
Abstract
Infantile malignant osteopetrosis (IMO) is an autosomal recessive disorder characterized by nonfunctional osteoclasts. Approximately 50% of the patients have mutations in the TCIRG1 gene, encoding for a subunit of the osteoclast proton pump. Gene therapy represents a potential alternative treatment to allogeneic stem cell transplantation for IMO. The oc/oc mouse is a model of IMO characterized by a 1,500 bp deletion in the TCIRG1 gene, severe osteopetrosis, and a life span of only 3 weeks. Here we show that the osteopetrotic phenotype in oc/oc mice can be reversed by hematopoietic stem cell-targeted gene therapy with a clinically applicable lentiviral vector expressing a wild-type form of human TCIRG1 under the mammalian promoter elongation factor 1α short (EFS-hT). oc/oc c-kit+ fetal liver cells transduced with EFS-hT were transplanted into sublethally irradiated oc/oc mice by temporal vein injection 1 day after birth. A total of 9 of 12 mice survived long term (19–25 weeks) with evidence of tooth eruption, uncharacteristic of oc/oc mice. Splenocytes were harvested 19–25 weeks after transplantation and differentiated into osteoclasts on bone slices to assess resorption and on plastic to assess TCIRG1 protein expression. Vector-corrected osteoclasts showed human TCIRG1 expression by Western blot. CTX-I release relative to that mediated by oc/oc-derived osteoclasts increased 8–239-fold. Resorption pits on bone slices were observed for osteoclasts derived from 7/9 surviving transplanted oc/oc mice. Histopathology of the bones of surviving animals showed varying degrees of rescued phenotype, the majority with almost full correction. The average vector copy number per cell in the bone marrow was 1.8 ± 0.5. Overall, 75% of transplanted mice exhibited long-term survival and marked reversal of the osteopetrotic bone phenotype. These findings represent a significant step toward the clinical application of gene therapy for IMO.
Introduction
Osteopetrosis is a heterogeneous group of diseases characterized by impaired osteoclast function leading to an increase in bone mass. Osteopetrosis is generally classified based on inheritance into two subgroups: infantile malignant osteopetrosis (IMO) is an autosomal recessive disorder and the most severe form of osteopetrosis, whereas autosomal dominant osteopetrosis generally is a milder form of the disease. 1 –3 IMO is a rare disease with an incidence of 1:250,000 births 4,5 and mutations in the TCIRG1 gene account for 50% of all IMO cases. 5 –7 The TCIRG1 gene encodes for the a3 subunit of the osteoclastic V-ATPase, essential for the establishment of an acidic environment in the osteoclast resorption area. IMO patients with TCIRG1 mutations have an increased or normal number of osteoclasts that are unable to resorb bone. 8 As a result, they have dense bones, suffer from severe hematologic deficits with bone marrow (BM) failure and hepatosplenomegaly, and they develop blindness and other neurologic abnormalities due to compression of the cranial nerves. 9 Around 70% of patients die before the age of 6, mainly due to recurrent infections. 9 The only treatment for IMO is hematopoietic stem cell (HSC) transplantation (HSCT), preferably performed as early as possible after diagnosis. HSCT may restore the resorptive function of osteoclasts as they are derived from HSCs via fusion of monocytes. 10,11 HSC-targeted gene therapy for IMO has the potential to reverse the phenotype without some of the complications associated with allogeneic HSCT. 12
A mutation in Tcirg1 is also the cause of osteopetrosis in the oc/oc mouse model, which closely resembles the human disease; the features of this severe model of osteopetrosis include small size, lack of teeth, increased bone density, absent BM spaces, and a short life span of 3 weeks. 13 We have previously demonstrated the rescue of the oc/oc mouse phenotype with gene therapy utilizing a gammaretroviral vector-mediated gene transfer of mouse Tcirg1 cDNA into mouse HSCs. 14 However, gammaretroviral vectors have been associated with a risk of insertional mutagenesis at transcription start sites, 15 potentially resulting in leukemia. 16,17 In addition, the incorporation of weaker cellular promoters, such as those being used in the X-linked severe combined immunodeficiency (X-SCID) and adenosine deaminase deficient severe combined immunodeficiency (ADA-SCID) trials 18 –21 (as opposed to strong viral promoters), can increase therapeutic safety. We thus created a self-inactivating (SIN) lentiviral vector (LV) suitable for use in a clinical setting, a vector with the mammalian promoter elongation factor 1α short (EFS) and without a reporter gene. We recently demonstrated 22 that this clinically applicable LV-EFS-human TCIRG1 vector (EFS-hT) was able to correct the function of IMO osteoclasts ex vivo, as well as in vitro, by transplantation of gene therapy-corrected CD34+ cells from IMO patients into NSG-mice and generation of osteoclasts from isolated putative stem cells after several months. In the present study, we further validated the vector by transplanting oc/oc HSCs transduced with EFS-hT into neonatal oc/oc mice and assessed their long-term survival and rescue of the osteopetrotic bone phenotype.
