
Abstract
Cystathionine β-synthase (CBS) deficiency is a recessive inborn error of metabolism characterized by extremely elevated total homocysteine (tHcy) in the blood. Patients diagnosed with CBS deficiency have a variety of clinical problems, including dislocated lenses, osteoporosis, cognitive and behavioral issues, and a significantly increased risk of thrombosis. Current treatment strategies involve a combination of vitamin supplementation and restriction of foods containing the homocysteine precursor methionine. Here, a mouse model for CBS deficiency (Tg-I278T Cbs−/– ) was used to evaluate the potential of minicircle-based naked DNA gene therapy to treat CBS deficiency. A 2.3 kb DNA-minicircle containing the liver-specific P3 promoter driving the human CBS cDNA (MC.P3-hCBS) was delivered into Tg-I278T Cbs−/– mice via a single hydrodynamic tail vein injection. Mean serum tHcy decreased from 351 μM before injection to 176 μM 7 days after injection (p = 0.0005), and remained decreased for at least 42 days. Western blot analysis reveals significant minicircle-directed CBS expression in the liver tissue. Liver CBS activity increased 34-fold (12.8 vs. 432 units; p = 0.0004) in MC.P3-hCBS-injected animals. Injection of MC.P3-hCBS in young mice, subsequently followed for 202 days, showed that the vector can ameliorate the mouse homocystinuria alopecia phenotype. The present findings show that minicircle-based gene therapy can lower tHcy in a mouse model of CBS deficiency.
Introduction
Cystathionine β-synthase (CBS) deficiency is the most common inborn error of sulfur amino acid metabolism. CBS catalyzes the conversion of homocysteine to cystathionine, and patients lacking CBS have a build up of homocysteine in extracellular fluids, including the blood and urine. Total homocysteine (tHcy) refers to the sum total of all forms of homocysteine containing both reduced and oxidized thiol groups. In human plasma, normal tHcy concentrations are generally between 5 and 15 μM, but in CBS-deficient patients at the time of diagnosis, a tHcy concentration of 100–200 μM is not uncommon. 1
Untreated CBS-deficient patients have a variety of clinical problems caused by elevated tHcy, including a greatly increased risk of thrombosis, osteoporosis, dislocated lenses, and behavioral and learning issues. 1 Treatment strategies focus on ways to lower patients' tHcy by nutritional and dietary means. 2 Initially, patients are generally given pharmacologic doses of pyridoxine that can lower tHcy to <50 μM in about half of cases. The mechanism of pyridoxine response is thought to work by increasing the intracellular concentration of pyridoxal phosphate, a key co-factor for the enzyme. For those patients who do not respond to pyridoxine, dietary methionine restriction and/or supplementation with betaine is recommended. However, compliance with a low-methionine diet is a major issue in caring for these patients. It is also important to point out that although treated patients do have fewer health issues than untreated patients, their tHcy values are generally still quite elevated. 3
Previously, a mouse model for CBS deficiency, Tg-I278T Cbs−/– was developed. 4 These mice contain a transgene cassette that expresses a patient-derived mutant CBS protein (p.I278T) under control of the zinc inducible MT-I promoter that allows the mice to survive the neonatal lethality associated with homozygosity for Cbs. Adult Tg-I278T Cbs−/– mice have a tHcy of ∼300 μM and have several phenotypes, including alopecia, loss of fat mass, reduced bone density, and shorter life-span. 5,6 As with human patients, these phenotypes can be ameliorated by restricting dietary methionine or supplementing with betaine. 7,8
Gene therapy is a promising avenue of treatment for many inherited disorders by directly addressing the genetic defect. Viral delivery of therapeutic DNA using adeno-associated virus (AAV) vectors has been used successful and is a promising approach primarily for liver-directed therapy. 9 Nevertheless, AAV vectors have some drawbacks, including immunogenicity, size limitations, insertional mutagenesis, expression stability, and cost. 10 Here, a non-viral system was explored utilizing minicircle DNA (MC-DNA) vectors delivered by hydrodynamic injection for experimental gene therapy in the mouse as a model organism. MC-DNA has the advantage of lacking bacterial DNA sequences, which stimulate inflammation and transgene silencing, 11,12 thus impeding the effectiveness of gene therapy.
