
Abstract
Adeno-associated virus (AAV) gene therapy for neurological diseases was revolutionized by the discovery that AAV9 crosses the blood–brain barrier (BBB) after systemic administration. Transformative results have been documented in various inherited diseases, but overall neuronal transduction efficiency is relatively low. The recent development of AAV-PHP.B with ∼60-fold higher efficiency than AAV9 in transducing the adult mouse brain was the major first step toward acquiring the ability to deliver genes to the majority of cells in the central nervous system (CNS). However, little is known about the mechanism utilized by AAV to cross the BBB, and how it may diverge across species. In this study, we show that AAV-PHP.B is ineffective for systemic CNS gene transfer in the inbred strains BALB/cJ, BALB/cByJ, A/J, NOD/ShiLtJ, NZO/HILtJ, C3H/HeJ, and CBA/J mice, but it is highly potent in C57BL/6J, FVB/NJ, DBA/2J, 129S1/SvImJ, and AKR/J mice and also the outbred strain CD-1. We used the power of classical genetics to uncover the molecular mechanisms AAV-PHP.B engages to transduce CNS at high efficiency, and by quantitative trait locus mapping we identify a 6 Mb region in chromosome 15 with an logarithm of the odds (LOD) score ∼20, including single nucleotide polymorphisms in the coding region of 9 different genes. Comparison of the publicly available data on the genome sequence of 16 different mouse strains, combined with RNA-seq data analysis of brain microcapillary endothelia, led us to conclude that the expression level of Ly6a is likely the determining factor for differential efficacy of AAV-PHP.B in transducing the CNS across different mouse strains.
Introduction
Development of effective gene therapies for neurological diseases depends on the availability of gene transfer vehicles capable of widespread high-efficiency transduction of cells in the central nervous system (CNS). Recombinant adeno-associated virus (AAV) vectors have become the platform of choice for development of gene therapies for neurodegenerative diseases as they transduce the CNS at high efficiency and express genes of interest for extended periods of time, 1,2 perhaps indefinitely. Before 2009, AAV gene transfer to CNS was based on intraparenchymal injections with distribution mostly limited to the targeted structure, which in some instances is preferable, but not necessarily for neurodegenerative diseases affecting the entire CNS. The discovery that AAV9 crosses the blood–brain barrier (BBB) after systemic administration in mice 3 started a new era of intravascular gene therapy for CNS diseases. Nonetheless AAV9 vectors delivered systemically to mature animals exhibit a significant limitation, as most transduced cells in the brain are astrocytes and endothelial cells, with very few neurons transduced. 4 –6 The transformative outcomes in children affected with spinal muscular atrophy (SMA) treated by intravascular administration of an AAV9 vector 7 are likely the result of its remarkable tropism for spinal cord motor neurons, which are the primary drivers of the SMA disease phenotype. In contrast, the majority of neurological diseases also affect neuronal populations in the brain and peripheral nervous system. Therefore, the need persists for potent AAV capsids capable of transducing a majority of neurons, astrocytes, oligodendrocytes, and microglia using minimally invasive delivery approaches.
Development of AAV capsids with novel CNS tropic properties has been accomplished using various approaches including in vivo selection of libraries generated by DNA shuffling of existing capsid genes, 8,9 peptide libraries displayed on the capsid surface, 10,11 screening of additional natural variants, 6,12 or even in silico design, 13 but until recently the improvements over AAV9 systemic gene transfer have been incremental.
In vivo selection using Cre recombinase-driven identification of AAV capsids that transduce target cells in the CNS has yielded AAV-PHP.B 14 and AAV-PHP.eB 15 capsids, which are 80- to 300-fold more efficient than AAV9. However, unlike AAV9 that displays consistent properties across species, 4,5,16,17 AAV-PHP.B has been shown to be only approximately twofold more efficient than AAV9 in rats 18 and also offers modest improvement in select neuronal populations in marmoset monkeys, 19 cats, 20 and sheep. 21 These findings in other species suggest that AAV-PHP.B is engaging a nonevolutionarily-conserved molecular mechanism to cross the BBB in mice at high efficiency, although one that remains unknown. Similarly, our knowledge as to how AAV9 crosses the BBB remains largely empirical, with no mechanistic understanding. Uncovering the means by which AAV9, AAV-PHP.B, and AAV-PHP.eB cross the BBB after systemic administration may provide the necessary information to rationally engineer new entities with enhanced potency and perhaps tissue or cell type specificity.
In the course of characterizing the tropism of AAV-PHP.B in our laboratory, we found that its remarkable CNS gene transfer properties in C57BL/6J mice were not reproduced in BALB/cJ mice, a finding also recently reported by others. 22 –25 This unexpected finding provided a unique opportunity to use the power of classical genetics to identify the gene(s) responsible for the differential properties across two inbred mouse strains and ultimately shed light on the molecular mechanisms some AAV capsids use to cross the BBB. We performed whole genome single nucleotide polymorphism analysis in C57BL/6J x BALB/cJ crosses and backcrosses, and we also studied the CNS tropism of AAV-PHP.B in >10 inbred mouse strains with sequenced genomes. This combined approach yielded several candidate genes in a small region of chromosome 15 and represents an important step toward understanding the molecular mechanisms AAV use to cross the BBB and transduce cells in the CNS at high efficiency.
Materials and Methods
AAV vector design and preparation
The single-stranded AAV vector used in this study encodes green fluorescent protein (GFP) with the nuclear localization sequence 3x(PKKKRKV) fused to the 3′ end (nlsGFP) with expression being driven by a chicken beta-actin (CBA) promoter (fused to the cytomegalovirus enhancer and a chimeric intron). The vector is flanked by AAV2 inverted terminal repeats and it also contains the woodchuck hepatitis virus post-transcriptional regulatory element.
AAV-PHP.B helper plasmid was generated from pAR-926 through PCR mutagenesis to match the published VP1 sequence (Genbank KU056473).
AAV vectors were prepared by triple transient transfection of HEK293T cells using polyethylenimine (PEI; linear MW 25,000; Polysciences). Briefly, cells were plated at a density of 2.1 × 107 cells per 15 cm dish. The next day, cells were transfected using PEI with 5.7 μg of the transgene plasmid, 11.4 μg of adenovirus helper plasmid, and 22.8 μg of rep-cap plasmid, per 15 cm dish. Medium was collected at 72 h post-transfection and stored at 4°C, until it was combined with the final collection at 120 h post-transfection. Viral particles from the medium were precipitated with 8% polyethylene glycol 8000 (PEG-8000, VWR) and 500 mM NaCl for a minimum of 2 h at 4°C and resuspended in 150 mM NaCl, 50 mM Tris, pH 8.4. Cell pellets were resuspended in the same buffer, freeze–thawed three times, and combined with the medium fraction. The combined fractions were treated with 50 U/mL Benzonase (Sigma-Aldrich) and then purified by iodixanol gradient centrifugation. 27 Buffer exchange to phosphate buffered saline (PBS) was done using Zeba Spin Desalting columns (7K MWCO; Thermo Fisher Scientific) and further concentration was achieved using Amicon Ultra-15 Centrifugal Filter Unit (100 kDa MWCO; Millipore). Vector titer was determined by quantitative PCR (qPCR) using primers and probes for the bovine growth hormone (BGH) polyadenylation signal (see hereunder for sequences). During the study, several independent batches of vectors were required. The titers of all different batches were normalized to the value of the first preparation used.