Materials and Methods
Mice
Two pairs of oc/+ mice (JAX strain no. 000230) were obtained from the Jackson Laboratory (Bar Harbor, ME) and maintained in the conventional animal facility at the Biomedical Center, Lund University. All experiments were performed according to protocols approved by the local ethics committee. oc/+ mice appeared normal and were phenotypically indistinguishable from wild-type (WT) littermates.
Harvest and enrichment of hematopoietic cells from fetal liver and spleen
On embryonic day 14.5, pregnant mice were sacrificed, embryos were removed, fetal livers (FLs) were dissected out, and single-cell suspensions were prepared by drawing liver cells through a 23-gauge needle followed by filtering through a 50 μm cell strainer. Individual FLs were genotyped by lysing a cell sample and running a PCR as previously described. 14 To enrich for stem and progenitor cells, c-kit+ cells were separated magnetically using magnetic-activated cell separation columns (Miltenyi Biotec, Bergisch Gladbach, Germany). For transplantation experiments, the c-kit+ FL cells were cultured in serum-free expansion medium StemSpan medium (StemCell Technologies, Vancouver, BC) with the following recombinant cytokines (100 ng/mL): mouse stem cell factor, human Flt3L, and human thrombopoietin, all from R&D Systems (Minneapolis, MN).
Vectors, viral production, and transduction of cells
All the vectors used in this study are SIN LVs with a pRRL backbone (Fig. 1). The rescue vectors used for transduction of the transplanted cells (EFS-hT and EFS-mT) contain the WT human TCIRG1 or mouse Tcirg1 cDNA, respectively, under the EFS promoter. The control vector expresses green fluorescent protein (GFP) under the EFS promoter (EFS-G). LVs were produced by transient transfection of the vector plasmids into 293T cells along with a packaging plasmid (pCMV ΔR8.91) and an envelope plasmid (VSV-G pMDG). Supernatants were concentrated by ultracentrifugation and titers were determined by flow cytometry (EFS-G) or quantitative PCR for vectors lacking GFP (EFS-hT, EFS-mT). Transductions were carried out in 24-well plates coated with Retronectin (Takara Bio, Otsu, Japan). A short transduction protocol was developed to allow efficient transduction while maintaining the stem/progenitor nature of the c-kit+ population for transplantation. The c-kit+ FL cells were isolated and transduced overnight with a multiplicity of infection (MOI) 10; the following day the second transduction was performed with MOI 10 for 6 h, after which the cells were frozen. The total culture time of the cells before being thawed for transplantation was ∼30 h.
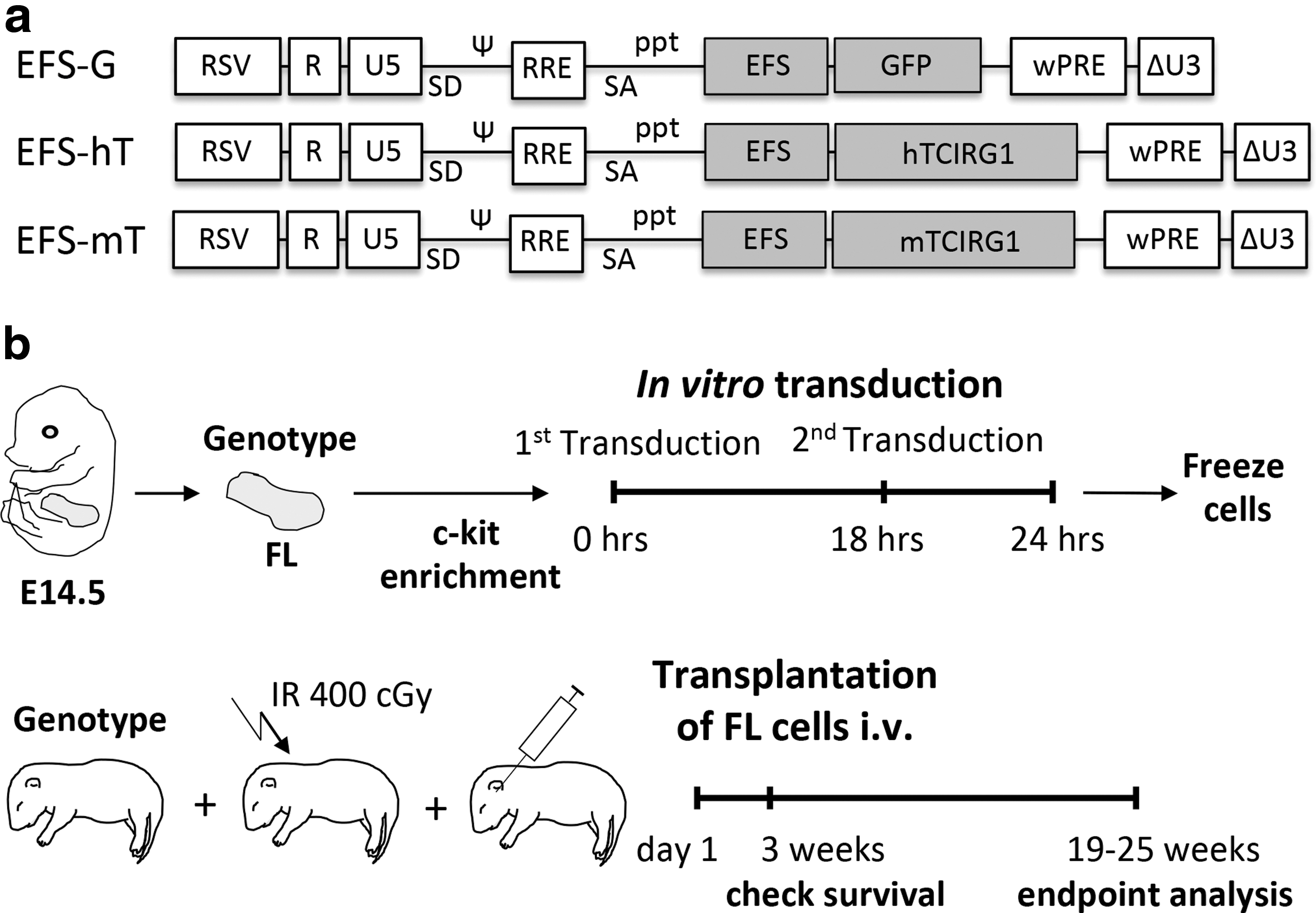
Schematic representation of the lentiviral vector design and experimental outline. The therapeutic vectors contain the human or the murine version of TCIRG1, whereas the control vector contains the marker gene GFP. For all vectors, the expression is driven by the EFS promoter
Transplantations