This study created a MC-DNA vector expressing the human CBS gene and introduced it into Tg-I278T Cbs−/– mice by hydrodynamic tail vein (HTV) injection. It was found that this approach could lower tHcy and lessen the alopecia phenotype in these animals. The findings suggest that MC-DNA-based gene therapy may be a viable approach in treating CBS deficiency.
Methods
Minicircle plasmid construction
MC-DNA was generated, as described previously. 13,14 Briefly, a human CBS cDNA fragment containing a N-terminal FLAG-tag (1.7 kb) was subcloned into NruI/HpaI restriction sites of pMC.PKU2014 to generate a plasmid having the hepatic-specific regulatory element P3 driving human CBS. The resultant plasmid, pMC.P3-hCBS, was transfected into bacterial strain ZYCY10P3S2T, which contains an inducible bacteriophage ØC31 integrase and I-SceI homing endonuclease. This strain was grown in broth containing 0.01% L-Arabinose to induce MC-vector production. The resultant MC-DNA was isolated from bacterial lysate by using the Qiagen endofree plasmid purification kit (cat. no. 12362) according to the manufacturer's protocol. Purified MC.P3-hCBS vectors were quantified by UV spectroscopy (Nanodrop ND1000).
Mouse model
Tg-I278T Cbs−/– mice were generated as previously described. 5 In brief, male Tg-I278T Cbs−/– mice were mated with female Tg-I278T Cbs+/– mice in cages with water bottles containing 25 mM ZnSO4. All pups were genotyped between 10 and 14 days of age. At the time of weaning (around 30 days), mice were put in new cages with non-zinc water. The average age of the mice in this study was 132 days (range 77–200 days), and an equal number of male and females were used. Serum was collected from animals by retro-orbital bleed. The Fox Chase Cancer Center Institutional Animal Care and Use Committee approved all animal procedures.
Gene delivery and experimental schemes
Before injection, all mice had serum collected for baseline tHcy and methionine determination. MC.P3-hCBS was delivered into Tg-I278T Cbs−/– mice via a single HTV injection. Mice were rapidly injected (5–8 s) with a solution containing 10% of the animal's body weight. The solutions contained either saline (control) or 5 or 50 μg of MC.P3-hCBS DNA. Serum was prepared from blood collected from orbital sinus using a micro-capillary tube. All mice were euthanized by isoflurane overdose.
Alanine aminotransferase assay and histological analysis
Alanine aminotransferase (ALT) activity was measured in serum using a spectrophotometric assay following the manufacturer's protocol (cat. no. 700260; Cayman Chemical). For the liver and skin, hematoxylin and eosin (H&E) staining was performed from formalin-fixed tissue. Skin was removed from the back, abdomen, and face. Paraffin-embedded sections (5 μm) were stained with Masson's trichrome. Hairs and whiskers were removed manually, rinsed in water, and placed in mounting medium (Vector Laboratory). Images were taken with the micrometer under the microscope, and the diameter of the hairs and whiskers was measured.
Immunoblots
Tissue homogenates from the liver and kidney were prepared in 10 mM Tris-HCl (pH 7.5) supplemented with protease inhibitors (Roche). Lysate (25 μg) was separated by 4–12% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (Invitrogen) under reducing conditions and transferred to nitrocellulose. Blots were probed with FLAG antibody (A01428; GenScript), HA antibody (H9658; Sigma–Aldrich), rabbit anti-CBS sera for total CBS protein, 15 and actin antibody (A5441; Sigma–Aldrich) for loading control. Gel images were captured and quantified using the FluorChem SP system (Alpha Innotech).
Measurement of serum tHcy and tissue CBS activity
CBS activity in the tissue was measured using a Biochrom 30 amino acid analyzer, as described previously. 16 One unit of activity is defined as nanomoles of cystathionine formed per milligram of tissue lysate protein per hour. Serum tHcy (a sum total of free and disulfide-bonded homocysteine) and methionine levels were measured by using the Biochrom 30 amino acid analyzer, as performed previously. 5
Statistical analysis
Values are the mean ± standard error of the mean. Differences between two groups were analyzed by the Mann–Whitney U-test (unpaired and nonparametric test). Significance between more than two groups was determined using one-way analysis of variance followed by Tukey's multiple comparison tests employing GraphPad Prism v6.0 (GraphPad Software, Inc.). Statistical significance was accepted at a p-value of <0.05.