Animal procedures
All procedures were performed as approved by the Institutional Animal Care and Use Committee at UMMS and Jackson Laboratory. Animals were maintained at 21 ± 1°C under a 12 h light/dark cycle with irradiated laboratory rodent food and water provided ad libitum.
The inbred strains C57BL/6J (stock no. 00064), C57BL/6NJ (005304), 129S1/SvImJ (002448), FVB/NJ (001800), DBA/2J (000671), AKR/J (000648), BALB/cJ (000651), BALB/cByJ (001026), A/J (0006446), NZO/HILtJ (002105), NOD/ShiLtJ (001289), C3H/HeJ (000659), WSB/EiJ (001145), CAST/EiJ (000928), PWK/PhJ (003715), and CB6F1/J (100007) were all purchased from The Jackson Laboratory. CD-1 mice (strain code: 022) were obtained from Charles River Laboratories. Backcrosses of CB6F1/J × BALB/cJ and BALB/cJ × CB6F1/J were used for genotyping and quantitative trait locus (QTL) analysis.
Systemic delivery
Six to eight weeks old male or female mice were injected intravenously in the lateral tail vein with either 1 × 1012 vg (for histology) or 5 × 1011 vg (for vector biodistribution) in 200 μL and euthanized 1 or 3 weeks postinjection depending on the experiment.
Serum neutralization assays
Intracerebroventricular delivery
Six to eight weeks old male mice were anesthetized using ketamine (75 mg/kg) and dexmedetomidine (0.5 mg/kg), and 5 × 1010 vg (in 5 μL) were injected in the right lateral ventricle using stereotaxic placement (coordinates to bregma: AP−0.5 mm, ML ±1 mm, DV−2 mm) at an infusion rate of 1 μL/min. Animals were recovered using atipamezole (1 mg/kg) and monitored for 7 days postsurgery. No adverse reactions to the surgery were ever observed.
Histology and image processing
Mice were anesthetized with overdose of ketamine (375 mg/kg) and xylazine (37.5 mg/kg) and transcardially perfused with ice-cold PBS. Brain and liver were collected and postfixed in 10% neutral buffered formalin overnight at 4°C and then cryoprotected with 30% sucrose and sectioned by cryostat. Immunofluorescent staining was performed on 10 μm-thick sections on slides (for systemic delivery) or on 40 μm-thick free-floating sections (for intracerebroventricular [icv] delivery). Tissues were permeabilized with 0.5% Triton X-100 in PBS for 10 min at room temperature, blocked with blocking buffer containing 5% fetal bovine serum +5% donkey serum in PBS, and then incubated overnight at 4°C with primary antibody rabbit anti-GFP (1:1,000; Invitrogen, A11122) or rat anti-Ly-6A/E (Sca-1) (1:100; clone D7 eBioscience; Thermo-Fisher Scientific, 14-5981-82). Sections were then washed and incubated with AlexaFluor-555 antirabbit immunoglobulin G (IgG) or AlexaFluor-555 antirat IgG (1:1,000; Invitrogen Molecular Probes, A31572 or A21434) for 1 h at room temperature. Slides were mounted in Vectashield containing DAPI (Vector Labs) and visualized under a Leica DM5500B microscope using the same exposure conditions across the same tissues/groups. All images were processed using Fiji software 28 and changes to brightness or contrast or gamma were kept consistent across groups.
Vector biodistribution
After euthanasia, brain and liver were quickly collected and snap frozen. DNA was isolated from the tissue samples using the QIAGEN DNeasy Blood & Tissue Kit, according to the manufacturer's instructions. Vector genome copy numbers were measured in 100 ng of total DNA by qPCR using the following primers and probes for the BGH polyA in the vector: forward primer CCTCGACTGTGCCTTCTAG; reverse primer TGCGATGCAATTTCCTCAT; probe 56-FAM/TGCCAGCCA/ZEN/TCTGTTGTTTGCC/3IABkFQ. The lower limit of detection in our qPCR assay is 100 gc/100 ng DNA and any sample less than this limit was considered nondetectable.
Experimental results are shown as mean ± standard deviation (SD). Before statistical analysis, the normality of the data was evaluated using the Shapiro–Wilk test. For all data sets considered normally distributed, statistical significance was evaluated by one-way analysis of variance (ANOVA) followed by uncorrected Fisher's Least Significant Difference test. All data sets that did not meet normality requirements were evaluated using the Kruskal–Wallis test followed by uncorrected Dunn's test. Differences were considered significant for p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), or p < 0.0001 (****), as designated. All statistical analyses were performed using GraphPad Prism version 8.0.1 for macOS (GraphPad Software, San Diego, CA).
Genotyping and QTL analysis
Mice (total 115) were genotyped at ∼143,000 markers using the GigaMUGA Mouse Universal Genotyping Array. 29 Genotypes were called using GenCall (7.0.0) in the GenomeStudio Genotyping Module (2.0.2) (Illumina). Genotypes with GenCall quality score GC <0.05 were regarded as missing (genotype “--”), and a table of genotype calls (AA/AB/BB/--) and GC scores for each marker was exported and read into R (3.5.2) for further analysis. For the QTL analysis of the CB6F1/J × BALB/cJ and BALB/cJ × CB6F1/J backcrosses (45 and 50 mice, respectively), only those markers for which two or more distinct genotypes were observed in at least 10 mice were considered, reducing the number of markers to ∼35,000. Here “--” was considered a distinct valid genotype, as missing calls can be informative, for example, for deletions. The estimated AAV counts per 100 ng from PCR were used as a quantitative trait, log10 transformed for better normality and homoscedasticity. Using a standard single-marker regression approach, 30 for each marker a linear model was fit with the trait as response and a categorical factor for genotype as predictor (R function lm). p-values were computed using the R function ANOVA, which is equivalent to a t-test when exactly two distinct genotypes are observed and an F-test otherwise (p-values were also computed while controlling for sex [male vs. female] and pedigree [CB6F1/J × BALB/cJ vs. BALB/cJ × CB6F1/J] of the mice by including these as additive factors in the linear models when testing for the significance of genotype, but this did not make an appreciable difference). Log-likelihoods were computed with the R function logLik.