One-day-old pups from oc/+ breeding pairs were irradiated with 400 cGy, genotyped using DNA extracted from the tip of the tail, and transplanted intravenously (IV) via the temporal vein 6 h after irradiation with 2 × 106 freshly thawed c-kit+-transduced FL cells in 20 μL phosphate-buffered saline. Transplanted oc/oc mice were fed DietGel Recovery (ClearH2O, Portland, ME) food due to lack of tooth eruption or dental abnormalities in the mice where teeth erupted. Because of abnormal weaning, teeth had to be kept short by cutting. After 19–25 weeks, the mice were weighed, sacrificed, and then splenocytes, BM cells, and bones were harvested for analysis.
Osteoclastogenesis
Osteoclasts were generated ex vivo from splenocytes isolated from 5- to 6-month-old transplanted mice and 2-week-old oc/oc control mice. The cells were seeded into 96-well plates on bovine cortical bone slices at a density of 1.5–2.5 × 105/well for resorption assays and into 24-well plates on plastic at a density of 0.6 × 106/well on plastic for Western blot. Resorption assays were performed with 5–6 technical replicates per condition, and only mean data from each condition were used for analysis. The cells were incubated at 37°C and 5% CO2 in alpha-Minimum Essential Medium (αMEM) containing 10% fetal bovine serum, 100 units/mL penicillin, 100 μg/mL streptomycin, and 388 μg/L thymidine. They were expanded for 3 days in the presence of 50 ng/mL human macrophage colony-stimulating factor (M-CSF) and were differentiated for an additional 10 days in the presence of 50 ng/mL human M-CSF and 100 ng/mL mouse RANKL, both from R&D Systems (Minneapolis, MN), with medium changes every 2–3 days. After 13 days, the cells were either fixed in 4% formaldehyde for further analyses or lysed for Western blot analysis. Resorption was assessed by measuring the release of c-terminal type I collagen (CTX-I) fragments into the media on day 13 and the formation of resorption pits. Osteoclastogenesis was assessed by tartrate-resistant acid phosphatase (TRAP) activity in the media at every medium change.
Western blot
Cells were lysed in radioimmunoprecipitation assay (RIPA) buffer. Protein concentrations were measured using the Bio-Rad Protein Assay Kit II (Bio-Rad, Hercules, CA). Three to 13 μg of total protein in sodium dodecyl sulfate (SDS) sample buffer was separated by gel electrophoresis in an SDS-polyacrylamide gel electrophoresis 4–12% gradient gel followed by blotting onto a nitrocellulose membrane. Membranes were then blocked in tris-buffered saline Tween 20 (TBST) with 5% skim milk powder for 1 h at room temperature, followed by incubation with a primary antibody overnight at 4°C in TBST with 5% skim milk powder using the following antibody dilutions: mouse monoclonal anti-TCIRG1 (catalog no. H00010312-M01A; Abnova, Taipei, Taiwan) 1:500–1:1,000, rabbit polyclonal anti-TCIRG1 (catalog no. ab139812; Abcam, Cambridge, United Kingdom) 1:1,000–1:2,000, and rabbit polyclonal anti-p38 MAPK (catalog no. 9212; Cell Signaling Technology, Danvers, MA) 1:1,000. The blots were then washed 3 × 10 min and incubated with the corresponding horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature or at 4°C followed by 3 × 10-min washes in TBST buffer. Blots were developed using ECL Western Blotting Reagents (GE Healthcare, Waukesha, WI).