Results
HTV injection of MC.P3-hCBS minicircles
MC-DNA that has a FLAG-tagged human CBS cDNA driven by the P3 promoter was produced, as shown by the scheme presented in Fig. 1A. The P3 promoter is an artificial hepatocyte-specific promoter derived from a minimal transthyretin promoter combined with the HS-CRM8 element from the human Serpina1 gene. 14,17 The minicircle-producing plasmid, pMC.P3-hCBS, contains attB and attP sites flanking the expression cassette, thus allowing deletion of the bacterial sequences in the presence of ØC31 endonuclease.
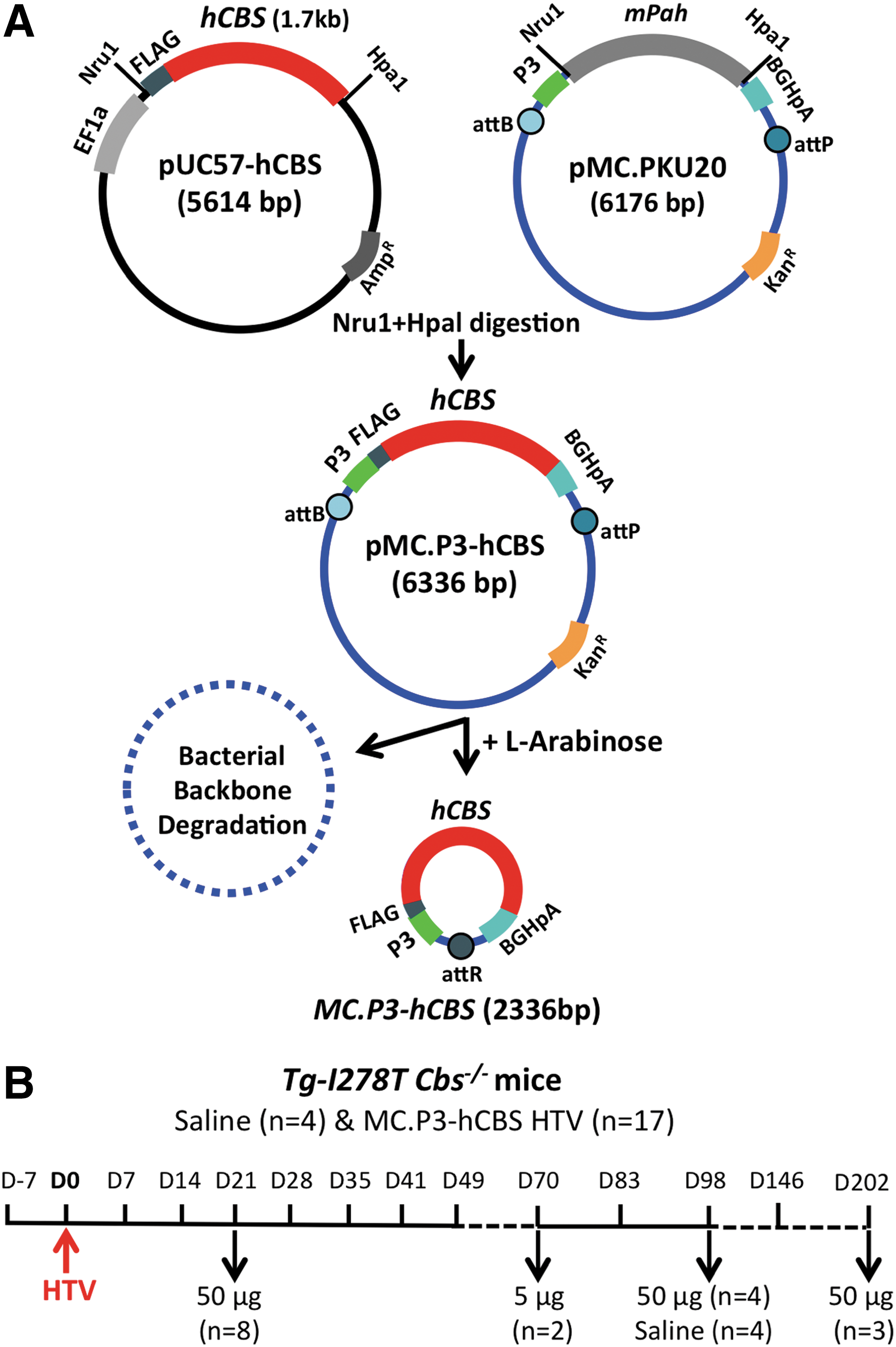
Minicircle-plasmid construction and experimental schemes.