Filtering of variants
Variants for all tested strains from the region of chr15:70–80 Mb were obtained by querying the Mouse Genome Project database (release REL-1505) from the Wellcome Sanger Institute (
Brain microcapillary endothelia analysis
For isolation of the brain microcapillary endothelium (BME), 8-week old males (C57BL/6J or BALB/cJ) were euthanized by overdose of ketamine/xylazine and the brain was quickly extracted and homogenized in 0.1 M HEPES with a dounce homogenizer. BME was isolated by centrifugation (3,000g, 20 min, 4°C) in 18% Dextran, as previously described. 33 –35
Total RNA from the BME was isolated using TriZol Reagent (Invitrogen) according to the manufacturer instructions and used for either RNA-Seq or qPCR analysis. Complementary DNA (cDNA) synthesis was performed with a mix of random hexamers and oligo(dT) using the High Capacity RNA to cDNA Kit (Applied Biosystems). qPCR was done using Taqman Gene Expression Master Mix (Applied Biosystems) and Taqman probes (Applied Biosystems) for mouse Ly6a (assay no. Mm00726565_s1), mouse Ly6c1 (assay no. Mm03009946_m1), mouse Cldn5 (assay no. Mm00727012_s1), and mouse Hprt (assay no. Mm00446968_m1). The ΔΔCt method was used for calculations of relative expression. 36 Results are reported as mean ± SD.
For protein expression analysis, BME samples were lysed in RIPA buffer containing protease inhibitors. Total protein in the lysates was determined by Bradford assay (Bio-Rad) using serial dilutions of bovine serum albumin as protein standard; 40 μg of total protein was separated in a 4–20% polyacrylamide SDS-PAGE gel (Bio-Rad) and then transferred to 0.2 μm pore nitrocellulose membrane (GE Healthcare, Amersham). Primary antibodies were rabbit monoclonal anti-Ly6A/E (1:1,500; clone EPR3355; Abcam, ab109211) and mouse monoclonal anti-GAPDH (1:500; Abcam, ab8245) for normalization. Detection was performed by chemiluminescence using Clarity Western ECL Substrate (Bio-Rad) and images were acquired using a ChemiDoc system (Bio-Rad).
The normality of the data was evaluated using the Shapiro–Wilk test and statistical significance was evaluated by unpaired t-test. Differences were considered significant for p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), or p < 0.0001 (****), as designated. All statistical analyses were performed using GraphPad Prism version 8.0.1 for macOS (GraphPad Software).
RNA-Seq analysis
Total RNA was extracted from BME samples as previously described, and it was sequenced on an Illumina HiSeq2000 instrument at the Yale Center for Genome Analysis. FASTQ files were mapped to the GRCm38 genome assembly using the STAR aligner (2.5.3a) in two-pass mode. 37 Read counts for each gene (based on Gencode M19 annotations) were computed using htseq-count (0.11.0) in strand-specific mode. 38 Differential expression analysis was performed using quasi-likelihood tests (functions glmQLFit and qlmQLFit) in the R (3.6.0) package edgeR (3.26.4). 39 False discovery rate (FDR) was computed from raw p-values (function topTags), and a cutoff of FDR <0.05 was used to control for multiple hypothesis testing.
In vitro assays
The Ly6a alleles (exons, introns, polyA, but no promoter) from C57BL/6J and BALB/cJ mice were cloned into the CSCW2 lentivirus vector plasmid under the CMV promoter. 40 Cloning of Ly6a alleles was done through initial PCR amplification of genomic sequence from either mouse strain, which was then combined with gene fragments synthesis (Genewiz) using Gibson assembly (NEBuilder, New England Biolabs). All sequences were validated by Sanger sequencing.
HEK293T cells were plated in 12-well multiwell plates and transfected using Lipofectamine 2000, according to manufacturer's instructions. At 72 h post-transfection, cells were harvested either for RNA or protein isolation. Samples for gene expression analysis were processed as described. For protein expression analysis, cells were washed with ice-cold PBS, lysed in RIPA buffer containing protease inhibitors, and processed as already described. Primary antibodies were rabbit monoclonal anti-Ly6A/E (1:1,000; Abcam, ab109211) and mouse monoclonal anti-α-tubulin (1:10,000; Sigma-Aldrich, T6199.) for normalization. Densitometry analysis of Western blot images was performed using ImageJ software (NIH).
In a parallel experiment, cells transfected with Ly6a expression plasmids were infected with AAV-PHP.B-CBA-nlsGFP vector (3,000 gc/cell) or AAV9-CBA-nlsGFP (10,000 gc/cell) at 48 h post-transfection. Twenty-four hours after infection, five fields of view per well were imaged for GFP expression in each experiment. Signal intensity was measured in all images using Fiji software. 28
Experimental results are the average of at least three independent experiments and are presented as mean ± SD. Before statistical analysis, the normality of the data was evaluated using the Shapiro–Wilk test and statistical significance was evaluated by one-way ANOVA followed by Sidak's multiple comparisons test. Differences were considered significant for p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), or p < 0.0001 (****), as designated. All statistical analyses were performed using GraphPad Prism version 8.0.1 for macOS (GraphPad Software).
Results
Three weeks after systemic administration of 1 × 1012 vg AAV-PHP.B-CBA-GFPnls vector in 6–8-week old C57BL/6J mice, GFP expression was widespread throughout the brain (Fig. 1A). In accordance with the original report, 14 the CNS gene transfer efficiency, as measured by vector genome content, was 60-fold higher than AAV9-CBA-nlsGFP (Fig. 1C). Surprisingly, in age-matched BALB/cJ mice treated identically, there was no evidence of GFP expression in the brain (Fig. 1A), and vector genome content was below the detection limit (Fig. 1C). AAV9-CBA-nlsGFP transduced cells in the brain as expected in both C57BL/6J and BALB/cJ mice, although at lower levels in the latter strain (Fig. 1A). This trend was also reflected in vector genome content (Fig. 1C). There were no significant differences in liver transduction between mouse strains treated with the same vector (Fig. 1B, D), with reduced liver transduction by AAV-PHP.B as previously reported. 14 These findings presented an opportunity to elucidate the mechanism used by AAV-PHP.B for efficient CNS gene transfer by systemic administration.