TRAP activity measurements
Media from resorption assays were diluted as needed with ultrapure water to a final volume of 20 μL with an optical density (OD) of less than 2.5 in the analyzed samples. The diluted samples were incubated with 80 μL freshly prepared reaction buffer (250 mM acetic acid, 0.125% Triton X-100, 250 mM NaCl, 2.50 mM ethylenediaminetetraacetic acid, 6.25 mM ascorbic acid, 25.00 mM disodium tartrate dihydrate, 6.06 mM 4-nitrophenyl phosphate disodium salt hexahydrate, pH 5.5) at 37°C for 1 h in the dark and the reaction was then stopped by adding 100 μL of 300 mM NaOH. Absorbance was measured at 405 nm with 650 nm as a reference using a VersaMax (Molecular Devices, Sunnyvale, CA) plate reader and the measured ODs were adjusted for the samples' dilution factors.
CTX-I
The release of CTX-I from resorbed bone slices was determined using the CrossLaps for Culture kit (IDS, The Boldons, United Kingdom), which was used according to the manufacturer's instructions.
Resorption pit formation
Resorption pits on the fixed bone slices were visualized by removing the osteoclasts using RIPA buffer and scrubbing with a cotton swab followed by rinsing with ultrapure water and staining with hematoxylin for 7 min. The stained bones were rinsed with ultrapure water, and excess dye was removed by scrubbing the bones with a cotton swab. Digital micrographs were obtained using a 10× objective and an Olympus DP71 digital camera mounted on an Olympus IX-70 microscope using the Olympus cellSens software (Olympus, Center Valley, PA).
Histopathology
Bones were dissected from the mice followed by fixation in 4% formaldehyde, decalcification in 20% modified formic acid (Histolab, Copenhagen, Denmark), neutralized in 5% sodium sulfate, dehydrated with 70% ethanol, and embedded in paraffin. Slides containing 5-μm-thick sections were stained with hematoxylin and eosin (H&E) and mounted with coverslips using Pertex. Sample and slide preparations were performed by Red Glead Discovery (Lund, Sweden).
Vector copy number
Vector copy number (VCN) analysis was performed on whole BM genomic DNA from samples harvested from mice 19 to 25 weeks after transplantation. The mean VCN per cell was determined by quantitative reverse transcription-PCR. Samples were measured in triplicate using 100 ng of genomic DNA. Primers for the woodchuck hepatitis virus posttranscriptional regulatory element of the vector were used to determine the amount of viral sequences, which was further normalized to a genomic reference sequence of the Ptbp2 gene. A serial dilution of a plasmid standard containing both sequences was measured in parallel to perform an absolute quantification. A cell line clone with predetermined VCN was used as an interplate calibrator.
Statistics
Comparison of survival curves was performed using the log-rank test. Weight data were analyzed with the Kruskal–Wallis test. Background biomarker measurements, obtained from cultures of bone slices without added cells, from the final day (CTX-I) or every day of the ex vivo resorption assay (TRAP) were subtracted from the sample measurements and the technical replicates were averaged. CTX-I data were normalized to the mean of the untreated oc/oc condition of the respective resorption assay and pooled. TRAP data were either pooled without normalization and plotted for every time point or used to calculate the area under the curve of each culture, followed by normalization to the untreated oc/oc of each respective assay and pooling of the data. The normalized pooled data were then analyzed using the Kruskal–Wallis test with Dunn's multiple comparisons test against the untreated oc/oc condition. Statistical analysis was performed using GraphPad Prism v7.01 for Windows (GraphPad Software, La Jolla, CA). Statistical significance was considered to be p < 0.05. In figures, statistical significance is indicated with *p < 0.05 and **p < 0.01. The data plotted in the graphs represent each individual mouse/osteoclast culture, the mean, and the standard errors of the mean where applicable.
Results
Neonatal transplantation of oc/oc FL cells transduced with EFS-hT results in long-term survival of oc/oc mice
The aim of this study was to determine if oc/oc FL cells transduced with the EFS-hT LV, expressing human TCIRG1 (hTCIRG1) under the EFS promoter (Fig. 1a), and transplanted IV into neonatal oc/oc mice could rescue these animals from fatal progression of osteopetrosis. c-kit+ FL cells were transduced and transplanted into recipients on day 1 after birth (Fig. 1b). Two treatment groups were used in this study: oc/oc mice transplanted with oc/oc cells transduced with either the EFS-hT LV (oc/oc tx oc/oc-EFS-hT, n = 12) or the EFS-mT LV (oc/oc tx oc/oc-EFS-mT, n = 8), expressing mouse TCIRG1 (mTCIRG1) under the EFS promoter, to test the efficacy of both vectors. The other two groups in the study represent positive controls: WT mice or oc/oc mice transplanted with WT cells transduced with the EFS-G LV, expressing GFP under the EFS promoter (WT tx WT-EFS-G, n = 6, and oc/oc tx WT-EFS-G, n = 6, respectively). All animals were assessed for survival after 3 weeks, the normal life span of oc/oc mice, and for up to 19–25 weeks, the endpoint for our analysis. All oc/oc mice transplanted with WT cells survived long term (6/6). In the oc/oc tx oc/oc-EFS-mT group, only 25% (2/8) mice survived to the endpoint, whereas in the oc/oc tx oc/oc-EFS-hT group 75% (9/12) survived long term (Fig. 2a). The size of oc/oc mice is severely reduced compared with littermates of the same age. All three groups of transplanted oc/oc mice increased in weight over time but did not reach the average WT group weight by the endpoint (Fig. 2b). Another distinguishing feature of the oc/oc mice is their lack of teeth. Tooth eruption was observed in all mice of the oc/oc tx WT-EFS-G group (Fig. 2d) and the oc/oc tx oc/oc-EFS-hT group (Fig. 2f), although with abnormalities compared with WT mice (Fig. 2c), whereas tooth eruption was absent in the two surviving mice of the oc/oc tx oc/oc-EFS-mT group (Fig. 2e). Based on these data, and due to EFS-hT being the clinically relevant vector, further analyses were only conducted with the vector expressing human Tcirg1.
