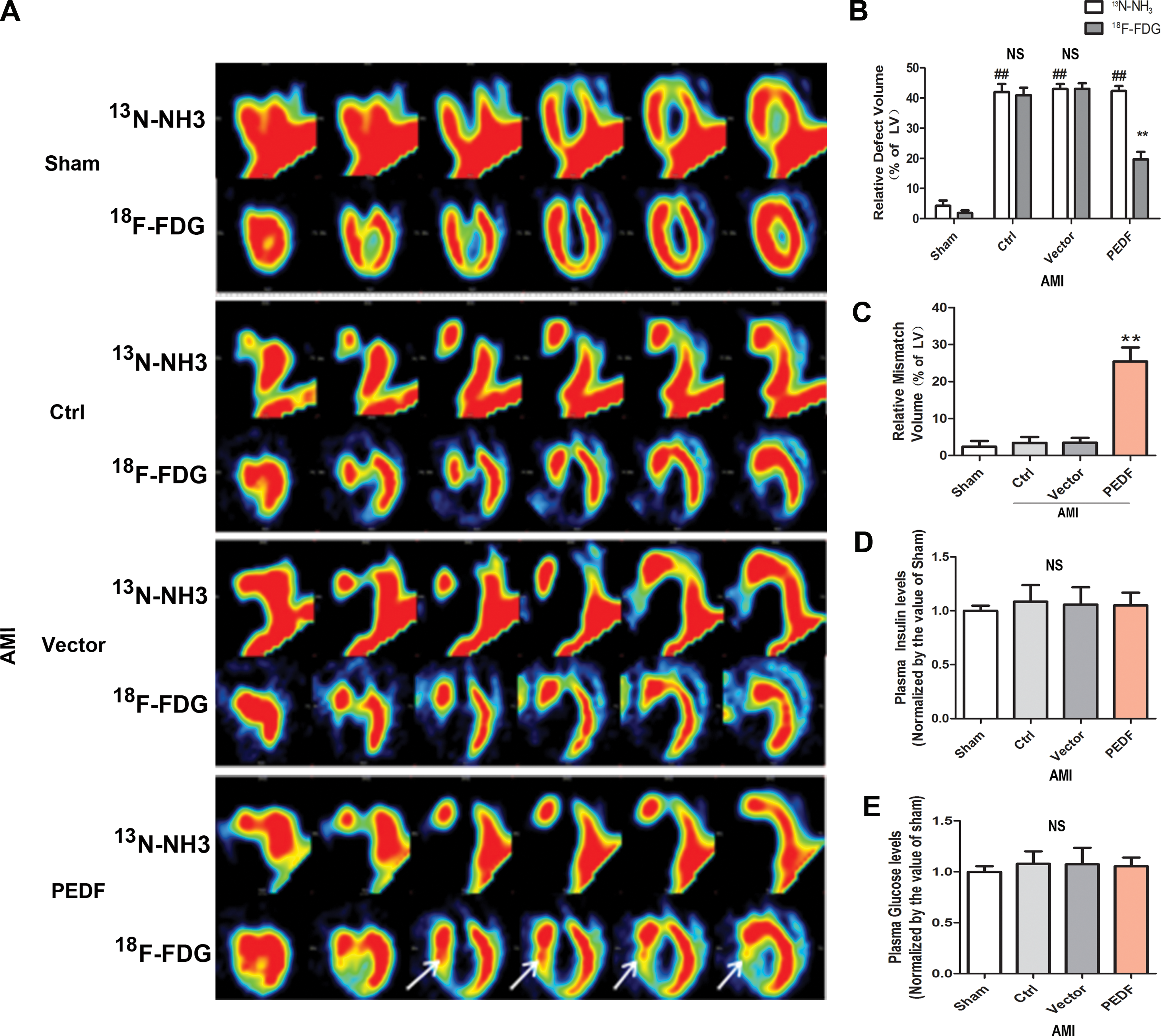
Positron emission tomography (PET) imaging and plasma glucose and insulin levels.