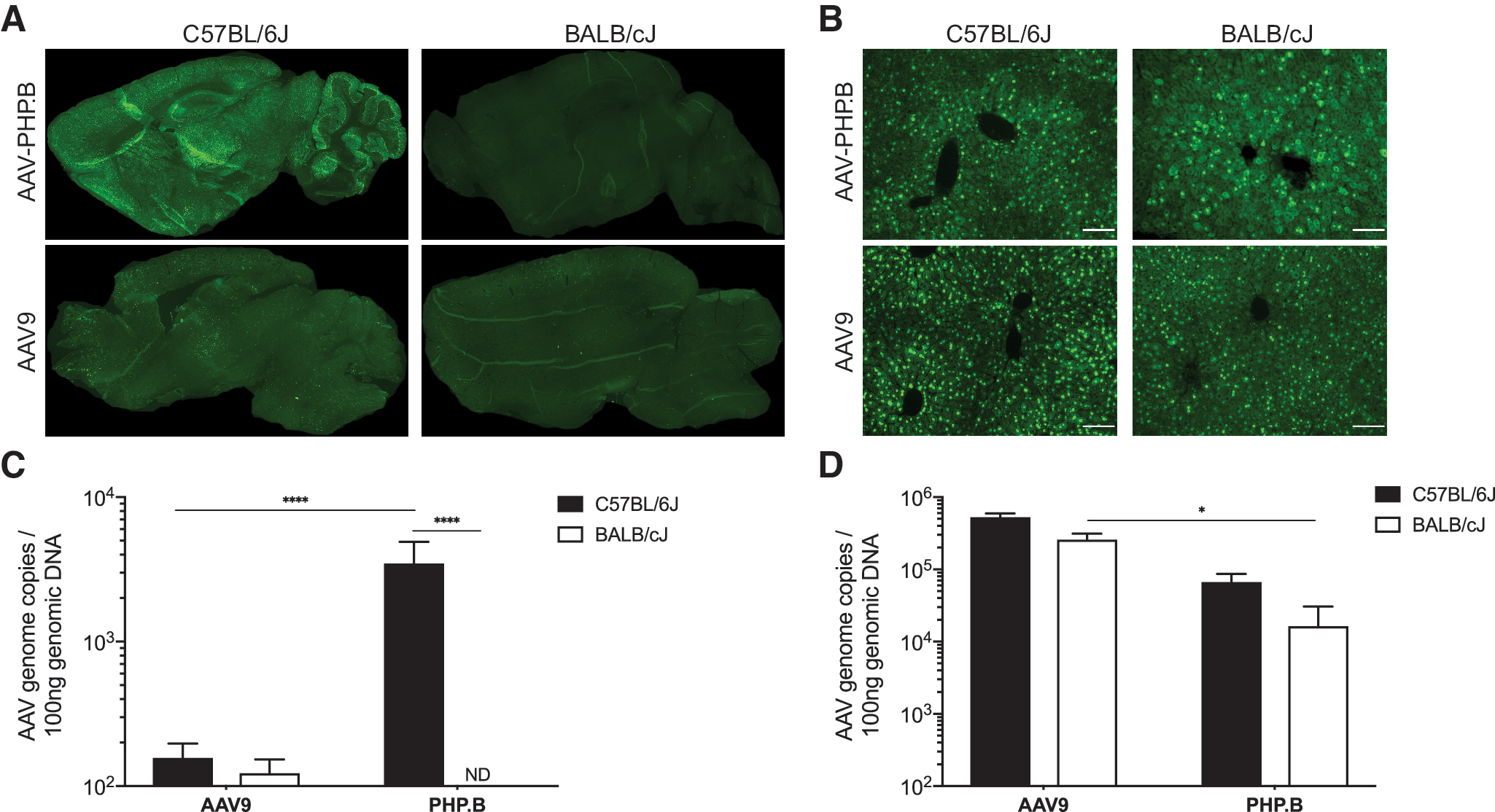
Brain and liver transduction in C57BL/6J and BALB/cJ mice after systemic delivery of AAV-PHP.B and AAV9.
AAV can interact with serum proteins, which can improve or reduce their efficacy. 41,42 To evaluate the role of serum proteins on the reduced CNS transduction in BALB/cJ mice, either through neutralization in BALB/cJ or enhancement in C57BL/6J mice, we tested the effects of preincubation and serum transfusion across mouse strains. The CNS transduction properties of AAV-PHP.B-GFP vector remained unchanged by serum (Fig. 2).
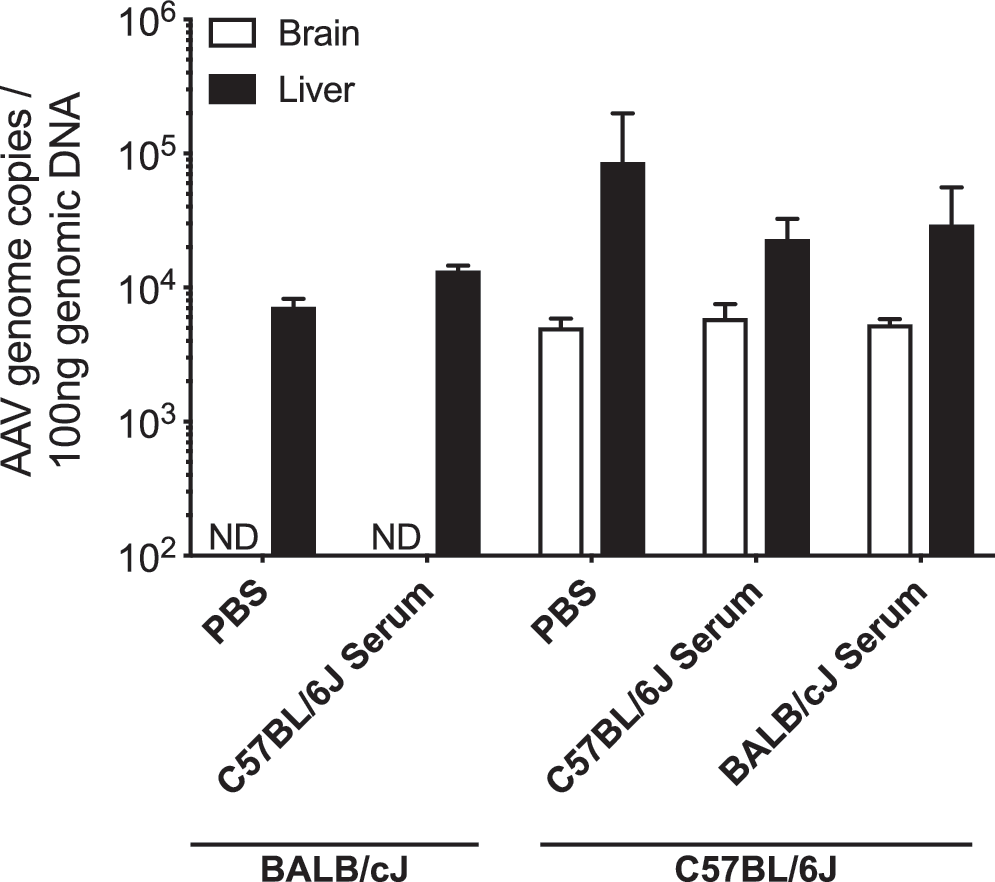
Serum neutralization assays in C57BL/6J and BALB/cJ mice. Quantification of AAV vector genome copies per 100 ng of total genomic DNA in brain and liver of C57BL/6J and BALB/cJ mice. Animals received 5 × 1011 vg
Systemic delivery of AAV-PHP.B-GFP was also tested in inbred FVB/NJ and DBA/2J mice as well as in outbred CD-1 mice. The vector genome content in the brain was comparable with C57BL/6J in both inbred strains but lower in outbred mice (Fig. 3A, B). To determine the inheritance pattern of the AAV-PHP.B phenotype in BALB/cJ mice, we crossed C57BL/6J and BALB/cJ mice and tested transduction of AAV-PHP.B in the F1 generation. Our data showed a 60% reduction in AAV vector genome content in the brain of CB6F1/J mice (Fig. 3C), whereas it was comparable with C57BL/6J in liver (Fig. 3D), suggesting a recessive genetic trait as the etiology of the differential transduction pattern.