Twenty-four Tg-I278T Cbs−/– mice were subjected to HTV injection. 18 Five mice were mock injected with saline, while the remaining mice were injected with either 5 μg (n = 2) or 50 μg (n = 17) MC.P3-hCBS DNA. Three of the mice (one injected with saline and two injected with 50 μg MC.P3-hCBS) died immediately after the injection, giving an 11% failure rate for HTV injection. The remaining mice had serum collected and analyzed for tHcy and were sacrificed according to the timeline shown (Fig. 1B).
HTV injection is thought to enhance DNA uptake into the liver through the fenestrated endothelium by transient hydrostatic pressure. 19 HTV injection is a standard experimental procedure that is used to deliver DNA into mouse hepatocytes with only transient liver damage. 20 Nevertheless, the study investigated whether this might cause liver injury that could confound the results. To explore this, ALT activities in the serum of eight mice (4 control and 4 injected with MC.P3-hCBS) both before (day −7) and after HTV delivery (days 7–70) were examined. Regardless of whether saline or MC-DNA had been injected, ALT activities were all in the normal range (<100 IU/L) during this period (Fig. 2A). In addition, H&E staining of the liver tissue showed no indication of liver damage in HTV-injected animals (Fig. 2B). These findings confirm that the HTV injection procedure does not cause persistent liver damage in Tg-I278T Cbs−/– mice.
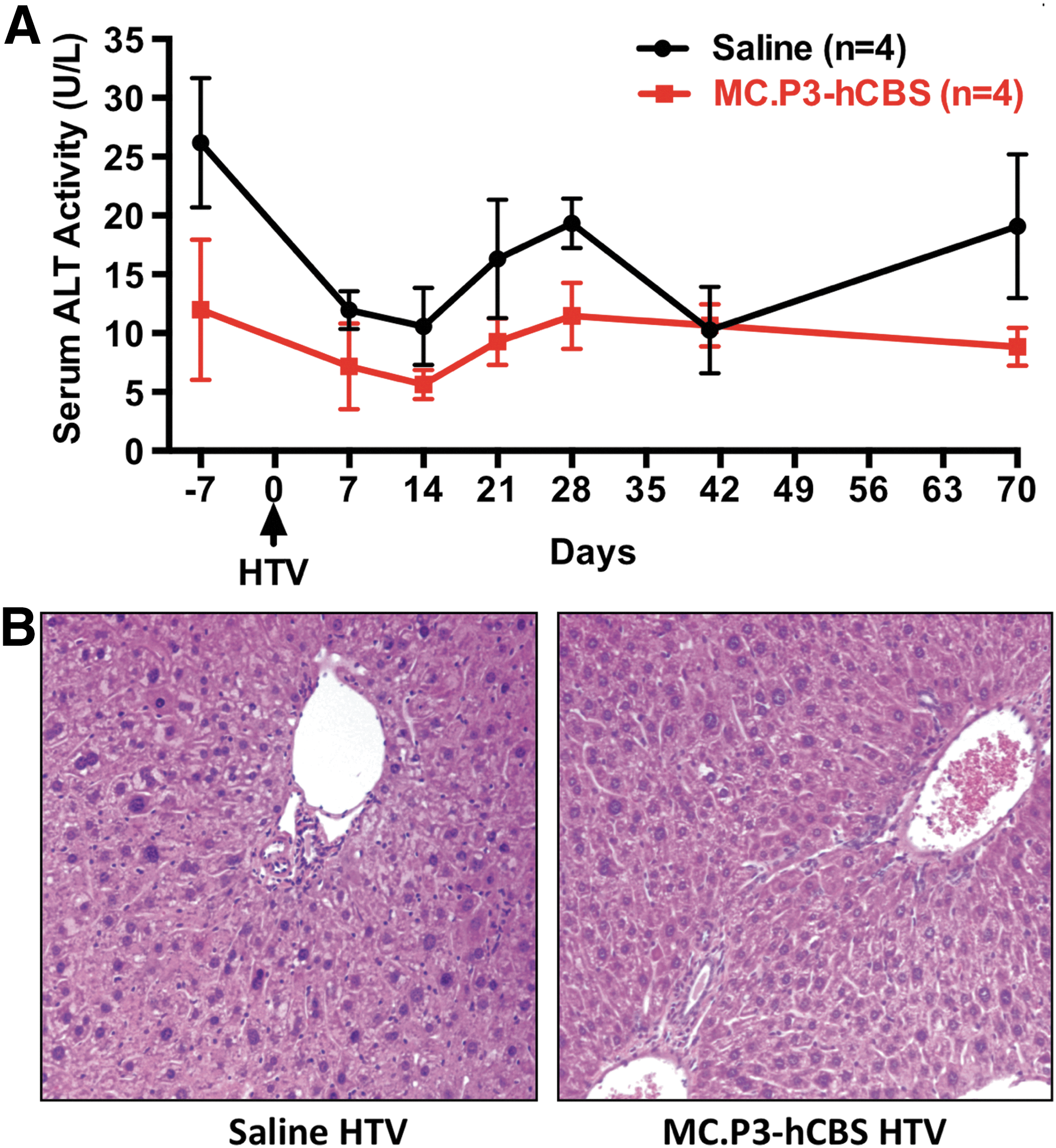
Lack of liver damage in CBS-deficient mice treated with hydrodynamic MC.P3-hCBS delivery.