Ultrastructure changes observed by electron microscopy
Several ultrastructural features are used as pathological indicators of a diagnosis of hibernating myocardium. 30 To investigate the ultrastructural changes of acute ischemic myocardium induced by PEDF overexpression, electron microscopy was performed on biopsy sections obtained from LAD distribution after echocardiographic and PET studies. The representative images are shown in Fig. 5A. Examination of the sections revealed that no morphologic abnormalities were found in biopsy specimens obtained from sham group myocardium. However, extensive loss of myofilaments and large areas of nonspecific cytoplasm without glycogen were observed in control and vector groups. It is noteworthy that there were several unique ultrastructural changes in PEDF-LV transduced acute ischemic myocardium, including reserved numerous contractile materials, despite partial myolysis within viable cardiomyocytes, marked glycogen accumulation at the margins of the mitochondria and myofilaments. These features are similar to the typical changes that occur in hibernating myocardium. However, small and dense mitochondria like the structures in hibernating myocardium were not observed. 31 The mitochondria of PEDF-LV transduced acute ischemic myocardium were large, sparse, and irregular. These results demonstrate that despite subtle differences, a number of typical ultrastructural changes that are analogous to chronic hibernating myocardium were induced by PEDF overexpression in acute ischemic myocardium.
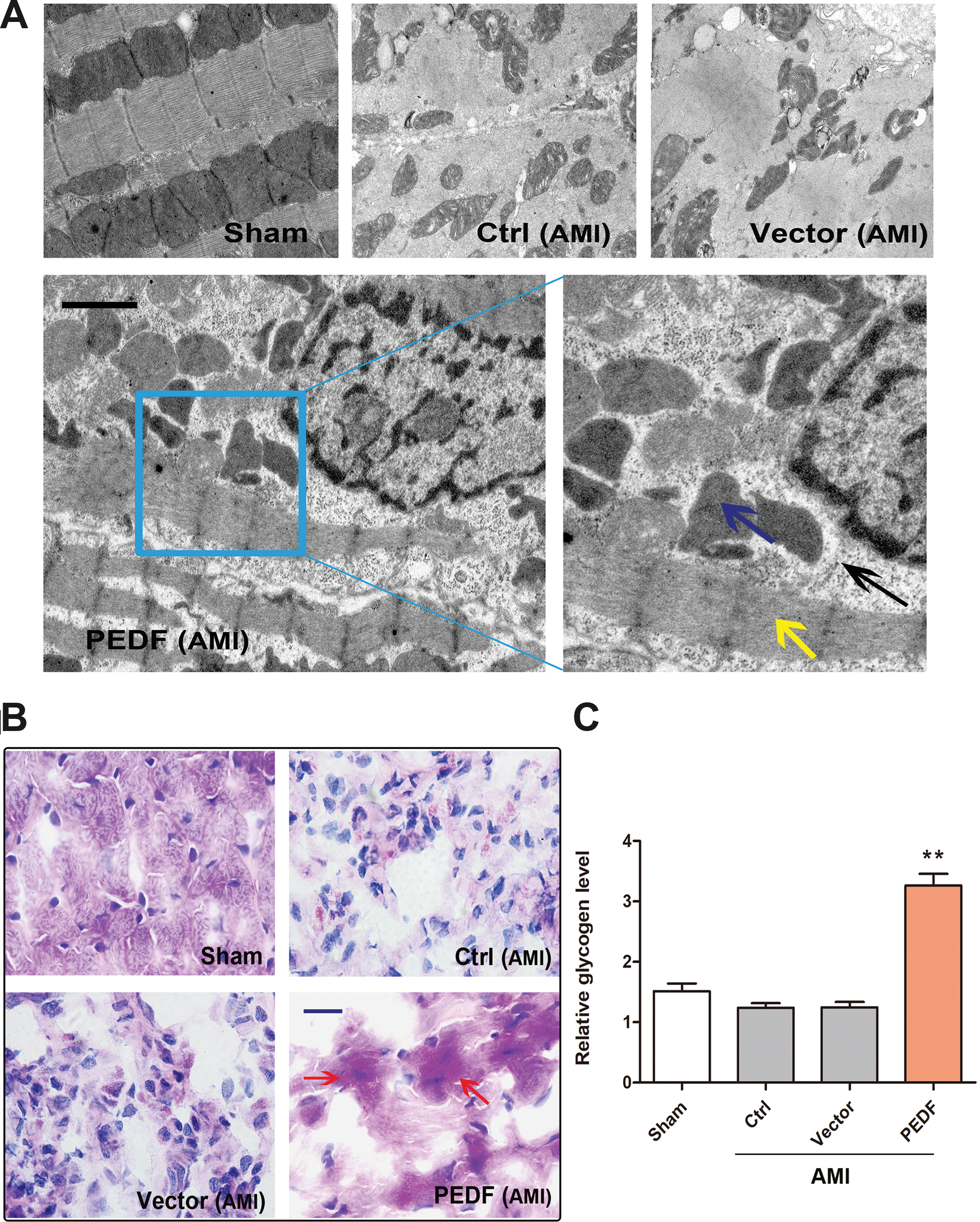
Ultrastructure change observed by electron microscopy and glycogen deposition demonstrated by Periodic acid–Schiff (PAS) staining.