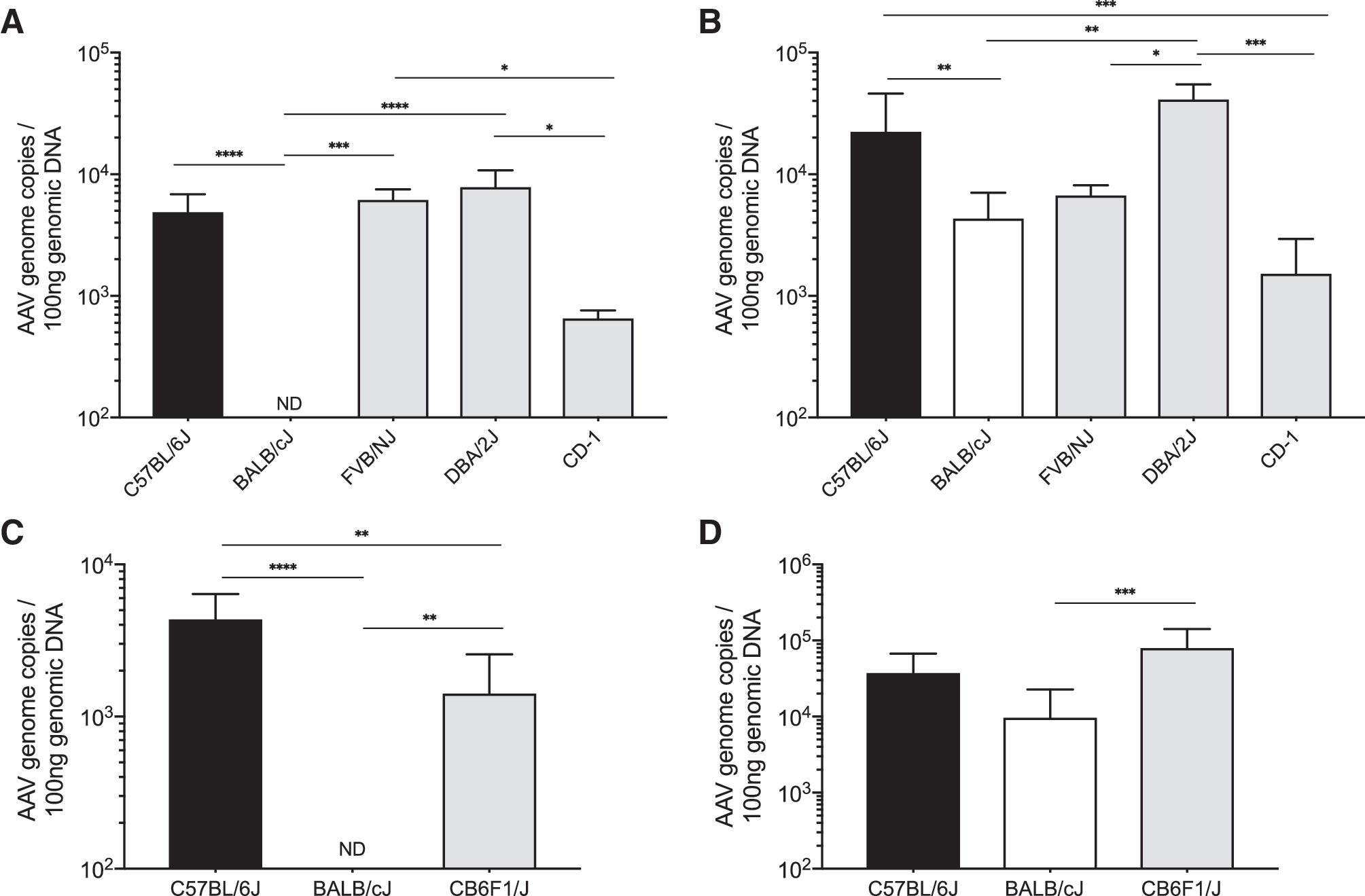
Brain and liver transduction by AAV-PHP.B in multiple mouse strains.
To determine the gene(s) involved, CB6F1/J mice were backcrossed to BALB/cJ and the phenotype of AAV-PHP.B transduction was assessed in the progeny and each mouse was genotyped for >143,000 SNP markers across the mouse genome. QTL analysis identified the most significant markers to be located in an interval on chromosome 15 from 72.2 to 78.6 Mb in mm9/MGSCv37 coordinates (Fig. 4B, C). This interval contains 79 markers with p < 10−38; it corresponds to the interval 72.4–78.8 Mb in mm10/GRCm38 coordinates, which encompasses ∼250 genes, ∼150 of which are protein-coding genes (Fig. 4D).