MC.P3-hCBS delivery to CBS-deficient mice reduces serum tHcy
The 15 Tg-I278T Cbs−/– mice successfully injected with 50 μg MC.P3-hCBS had significantly lower serum tHcy from days 7 to 41 after injection compared to saline-injected controls. Maximal lowering of tHcy was observed at day 21, after which time levels began to rise (Fig. 3A). By day 98, tHcy were back to near pre-injection levels. At day 21, the mean tHcy was 131 μM, which is a 64% reduction from the tHcy at day 7 (p < 10–10). Serum methionine levels were not significantly altered by the 50 μg injection (Fig. 3B), but this lack of effect on serum methionine is not surprising, as CBS-deficient mice, unlike humans, do not show hypermethionemia. 5 For comparison, wild-type C57BL6 mice have mean serum tHcy of 5.4 μM and methionine of 66 μM. 21 In the two mice injected with the 5 μg dose, no lowering of tHcy or methionine was observed.

Effect of MC.P3-hCBS on total homocysteine (tHcy) and methionine.
MC-DNA human CBS is expressed in liver tissue
Liver CBS protein and activity was examined in eight of the 50 μg MC.P3-hCBS-treated mice that were sacrificed at day 21. Liver lysates were examined for CBS expression by Western blot analysis (Fig. 4A). Three different CBS antibodies were used: (1) a FLAG-tag antibody, which only detects MC-DNA-derived human CBS protein; (2) an anti-HA antibody, which detects the transgene encoded human I278T CBS protein present in the Tg-I278T mice; and (3) an antibody that detects both CBS proteins. All of the MC.P3-hCBS-injected mice showed strong MC-DNA-directed FLAG-tag CBS expression compared to controls. In some of the injected mice and one of the controls, low-level expression of the I278T CBS was also observed, presumably due to variable promoter leakage in the absence of the Zn inducer. However, the total CBS antibody suggests that the overwhelming amount of CBS protein is coming from the minicircle vector. It should be noted that the I278T CBS protein has only 2–5% of the enzyme activity of wild-type hCBS. So, even if present, it would contribute a miniscule amount of CBS activity.