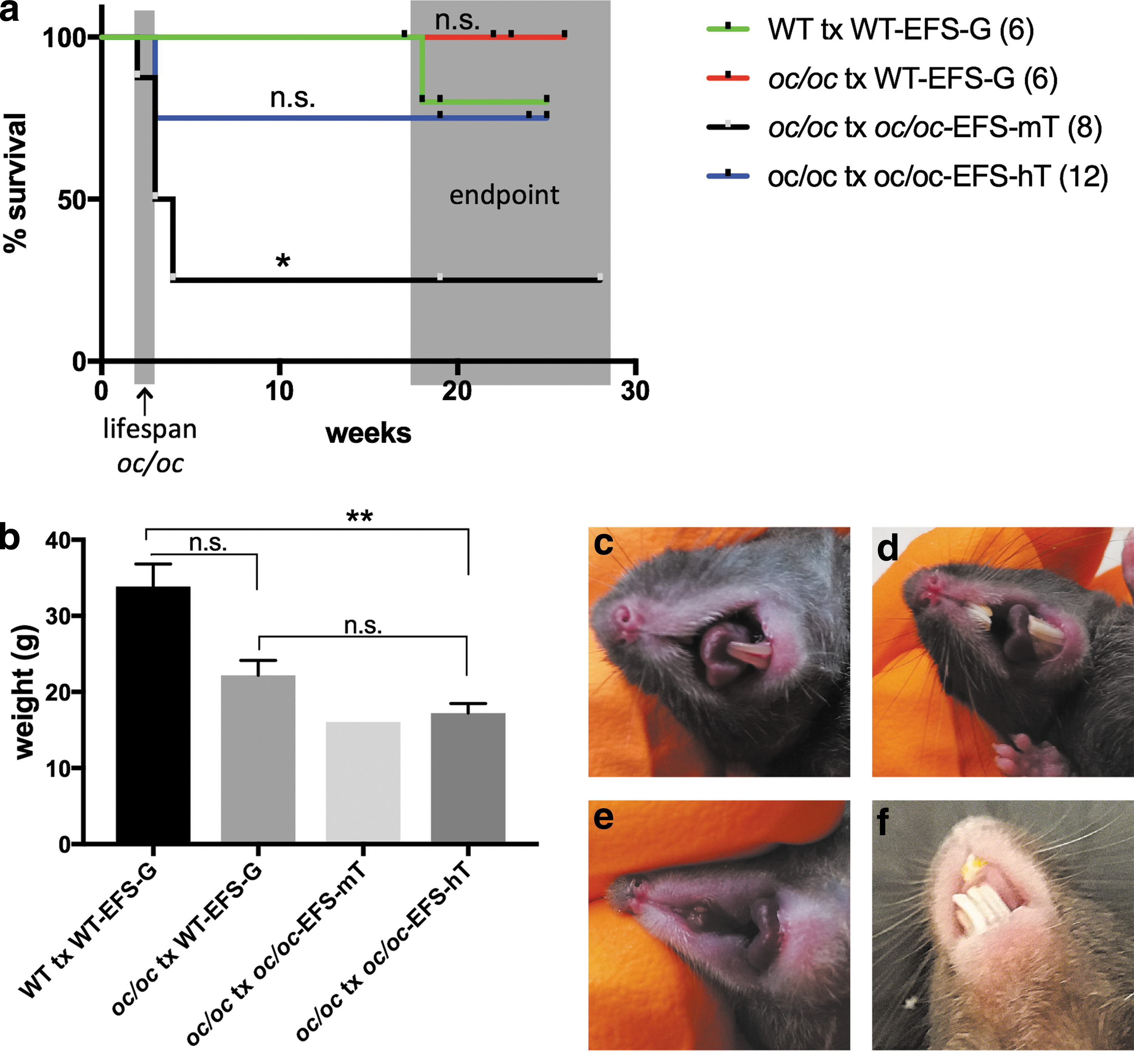
Long-term survival of 9/12 oc/oc mice transplanted with oc/oc FL cells transduced with the EFS-hT vector. Transduced FL cells were transplanted IV into irradiated newborn mice. Kaplan–Meier curves show the survival of transplanted mice over time
Splenocytes harvested from transplanted oc/oc mice express hTCIRG1 and can form bone-resorbing osteoclasts ex vivo
Splenocytes were harvested from WT and oc/oc mice 19–25 weeks after transplantation, the mononuclear fraction was isolated, seeded on plastic or bone slices, and differentiated into osteoclasts as described. Western blot analysis performed on lysates from mature osteoclasts detected an hTCIRG1 band in all mice transplanted with EFS-hT-transduced cells but not in untreated oc/oc controls or WT mice (Fig. 3a). Cells from transplanted oc/oc mouse no. 4 were not assessed by Western blot due to an insufficient amount of cells (Table 1). mTCIRG1 presence was verified in the WT mice and was not detectable in untreated oc/oc mice. Osteoclastogenesis was verified by assessing TRAP activity in the media, and the ability of osteoclasts to resorb bone was evaluated by measuring the release of CTX-I into the media. TRAP activity increased over time as expected in all cultures (Fig. 3b and Supplementary Fig. S1), but the cells from oc/oc mice as well as one oc/oc tx oc/oc-EFS-hT mouse had earlier osteoclastogenesis than the WT mice and the other oc/oc tx oc/oc-EFS-hT mice (Supplementary Fig. S1). Relative to the oc/oc group, there was a trend toward a decrease in overall TRAP activity in the analyzed specimens from the oc/oc tx oc/oc-EFS-hT group (Fig. 3b), but it was not statistically significant. All analyzed specimens from the oc/oc tx oc/oc-EFS-hT group had increased CTX-I release (8–239-fold of oc/oc) and the average CTX-I release was ∼22% that of osteoclasts derived from WT tx WT-EFS-G mice (Fig. 3c), although this difference was not statistically significant. Resorption pits on bone slices were observed for osteoclasts derived from 7/9 transplanted oc/oc mice (Fig. 3e and Table 1), although in lower numbers than the WT mice (Fig. 3f). Overall, the ex vivo analysis thus indicates that a partial rescue of the osteoclast resorptive function was obtained in a majority of mice.