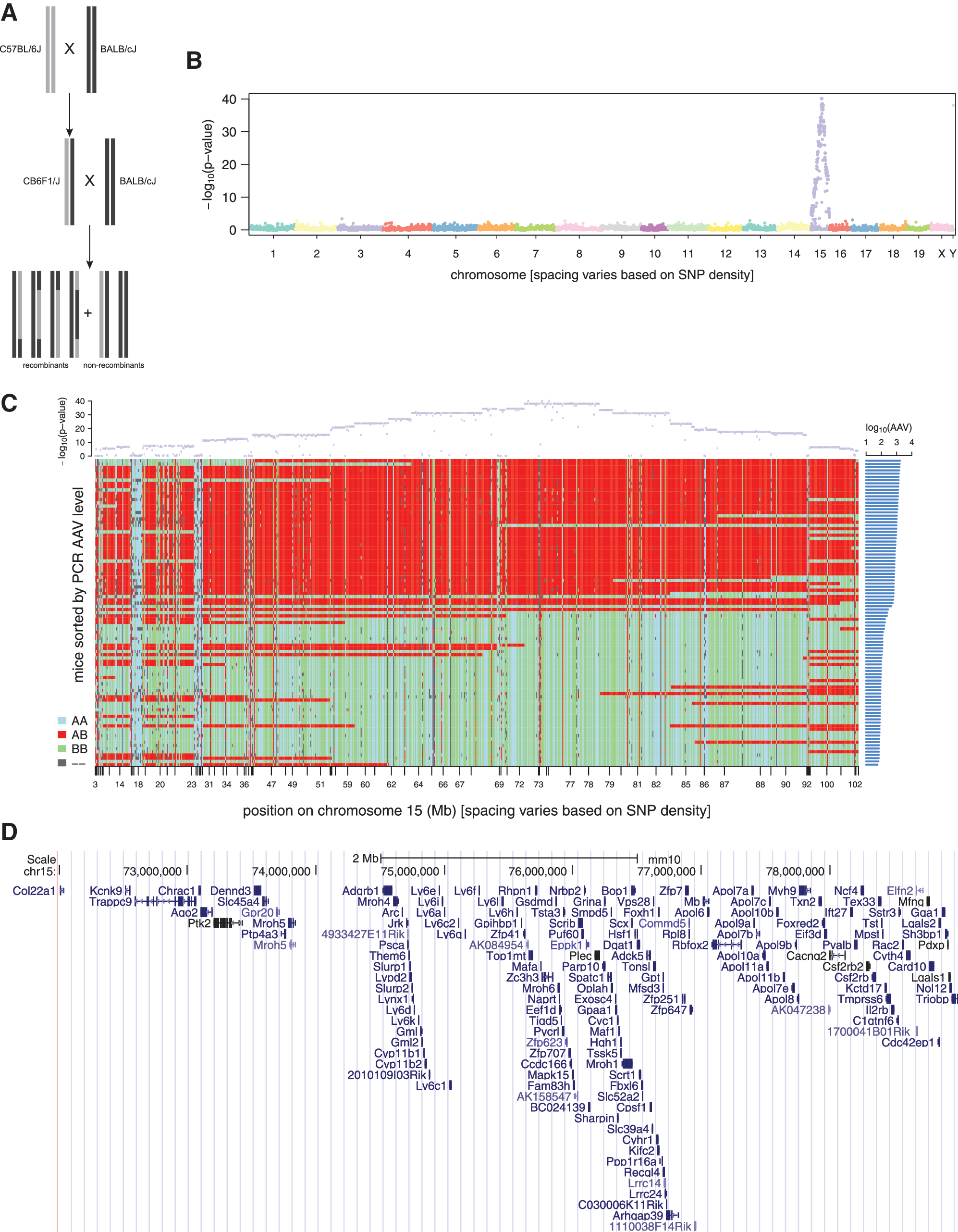
QTL analysis.
To identify candidate causal variants in this region, we considered all variants in this region that were present in public whole genome sequencing (WGS) data, as not all sites are assessed by the GigaMUGA chip. Variants were filtered and prioritized based on their predicted functional consequences, and on whether genotypes at these positions were consistent with AAV levels for other mouse strains as well as the CB6F1/J and BALB/cJ strains (Fig. 5A).
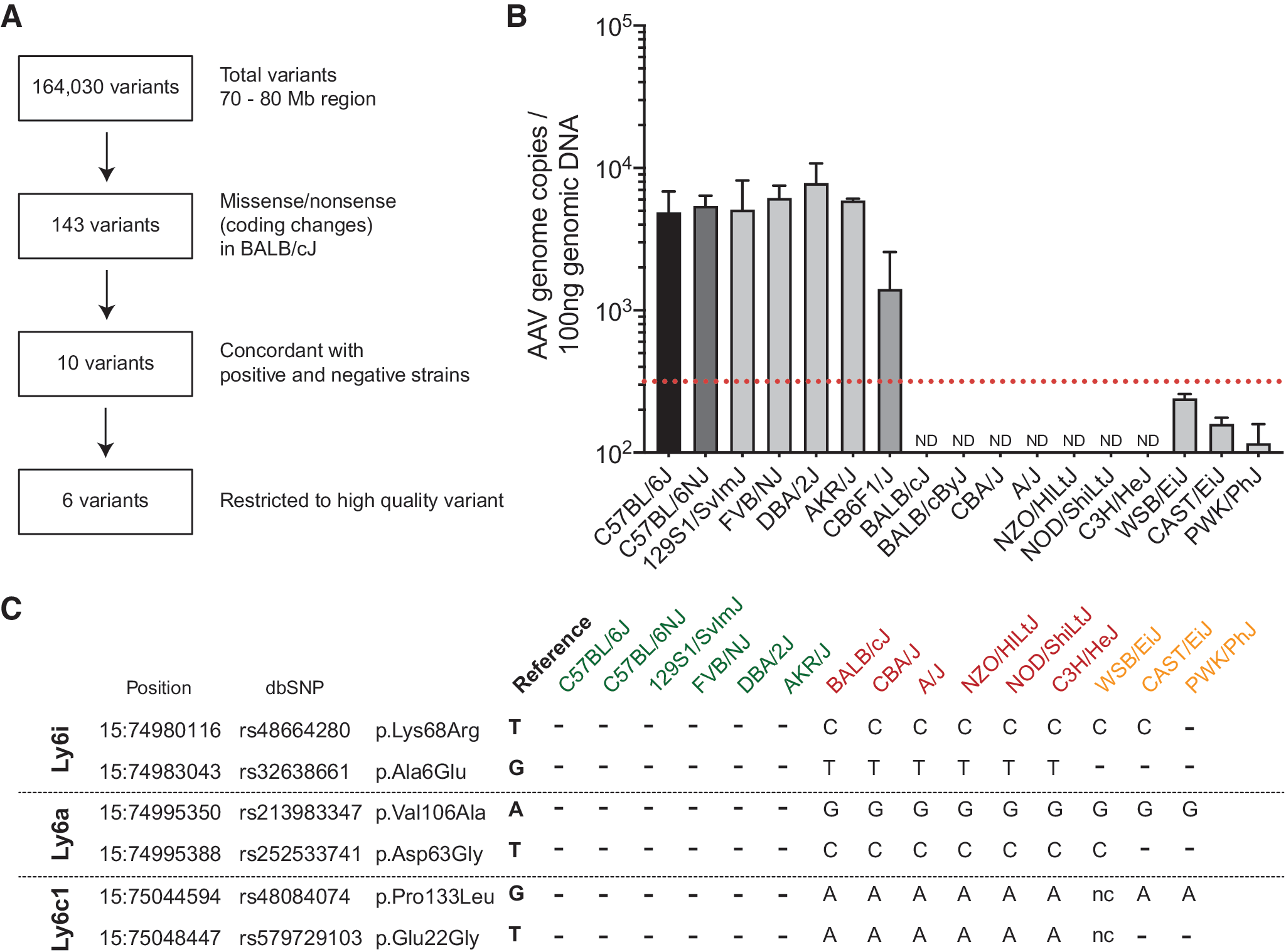
Three candidate genes emerge from analysis of AAV-PHP.B transduction profile in CNS of additional inbred mouse strains.