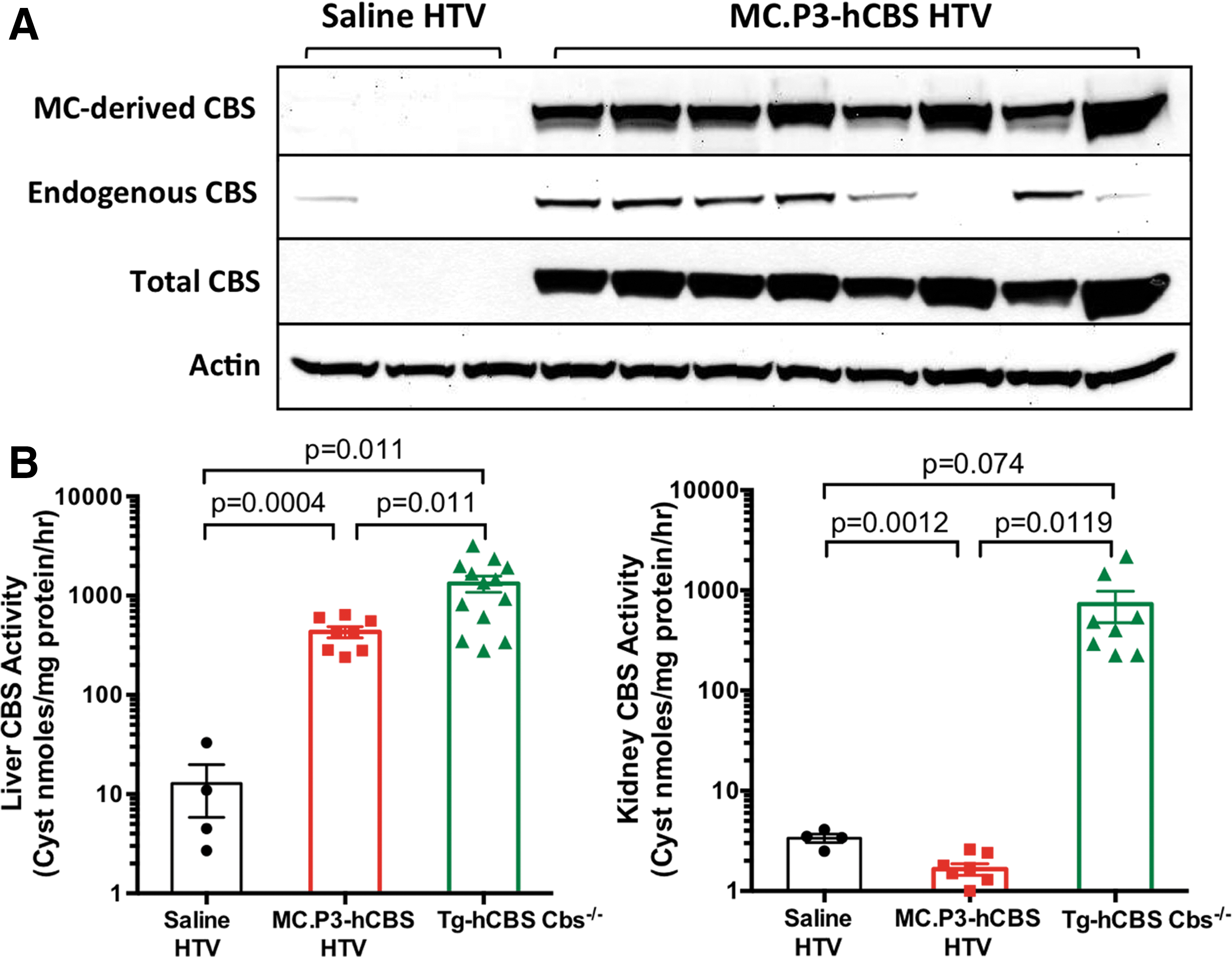
MC.P3-hCBS-directed CBS expression.
CBS enzyme activity was measured in both the liver and kidney tissues that normally express high levels of CBS activity (Fig. 4B). 16 In the liver of MC.P3-hCBS-injected mice, the mean CBS activity was 432 units compared to only 12 units in saline-injected mice. Activity in zinc-treated Tg-hCBS Cbs−/– mice was also measured, where the mean activity was 1,332 units. These mice expressed wild-type human CBS protein and had a mean tHcy of 20 μM. The threefold difference in activity between the MC.P3-hCBS group and the Tg-hCBS Cbs−/– group likely explains the higher tHcy levels in the MC.P3-hCBS-injected mice. No enhancement of CBS activity by MC.P3-hCBS treatment was observed in the kidney, confirming the specificity of the P3 promoter that has been observed previously. 14
Phenotype rescue by MC.P3-hCBS HTV injection
Three young mice (39 days old, 14.7 ± 0.8 g, one female and two males) injected with 50 μg MC.P3-hCBS were monitored for phenotypic changes, along with measurements of serum tHcy and Met levels over a period of 202 days. In these three mice, consistently lower levels of tHcy were observed compared to age-matched non-injected Tg-I278T Cbs−/– control mice (Fig. 5A), although the difference only met conventional statistical significance (p < 0.05) in three of the eight time points. However, the likelihood of observing the mean being lower at all eight time points considered against the null hypothesis that there was no difference between injected and control animals is highly significant (p = 0.0039). Examination of the area under the curve (AUC) shows that the MC.P3-hCBS-treated mice had an 18.1% reduction in tHcy exposure compared to untreated animals. Similar analysis for methionine showed that MC.P3-hCBS-treated mice had a 9.3% increase. At the end of the time period, the mice were euthanized, and liver CBS protein and activity were analyzed (Fig. 5B). All three mice had detectable levels of minicircle-derived CBS expression by Western blot. Two of three mice had liver activity levels elevated relative to day 98 saline-injected control mice, but the amount of activity was lower than that observed in the day 21 mice shown in Fig. 4B.