Osteoclasts differentiated from splenocytes of transplanted oc/oc mice express hTCIRG1 and show varying levels of bone resorption ex vivo. Splenocytes were harvested 19–25 weeks after transplantation, seeded on bone slices, and differentiated into osteoclasts for 13 days in the presence of M-CSF and RANKL. Western blot analysis was performed on cell lysates from mature osteoclasts
Data from individual oc/oc mice transplanted with oc/oc cells transduced with EFS-hT
BM, bone marrow; S, sacrificed for osteoclastogenesis, VCN and histology; †, died; n.d., not determined; VCN, vector copy number; WB, western blot analysis.
oc/oc mice transplanted with the EFS-hT-transduced oc/oc cells exhibit a normalized bone phenotype
The bones were analyzed by histopathology 19–25 weeks after transplantation and compared with the osteopetrotic phenotype of oc/oc bones that are solid and lack marrow spaces (Fig. 4b). The oc/oc mice transplanted with EFS-G-transduced WT cells (Fig. 4c) had a normal bone phenotype with normal BM spaces, similar to that of WT mice transplanted with WT-EFS-G cells (Fig. 4a). Of the nine oc/oc mice transplanted with EFS-hT-transduced cells that survived long term, eight mice had a rescued bone phenotype; representative images of the femur of each animal (Fig. 4d–l) show an almost complete recovery of the BM space and normal bone thickness. Only one of the mice that survived until the endpoint (mouse no. 4, Fig. 4g and Supplementary Fig. S2b) exhibited clear signs of osteopetrosis, with a nearly completely solid bone and only a few patches of marrow cells. Histopathology was also performed on tibiae, calvariae, and vertebrae (Supplementary Fig. S3). In all these three bone types, the oc/oc tx oc/oc-EFS-hT mice, except mouse no. 4, had a rescued phenotype comparable with that of the WT animals.