We analyzed transduction efficiency of AAV-PHP.B in 16 inbred mouse strains (Fig. 5B). Some strains had brain AAV genome copy levels intermediate between CB6F1/J and BALB/cJ (e.g., WSB/EiJ), whereas others resembled BALB/cJ. Although AAV levels could be determined by multigenic factors or multiallelic loci, we began with a binary classification of strains as having high or low AAV copy number. Based on AAV levels observed in the backcrossed mice in the QTL analysis, we determined the cutoff between high and low levels (or “positive” and “negative” strains) to correspond to normalized AAV counts of 102.5 = 316 per 100 ng genomic DNA. We then compared the genome sequences of all the mouse strains (except CD-1 strain) using the Mouse Genome Project database. A total of 164,030 variants were retrieved for the 70–80 Mb region of chromosome 15. Next these were filtered to coding change variants, which restricted the number of variants to 143. These variants were then restricted to those that were concordant in all positive strains (C57BL/6J, 129S1/SvImJ, FVB/NJ, DBA/2J, and AKR/J) and also concordant in the main negative strains (BALB/cJ, CBA/J, A/J, NZO/HlLtJ, NOD/ShiLtJ, and C3H/HeJ). Restricting analysis to high-quality variants identified six amino acid changes in three Ly6 genes: Ly6i,
In brain, Ly6i, Ly6a, and Ly6c1 are expressed mainly in the endothelial cells, 43 and we, therefore, performed transcriptomic profiling on brain microcapillary endothelia from C57BL/6J and BALB/cJ mice. This allowed us to test for expression differences in these particular genes while also getting a global view of differences in transcript levels between the strains. The analysis of the RNA-Seq data indicated differential expression in 701 genes at a cutoff of FDR <0.05, which here corresponded to raw p-value <0.00064 (Fig. 6A, B; Supplementary Table S2). Of these 701 genes, 14 were located on chromosome 15 and 2 were in the interval 70–80 Mb: Ly6a (p = 2.3e-13), which had 7.5-fold higher expression in C57BL/6J than in BALB/cJ samples, and Tonsl (p = 1.9e-04), which had 2.1-fold lower expression in C57BL/6J than in BALB/cJ. Neither Ly6i nor Ly6c1 showed a significant difference (p = 0.28 and p = 0.46, respectively). Ly6i had low expression levels in general (0–4 reads per sample), whereas Ly6c1 had higher levels (14–197 reads per sample) but levels differed by <1.15-fold between strains (Fig. 6B). Expression levels for Ly6c1 and the decreased levels of Ly6a in BALB/cJ mice were confirmed by qPCR (Fig. 6C, D); however, Ly6i messenger RNA levels are below the detection limit of our qPCR assay. Furthermore, we were unable to detect Ly6a protein in the midbrain (Fig. 6E) or in the microcapillary endothelia (Fig. 6F) of BALB/cJ mice.
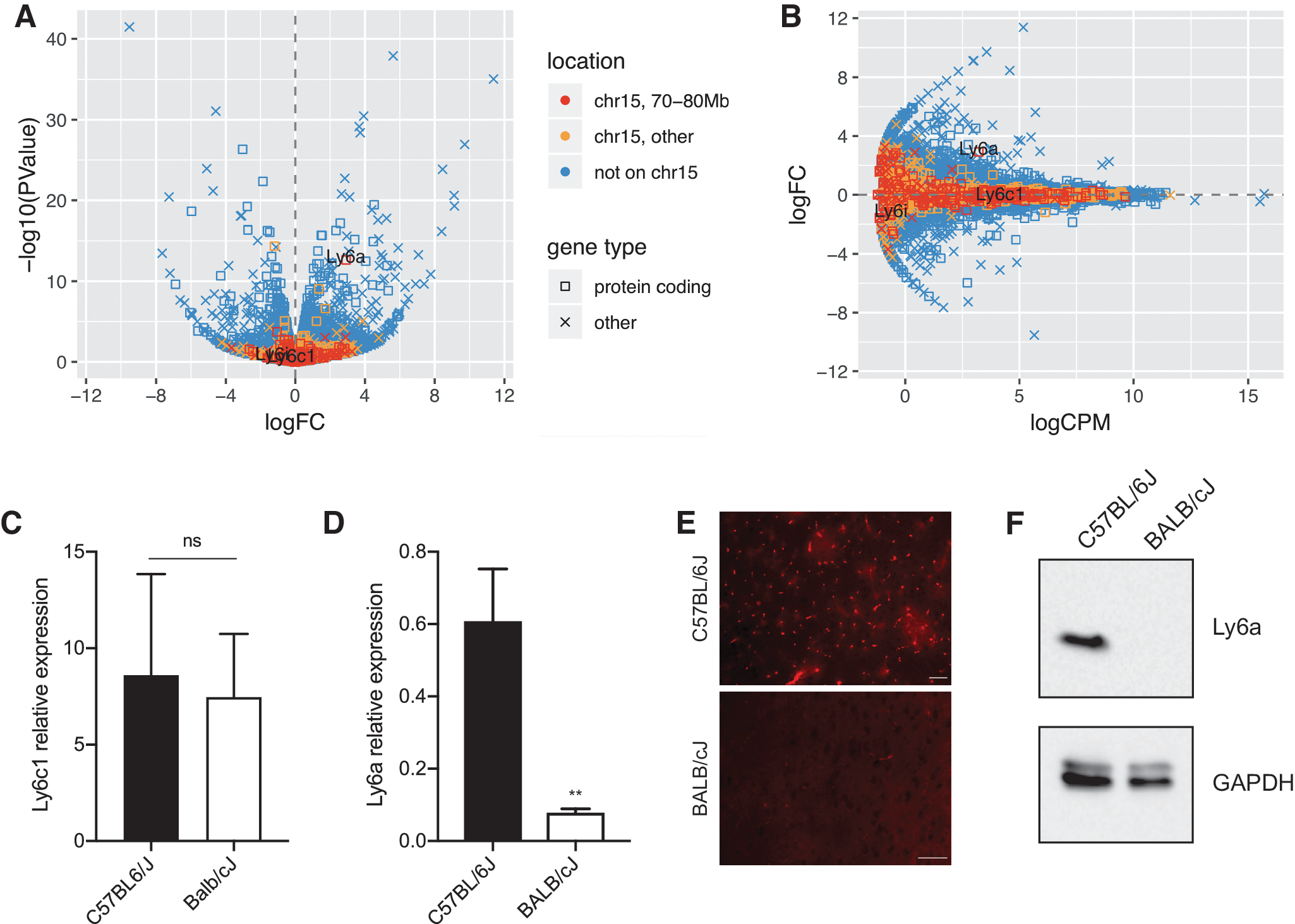
Ly6a expression levels in brain microcapillary endothelium of C57BL/6J and BALB/cJ mice.
To determine whether the CNS transduction profile of AAV-PHP.B across mouse strains was caused either by amino acid changes in the coding sequence of Ly6a or by differential expression levels, HEK293T cells were transiently transfected with expression plasmids carrying either the BALB/cJ or C57BL/6J Ly6a allele. No significant differences in Ly6a expression were detected, either at the RNA (Fig. 7A) or protein level (Fig. 7B, C). Interestingly, expression of C57BL/6J and BALB/cJ Ly6a alleles increased by five-fold the transduction efficiency of AAV-PHP.B in HEK293T cells when compared with mock-transfected controls (Fig. 7D–G), whereas no significant differences were observed in the transduction efficiency of AAV9 for the same cell line (Fig. 7H–K). These data suggest that Ly6a expression levels are more important than amino acid differences between alleles.