PEDF overexpression increases glycogen deposition in ischemic myocardium
To maintain viability, hibernating myocardium increases energy reserves by enhancing glucose uptake and glycogenic deposition. 32 Glycogen deposition is considered to be histologically and metabolically important for identifying ischemic but viable myocardium. 33 To confirm the glycogen deposition in PEDF-LV transduced ischemic myocardium further, PAS staining and glycogen quantitative detection were performed 3 days after AMI. As shown in Fig. 5B, PAS staining revealed distinct glycogen accumulation in the PEDF overexpressed ischemic myocardium. Quantification demonstrated that the glycogen content in the PEDF group was markedly increased compared to the control group or vector group (Fig. 5C). These results imply that PEDF overexpression can increase glycogen deposition in acute ischemic myocardium, which is similar to hibernating myocardium.
PEDF overexpression protects cardiomyocytes against anoxic injury and increases glycogen deposition in anoxic cardiomyocytes
Cultured neonatal cardiomyocytes provide a useful model for understanding cardiovascular diseases. First, PEDF protein expression was examined in rat neonatal cardiomyocytes. Results indicated that the basal PEDF protein levels were high in normoxic neonatal cardiomyocytes and gradually decreased during hypoxic condition. Exogenous PEDF-LV transduction abolished the decrease in PEDF protein levels in anoxic cardiomyocytes (Fig. 6A and B). Then, the protective roles of PEDF overexpression on cultured rat neonatal cardiomyocytes were tested using RIP3 protein detection and CCK-8 assay. It was found that the levels of RIP3 significantly increased after 4 h of hypoxia, and cell viability was decreased in a time-dependent manner in relation to the duration of hypoxia exposure. Nevertheless, PEDF overexpression significantly reduced the RIP3 protein expression and alleviated the decrease in cell viability (Fig. 6C and D). These data demonstrated that PEDF overexpression protected neonatal cardiomyocytes against hypoxia-induced necrosis and maintained the viability of cardiomyocytes under hypoxia conditions. The PAS staining and glycogen quantitative detection showed that PEDF overexpression significantly increased the glycogen accumulation in anoxic cardiomyocytes, which was consistent with the results in vivo (Fig. 6E and F).
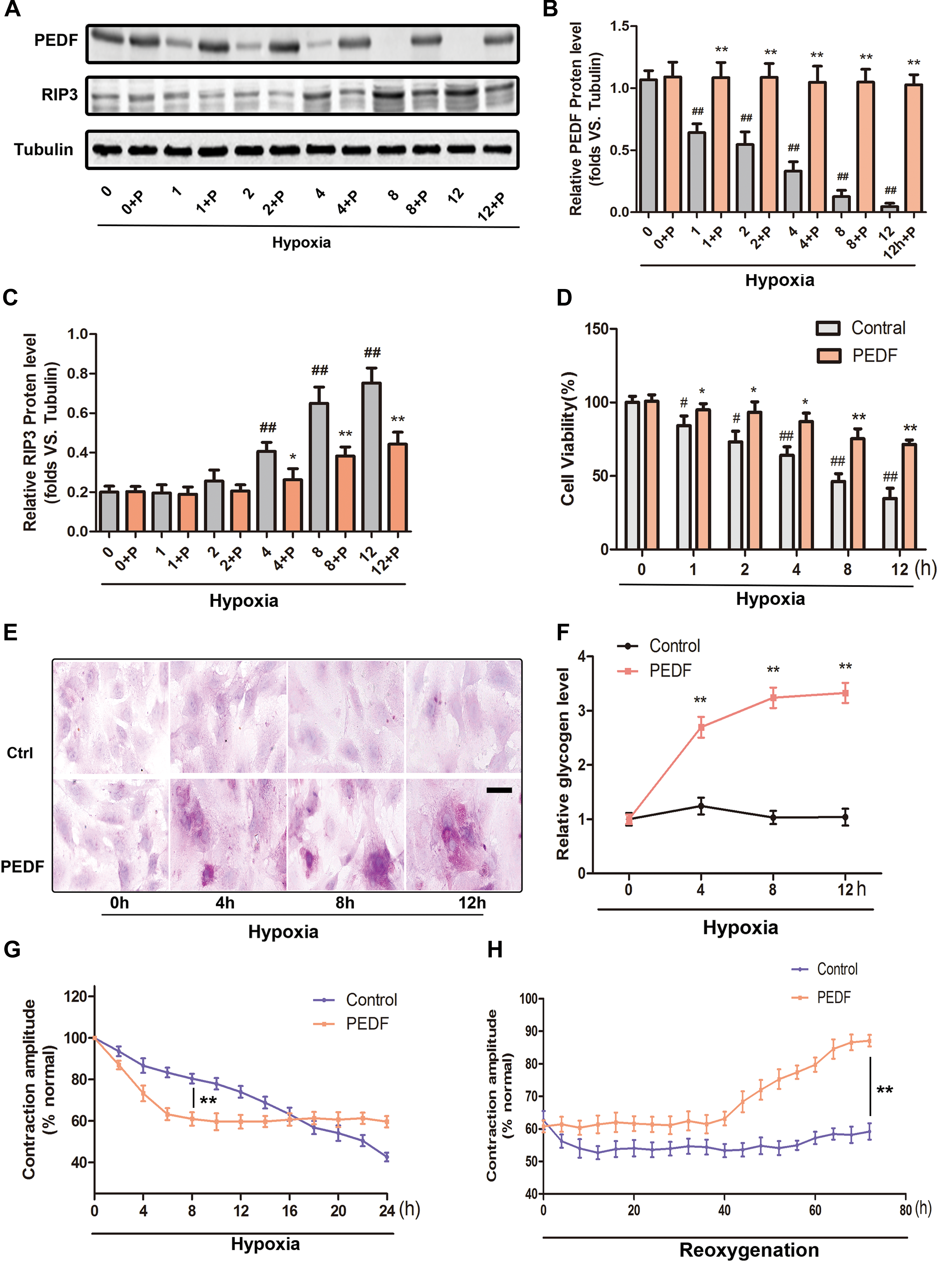
Cardioprotective effect of PEDF in neonatal cardiomyocytes and contractile recoverability of anoxic cardiomyocytes.
PEDF overexpression reduces the contractile function of anoxic cardiomyocytes but maintains their functional recoverability