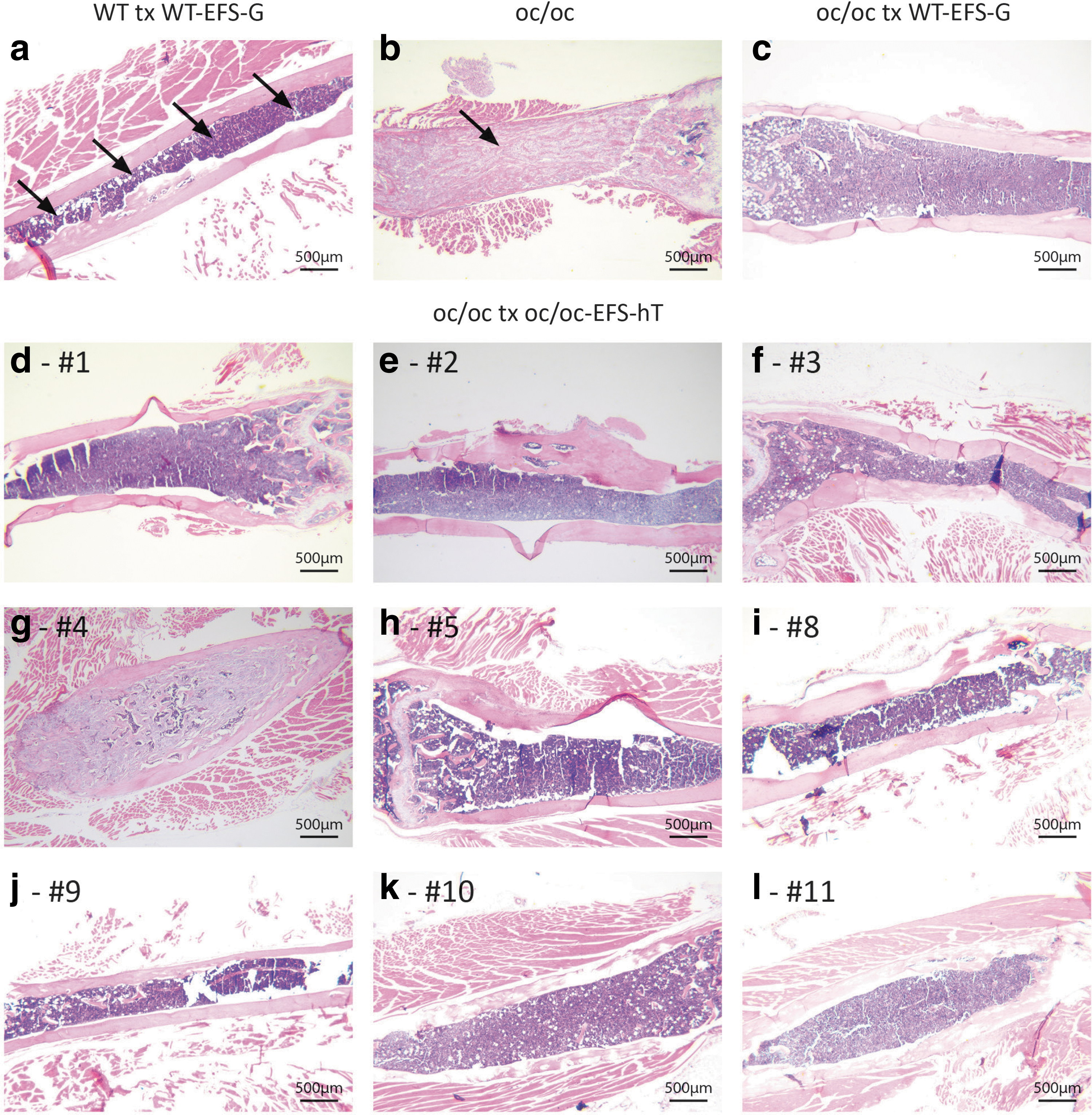
Reversal of the osteopetrotic bone phenotype in oc/oc mice after transplantation of oc/oc cells transduced with EFS-hT as demonstrated by histopathology. Representative images of the femur (2.5 × objective) of a WT tx WT-EFS-G mouse
Isolated BM cells from oc/oc mice transplanted with EFS-hT-transduced cells were also used to assess the BM VCN that was 1.8 ± 0.5. Only 1/9 mice had a low VCN of 0.14 (Table 1, mouse no. 4) and, interestingly, this mouse also had poor histological rescue, as mentioned above (mouse no. 4, Fig. 4g and Supplementary Fig. S2b).
Discussion
The first ex vivo gene therapy based on a gammaretroviral vector for ADA-SCID 23 has been approved and is available in the European Union. Today, more than 200 patients worldwide have received LV-based HSC gene therapy in clinical trials for different diseases and several of these gene therapy products are expected to become registered drugs. 24 The aim of this study together with our previous preclinical work 22 is to prove the safety and efficiency of LV-based HSC gene therapy for IMO.
In this study, we demonstrate that 75% of treated oc/oc mice survived long term after IV neonatal transplantation of c-kit-enriched FL cells transduced with an SIN LV expressing hTCIRG1 under the mammalian EFS promoter in the absence of a marker gene. A total of 9 of 12 transplanted mice survived beyond the oc/oc normal life span of 3 weeks, and at 19–25 weeks of age the mice exhibited signs of osteoclast activity and tooth eruption. With the EFS-mT vector, expressing mouse TCIRG1, only 25% (2/8) of oc/oc mice survived long term and did not show signs of tooth eruption, although the reason for the poor rescue remains unclear. The overall response rate in the present study with EFS-hT is an improvement compared with our previous gene therapy study. 14 The unsuccessful oc/oc-EFS-hT transplantations could be due to low transduction efficiency in the transplanted cells, as in the case of mice nos. 6 and 7 (Table 1) that died after 3 weeks and had lower than average VCN in transplanted cells, or could be due to low engraftment, as may be the case for mouse no. 12, which also died after 3 weeks despite a high VCN of transplanted cells. We tested the EFS-mT vector in parallel with our vector of interest, EFS-hT, as the sequence identity between mouse and human TCIRG1 is only 84%. However, the vector expressing human Tcirg1 provided better rescue in vivo and is clinically relevant, so we focused our continued analysis on this vector.