Ly6a alleles enhance AAV-PHP.B transduction of HEK293T cells.
Finally, we analyzed whether the CNS transduction profile of AAV-PHP.B infused into CSF also varied between C57BL/6J and BALB/cJ mice. icv delivery of 5 × 1010 vg of AAV-PHP.B-CBA-nlsGFP in C57BL/6J mice resulted in widespread and uniform distribution throughout the entire brain (Fig. 8A). In contrast, in BALB/cJ mice, brain GFP expression was restricted to the injection site (Fig. 8B), similar to what has been observed for AAV9. 44 Further studies are necessary to determine whether this is a function of CSF flow or vascular adsorption of AAV-PHP.B and subsequent transduction, but it suggests that BBB crossing is not the only mechanism differentially regulated across mouse strains.

GFP expression in brain after icv delivery of AAV-PHP.B. Distribution of GFP expression in cerebrum (top panel) and cerebellum (bottom panel) of
Discussion
The recent development of AAV-PHP.B has shown that AAV capsids can be engineered to more efficiently cross the BBB. 14 Although BBB architecture is similar across species, the mechanisms of transport are still not fully understood. In this study we used classical mouse genetics to determine the potential mechanism by which AAV-PHP.B transduces the brain with exceptional efficiency after systemic administration.
In a fortuitous experiment, our group discovered the exceptional systemic CNS gene transfer properties of AAV-PHP.B documented in C57BL/6J mice were not replicated in BALB/cJ mice and several other strains, a finding also reported by others recently. 20 –25 Using a combination of QTL analysis of SNPs in C57BL/6J, BALB/cJ, and F1 backcrosses as well as in silico analysis of coding region SNPs in the candidate region across mouse strains, we identified Ly6a as the gene responsible for the differential CNS transduction efficiency across mouse strains.
During preparation of our article, two other reports were published showing that Ly6a is the gene responsible for the exceptional CNS transduction properties of AAV-PHP.B. 24,25 One of the reports concluded that the V106A change causes the differential transduction efficiency across mouse strains. 25 However, we show here that overexpression of either Ly6a protein isoform greatly enhanced the transduction efficiency of AAV-PHP.B, suggesting that amino acid differences between alleles are not the major determinant of the differential transduction profile. Furthermore, expression analysis of Ly6a in brain microcapillary endothelial cells indicates that lower Ly6a expression in BALB/cJ mice is likely why AAV-PHP.B is unable to transduce their CNS after systemic administration. These data are in accordance with the first study to be published on this topic by Hordeaux et al., 24 wherein it is shown that AAV-PHP.B CNS gene transfer is absent in Ly6a knockout mice. The mechanism responsible for lower Ly6a expression levels between alleles is unknown, but there are numerous SNPs in noncoding regions, including the region upstream of the start codon, which could be responsible for epigenetic silencing of the BALB/cJ allele. Further studies will be necessary to further investigate this possibility.
Ly6a (lymphocyte activation protein-6A), also known as Sca-1 (stem cell antigen-1), is a glycosylphosphatidylinositol-anchored cell-surface glycoprotein, which is part of the lymphocyte antigen-6 (Ly6)/urokinase-type plasminogen activator receptor family. 45 Ly6a is a common marker of murine hematopoietic stem cells (HSCs) and it also plays an important role in regulating T cell proliferation in mice. 45,46 Ly6a is expressed at variable levels throughout different organs in adult mice (Mouse ENCODE transcriptome project 47 ), with kidney showing the highest expression levels (234.89 Reads Per Kilobase of transcript, per Million mapped reads [RPKM]), whereas liver shows approximately half the expression level of cortex (6.29 vs. 15.18 RPKM). Moreover, transcriptome analysis of the cerebral cortex reveals that Ly6a is expressed almost exclusively in endothelial cells. 43 Interestingly, this gene has two haplotypes, Ly6a and Ly6b, that represent two allelic variants Ly6E.1 and Ly6A.2, respectively, each one of them found in distinct groups of inbred mouse strains. 48,49 Furthermore, Ly6A.2 expression (C57BL/6J-like) is higher and more widespread, whereas strains that carry the haplotype Ly6a (BALB/cJ-like) are resistant to infection by adenovirus, 50 suggesting an evolutionary etiology for divergence.
Despite the important role it plays in HSCs, Ly6a has no known ligands, but may coregulate lipid raft signaling. 45 Since Ly6a can affect the signaling and structure of lipid rafts, it is possible that lack of Ly6a in BALB/cJ mice affects endocytosis and intracellular trafficking. This hypothesis could also explain why after CSF delivery, AAV-PHP.B displays a vastly different transduction profile in BALB/cJ and C57BL/6J. In addition, it is possible that expression of Ly6a in neurons and glia, although at lower levels than in endothelial cells, may be a major factor in the transduction profile of AAV-PHP.B in C57BL/6J mice as suggested by our CSF delivery data. Efficient CNS gene transfer by systemic administration may be achieved by future AAV capsids that target receptors that are expressed in endothelial cells, but also in target cells in the brain. The unique widespread CNS transduction profile of AAV-PHP.B after icv delivery in adult mice raises interesting questions about the mechanisms that govern AAV distribution/transduction after CSF delivery. Most AAV capsids shown to transduce neurons and glia at high efficiency after direct intraparenchymal injection (e.g., AAV9, AAVrh10, and AAVrh8) are very inefficient when delivered icv in adult mice. 44,51 The mechanism by which Ly6a mediates the broad transduction of AAV-PHP.B in C57BL/6J mice remains to be elucidated in future studies using conditional Ly6a knockout mice that will allow selective deletion of the gene in specific cell populations in the CNS.
The existence of homologs in other species is also unknown, with exception of the rat, where an ortholog with 63% homology exists. This could explain why AAV-PHP.B exceeds AAV9 transduction in rats by twofold. 18 Conversely, species with no known orthologs such as marmosets, NHPs, cats, or sheep show no increased benefit of AAV-PHPB over AAV9. 19 –21
In humans there is no direct homolog to Ly6a, however, several genes from the LY6 family are located on chromosome 8q24. 46,52 These include LY6E, a gene that is upregulated in AIDS and has been correlated with severe HIV-1 infection by promoting entry into host cells. 53 Further experiments are needed to determine whether a human homolog can be targeted to increase AAV transduction of the CNS in patients.
In conclusion, this study identified a potential new target for AAV engineering to dramatically improve CNS transduction upon systemic or CSF delivery. Nonetheless, further experiments are needed to elucidate the exact function of Ly6a in CNS transduction and whether a human homolog can be utilized to increase CNS gene transfer in patients.
Footnotes
Author Disclosure
The authors have no conflicts of interest to declare.
Funding Information
This study was supported by the CHDI Foundation.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.