The key feature of viable myocardium in hibernation is that its contractile dysfunction can return to normal (or near normal) after revascularization. 9 To investigate the effects of PEDF overexpression on the contractile function of anoxic cardiomyocytes, a video edge detection system was used to monitor single-cell contractile changes during hypoxia and after reoxygenation.
First, cardiomyocytes were exposed to hypoxia for 24 h with or without PEDF-LV transduction. As shown in Fig. 6G, the contractile function of cardiomyocytes from the two groups decreased as the duration of exposure to hypoxia increased. Moreover, PEDF overexpression produced a more rapid time-dependent reduction of cardiomyocyte contraction, which persisted for about 8 h. The contraction of control cardiomyocytes was persistently decreased with a gentler slope, and was similar to that of the PEDF group about 16 h after hypoxia. Subsequently, the cardiomyocytes were reoxygenated after 16 h of hypoxia (Fig. 6H). It was found that after a period of low functional state, the contraction of PEDF overexpressed cardiomyocytes showed a slow recovery response after reoxygenation. At 72 h of reoxygenation, the contraction of PEDF overexpressed cardiomyocytes recovered to about 85% of the normoxic value. However, there was no obvious functional recovery after reoxygenation in control cardiomyocytes. These results clearly suggest that PEDF overexpression could rapidly reduce the contractile function of cardiomyocytes in the early stages of hypoxia but retain their functional recoverability. The functional changes induced by PEDF overexpression are characteristic and highly suggestive of viable cardiomyocytes in a hibernating state.
Discussion
This study demonstrates that PEDF local overexpression can maintain the viability of acute ischemic myocardium. Furthermore, the viable state of acute ischemic myocardium induced by PEDF overexpression is extremely similar to the typical characteristics of chronic hibernating myocardium in function, metabolism, and ultrastructure. These results have the dramatic implication that PEDF may induce acute ischemic myocardium into a “hibernating-like” viable state.
PEDF is downregulated in both ischemic myocardium in vivo and anoxic cardiomyocytes in vitro. 13 Previous research has proven that delivering LVs carrying a target gene by using intra-myocardial injection is an effective method for modifying gene expression. 15,16 This study used this method to achieve a stable and sustained local overexpression of PEDF protein in ischemic myocardium. In addition, PEDF-LV transduction also realized PEDF overexpression in hypoxic cardiomyocytes. The cardioprotective effects of PEDF were verified both in vivo and in vitro once again, consistent with previous studies. Excitingly, in the process of evaluating the direct effect of PEDF overexpression on myocardial viability, some analogous changes to the typical characteristics in chronic hibernating myocardium occurred in PEDF overexpressed acute ischemic myocardium.