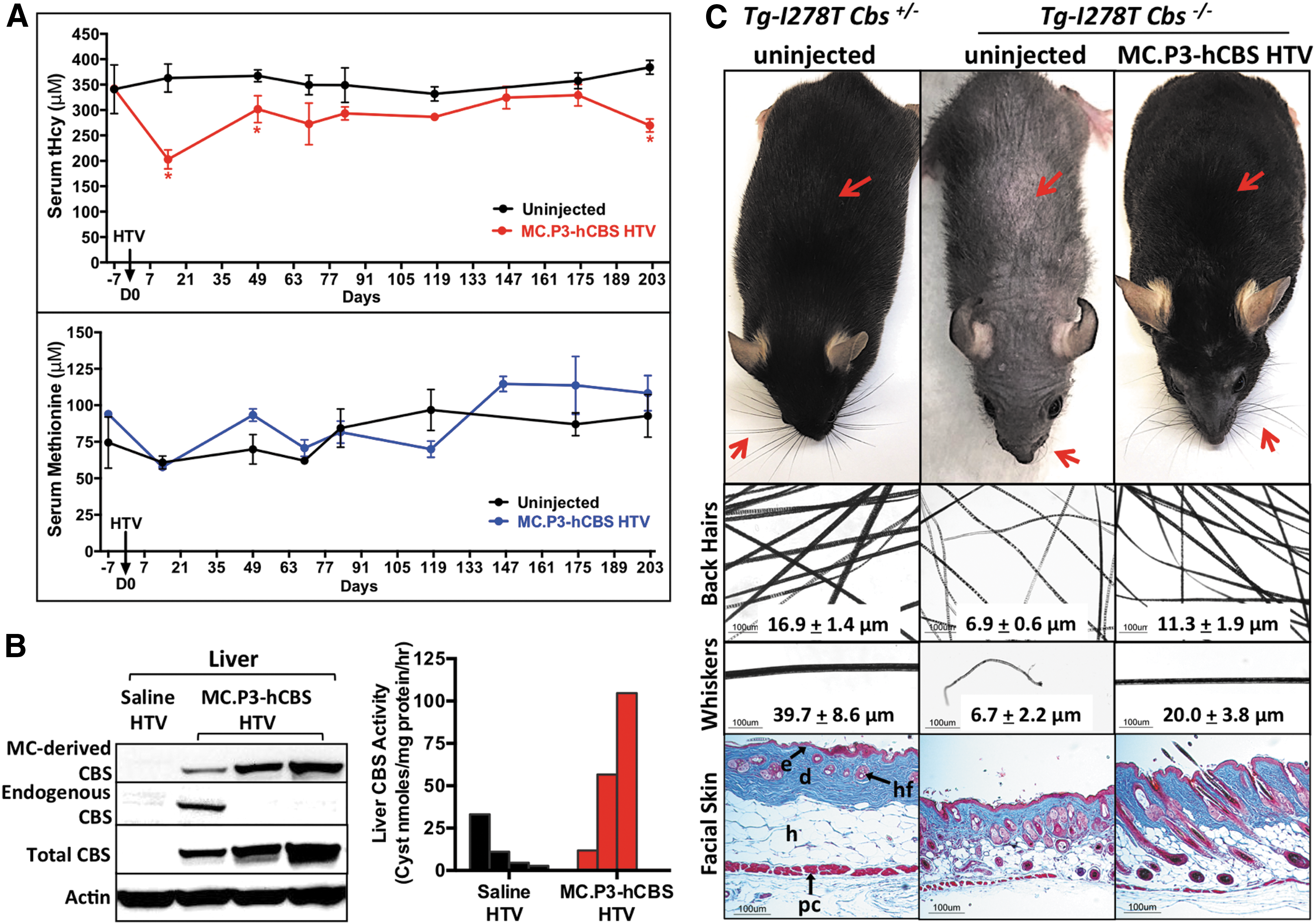
Rescue by MC.P3-hCBS on alopecia phenotype in Tg-I278T Cbs−/–
mice.