When osteoclasts were differentiated ex vivo from splenocytes of transplanted mice, the oc/oc tx oc/oc-EFS-hT-derived osteoclasts showed a partial restoration of resorptive function as evaluated by the concentration of the resorption biomarker CTX-I in the media and the presence of resorption pits on bone slices. The trend of increased CTX-I release seen for osteoclasts from all rescued mice was not statistically significant compared with that of oc/oc osteoclasts due to a high variation between cultures. However, CTX-I was substantially increased and was on average 22% that of the CTX-I release of osteoclasts from WT mice and 61-fold of the oc/oc. The different kinetics in TRAP activity between cells derived from oc/oc and oc/oc tx oc/oc-EFS-hT mice could be due to increased osteoclast numbers in nonresorbing cultures, as bone resorption induces osteoclast apoptosis, as part of an osteoclast-rich osteopetrotic phenotype. 25 Despite the apparent kinetic differences in osteoclastogenesis, day 13 was still considered ideal to assess resorption outputs as TRAP activity in WT mice and most transplanted oc/oc mice was similar and peaked at this time point. VCN analysis showed that the transgene is present in BM cells of transplanted mice at the endpoint, and Western blot showed that hTCIRG1 is expressed in mature osteoclasts generated ex vivo from the transplanted animals, although at varying levels. In conclusion, the modest resorption seen ex vivo could either be due to a low fraction of corrected cells or a low level of expression of the transgene in the transduced cells. However, in our previous study, we showed that transplantation of gene therapy-corrected cells in oc/oc mice reversed the disease, even though the in vitro bone resorption capacity of these cells was only 10% of WT cells. 14 In terms of the human disease form, we have shown that the addition of 30% umbilical cord blood CD34+ cells to IMO CD34+ cells in vitro followed by osteoclast differentiation was sufficient to completely restore resorptive function, and levels as low as 2.5% cord blood CD34+ cells mixed with IMO CD34+ cells resulted in significant resorption, likely due to fusion of normal and mutated preosteoclasts to form mature osteoclasts. 26 Thus, gene correction of only a fraction of cells and partial rescue of osteoclast function may be sufficient for clinical benefit when treating patients with IMO.
These conclusions are also supported by the observation that despite the low level of rescue ex vivo, there was an almost complete normalization of the skeletal phenotype in all bones investigated as shown by the histology in the mice that survived long term, which would indicate that only a small proportion of bone-resorbing osteoclasts are needed. Only one transplanted oc/oc mouse still showed signs of osteopetrotic bone with patches of normal BM. This was also the mouse with lowest endpoint VCN (0.14), indicating that BM VCN may correlate with bone phenotype rescue. Similarly, osteoclasts generated from this mouse had relatively low CTX-I release ex vivo and the TRAP activity in the culture medium followed the pattern observed for the oc/oc mice, suggesting that the phenotypic rescue in this mouse was low overall as oc/oc mice are known to have increased osteoclastogenesis. 27 Another sign of reversal of the osteopetrotic phenotype was the presence of tooth eruption. Previously, we had only been able to observe tooth eruption in oc/oc mice by transplantation with WT BM cells 28 and not using genetically modified oc/oc cells. 14 This indicates that the experimental procedure and vector used in the present study have resulted in clinically meaningful improvements in treatment outcome. However, rescued mice remain growth retarded, just as mice transplanted with WT cells. The experience from HSCT in IMO patients, even when performed early, shows that growth retardation can be expected also after gene therapy. 29
In conclusion, we have shown that osteopetrosis in oc/oc mice can be corrected by neonatal transplantation of lentiviral-mediated gene therapy of HSCs. Achieving these results using a clinically applicable vector represents a significant step toward the development of gene therapy for IMO.
Footnotes
Acknowledgments
This work was supported by a research grant to Lund University from Rocket Pharmaceuticals (NY, NY). K.H. is supported by the Danish Research Foundation (Den Danske Forskningsfond). J.R. is supported by a Clinical Research Award from Lund University Hospital and the Foundations of Lund University Hospital. A.S. and M.R. were supported by the European Union (FP7-Health-2010-CELL-PID: no. 261387). H.L. is supported by Marie Curie Initial Training Networks (Euroclast, FP7-People-2013-ITN: no. 607446). The Lund Stem Cell Center is supported by a Center of Excellence grant in life sciences from the Swedish Foundation for Strategic Research. The funders had no role in study design, data collection and analysis, and decision to publish or preparation of the article. Conceptualization: I.M. and J.R.; investigation: H.L., M.R., and I.M.; writing—original draft: H.L., J.R., and I.M.; writing—review and editing: H.L., M.R., A.S., K.H., J.R., and I.M.; funding acquisition: J.R.
Author Disclosure
J.R. has a consultancy agreement with Rocket Pharmaceuticals (New York, NY) for the development of clinical gene therapy of IMO. K.H. holds stocks in Nordic Bioscience. All other authors declare no competing financial interests.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.