Hibernating myocardium has been described as “dysfunctional yet viable” myocardium following persistent regional hypoperfusion in myocardial territory. 7 By definition, contractile dysfunction is a requisite feature of hibernation. In this study, PEDF overexpression reduced the infarct size after AMI but did not increase the left ventricular function of rats, which indicated the possibility that acute ischemic myocardium protected by PEDF was also in a dysfunctional state. This proposition was further verified by in vitro contractile function tests, which indicated that PEDF overexpression reduced the contractile function of anoxic cardiomyocytes. Echocardiography and PET are commonly used to identify myocardial viability in clinical practice. 34 In the present study, lower ESV and a thicker left ventricular wall assessed by echocardiography at rest reflect myocardial viability. 25 LDDE in this study demonstrated the contractile reserve of the total heart rather than the ventricular wall movement in the PEDF-LV transduced region. However, PET imaging confirmed the existence of viable myocardium in the PEDF-LV transduced ischemia area. Hence, the contractile reserve demonstrated by LDDE was attributed to the viable myocardium created by PEDF overexpression in the ischemic area. These indicators show that PEDF overexpression may induce acute ischemic myocardium into a “dysfunctional yet viable” state, which is consistent with the hallmark of chronic hibernation. Moreover, the ultrastructural changes and glycogen deposition of the viable myocardium were similar to the histological characteristics of hibernating myocardium.
Despite of all these, there were some differences as well. First, the formation and survival conditions are different. Hibernating myocardium is an endogenous protective mechanism that enables myocardium to adapt to chronic ischemia. 7 The blood flow in hibernating myocardium is partly retained or even normal at rest, but the perfusion reserve is reduced. 35 In this study, a remodeling similar to hibernating myocardium in acute ischemic myocardium was induced by PEDF local overexpression. Since the blood flow was completely occluded, the energy source of survival myocardium may be glucose from intercellular space and residual collateral circulation. 36 In addition, structurally, the typical mitochondrial changes that exist in hibernating myocardium were not found in PEDF overexpressed acute ischemic myocardium. The number of mitochondria in PEDF overexpressed ischemic myocardium was significantly reduced, which is probably attributed to mitochondrial autophagy promoted by PEDF in ischemic cardiomyocytes. 37 In summary, the data presented here imply that PEDF overexpression may induce acute ischemic myocardium into a special viable state, which is similar to chronic hibernating myocardium. This viable state is carefully defined as a “hibernating-like” state.
Functional reversibility is considered as one of the most important and valuable characteristic of hibernating myocardium. 38 Multiple evidence suggests that patients with ischemic cardiomyopathy and the presence of viable myocardium in hibernation will gain increased prognostic benefits from revascularization compared to those with no hibernating myocardium. 18,39 LEED and PET are commonly used to predict functional recovery of the ischemic heart after reperfusion. 39 A large collection of data from non-randomized observational studies has confirmed their role in clinical practice over the past three decades. 40 In the present study, contractile reserve assessed by LDDE and “perfusion-metabolism mismatch” determined by PET were used to predict the functional recoverability of the myocardium in a “hibernating-like” state induced by PEDF overexpression. To investigate further, in vitro evidence was provided that PEDF overexpression preserved the functional recovery potential in hypoxic cardiomyocytes after reoxygenation. These led to the conjecture that PEDF overexpression in ischemic myocardium may prolong the “therapeutic window” after AMI and play an important role in functional recovery after revascularization (at least partly).
Limitations
The limitation of this study is the lack of direct demonstration of left ventricular functional reversibility in vivo after revascularization. The reason is that it is almost impossible to revascularize after 3 days of coronary artery ligation in this rat model. However, the contraction changes that were reported in vitro may be seen as “evidence” of left ventricular functional recovery, at least in part.
Conclusion
In summary, this study demonstrates that PEDF local overexpression may induce acute ischemic myocardium into a “hibernating-like” state and maintain its viability after AMI. This novel effect of PEDF presents an important clinical approach to prolong the “therapeutic window” after AMI and to enhance functional recovery after revascularization therapy. Importantly, this study may provide a novel therapeutic strategy for AMI.
Footnotes
Acknowledgments
We thank Weifeng Liu (MITRO Biotech Co. Ltd. NanJing, P.R. China) for the PET analysis. This work was supported by the grant from National Nature Science Foundation of China (81570242 and 81270173).
Author Disclosure
The authors disclose no conflict of interest.