As the mice aged, it became apparent that the well-characterized alopecia phenotype of Tg-I278T Cbs−/– mice was greatly reduced (Fig. 5C). This was observed for both whiskers and body hair. Microscopic examination of the hairs showed that the hairs of the treated mice were significantly thicker compared to sex- and age-matched untreated Tg-I278T Cbs−/– mice. The mean diameter of the hair shaft on the back of the treated animals increased from 6.9 to 11.3 μm (p < 0.001), while the diameter of the whiskers increased from 6.7 to 20 μm (p < 0.012). However, it should be noted that the hair thickness was still less than that observed in control Tg-I278T Cbs+/– mice (tHcy ∼7 μM). 5 Microscopic examination of the skin revealed that Tg-I278T Cbs−/– mice had thinner skin compared to control Tg-I278T Cbs+/– animals, primarily due to a reduction in thickness of the dermal layer. However, in MC.P3-hCBS-treated animals, this was largely reversed.
Discussion
This study shows that a naked MC-DNA vector containing the wild-type human CBS cDNA driven by a liver-specific promoter (MC.P3-hCBS) can partially correct the metabolic and phenotypic defects observed in CBS-deficient mice. The MC-DNA vector lacks bacterial DNA sequences, which are eliminated by an inducible site-specific recombinase present in the bacterial strain used for plasmid propagation. The lack of bacterial sequence in the vector is thought to prevent transgene silencing, which is a major concern in gene therapy. 12 The MC-DNA was introduced by HTV injection, which involves injecting a MC-DNA-containing solution that is 8–10% of an animal's body weight in 5–7 s. Although this approach has been used successfully for delivery of non-viral gene therapy into mouse and rat livers for a variety of diseases, it is stressful for the animal, resulting temporary dysfunction of the cardiac system, significant liver expansion, and structural deformation. 22 In this study, 3/24 (11%) animals did not survive the procedure. However, in the animals that did survive showed no indication of liver damage 1 week later. While systemic hydrodynamic gene delivery may not be feasible in humans, local hydrodynamic gene delivery using image-guided catheterization of the hepatic vein has been successfully used in large swine. 23
The effectiveness of MC.P3-hCBS varied with the amount and with the length of time after injection. Maximal tHcy lowering was observed 21 days after injection with 50 μg injected. At this time, a tHcy of 64% from pre-injection levels was observed. Liver CBS activity at this time was about one-third of that observed in Tg-hCBS Cbs−/– mice, which express wild-type human CBS. As time went on, the effects slowly wore off, as judged by increasing serum tHcy. However, at day 202, two of the three mice tested still had liver CBS activity well above that of saline-injected control animals. In a previous study, PKU mice were treated with a MC-DNA vector using the P3 promoter to drive PAH gene expression. Here, blood phenylalanine (Phe) levels remained low for up to a year. 14 However, in this study, liver PAH enzyme activity was significantly lower at day 179 compared to day 4. The difference in behavior of tHcy and Phe over time may simply reflect that higher levels of CBS are needed for tHcy lowering relative to PAH. In the PAH study, the MC-DNA copy number was determined, and it was shown that the amount of DNA in the liver was proportional to the amount of DNA injected. In this study, tHcy lowering was observed with 50 μg but not with 5 μg of vector (although only two animals were injected with the lower amount). Given this, it is possible that injection with additional DNA template may cause longer and more effective tHcy lowering.
Interestingly, in the three animals that were followed till day 202, a significant correction of the alopecia phenotype was observed. The treated mice had significantly thicker hair, whiskers, and dermal layer of the skin compared to untreated control mice. Because multiple longitudinally collected tHcy data were available for each mouse, AUC analysis could be used to assess their total Hcy exposure. It is estimated that the overall exposure to tHcy was about 18% less in treated versus untreated animals. These data are consistent with other data generated by the authors' group, showing that the alopecia phenotype can be corrected with only a modest lowering of tHcy. 5,7,8
In summary, this study demonstrates that a MC-DNA vector delivered by HTV injection can be successfully used to treat a mouse model of CBS deficiency. The findings suggest that minicircle-vector-based gene therapy may be useful in the treatment of this disease.
Footnotes
Acknowledgments
This work was funded in part by grants CA06927 and R01GM098772 from the National Institutes of Health, and an appropriation from the Commonwealth of Pennsylvania. We also thank the Genomics and Laboratory Animal Facilities of Fox Chase Cancer Center for their assistance.
Author Disclosure
All the authors declare that they have no conflict of interest.