
Abstract
Recombinant adeno-associated virus (rAAV) vectors have become one of the most promising and efficacious delivery vehicles for human gene therapy; however, low infectivity remains a major ongoing obstacle in the clinical application of rAAV vectors. Multiple strategies, including rAAV capsid modification and the application of pharmacological reagents, have been explored to enhance rAAV vector gene delivery. Recently, a new strategy using native proteins or various peptides has shown promise for increasing rAAV transduction locally or globally. This review summarizes the current status of protein- and peptide-based strategies and mechanisms to modulate rAAV transduction. We also provide a potential insight regarding the design of effective approaches for rAAV transduction enhancement in future clinical studies.
Introduction
In recent decades, nonpathogenic recombinant adeno-associated virus (rAAV) vector-based gene therapy has expanded substantially and achieved sustained phenotypic corrections in clinical trials. 1,2 Among the current gene therapy mechanisms, the rAAV vector has multiple advantages: it confers a poor immune response, exhibits a well-tolerated safety profile, and promotes long-term persistence of transgene expression. 3 –6 Its capsid, which is composed of three viral proteins (VP1, VP2, and VP3), is highly variable among the 13 known AAV serotypes and numerous natural variants, 7 which leads to remarkable variability in tissue tropism and a wide application for targeted therapies. 8
The therapeutic efficacy and safety of rAAV vectors have been broadly verified in clinical trials. Notably, two rAAV gene therapy products for inherited vision loss and spinal muscular atrophy separately have been approved by the U.S. Food and Drug Administration (FDA). 2,9 However, a few challenges, such as low infectivity in specific tissues, pre-existing neutralizing antibodies (Nabs), nonspecific tropism, capsid-specific cytotoxic T lymphocyte (CTL) response, and potential toxicity, remain for the clinical application of the rAAV vector. 4,10 –13 Furthermore, relatively low AAV infectivity leads to the need for very high viral vector doses. Different strategies, such as AAV capsid engineering and the utilization of a pharmacological agent, have been explored extensively to increase rAAV transduction. The current methods for rAAV capsid engineering include rational design, directed evolution, and computationally designed capsids. 1 Rationally designed rAAV vectors have been successfully developed by many researchers to enhance transduction. For example, the galactose receptor-binding site of AAV9 was engrafted into the AAV2 capsid to improve capsid transduction. 14 In another case, the mutated surface-exposed tyrosine residues of the AAV capsids dramatically increased the mouse liver and retinal transduction of AAV2 as well as the muscle transduction of AAV6. 15 –17 Notably, the chimeric AAV capsid AAV2.5 was generated from the AAV2 capsid with five mutations from AAV1 and has been used in clinical trials for Duchenne muscular dystrophy therapy because of its increased muscle tropism. 3 Among the directed evolution techniques, AAV-shuffled library-based screening is a popular approach to develop AAV mutants for enhancing AAV transduction in vivo. For example, a novel central nervous system tropic rAAV-B1 vector was isolated via a single round of in vivo selection from a DNA-shuffled AAV library, resulting in widespread neuronal subpopulation transduction in the mouse brain after systemic administration. 18 We isolated novel AAV chimeric capsid mutants with the ability to escape the Nabs from patient serum using directed evolution techniques in vitro and in mouse muscle. From these mutants, we identified a motif in variable region III of the AAV6 capsid that confers robust muscle tropism. 19 In addition, AAV variants that were screened and selected from an AAV-shuffled library in a xenograft liver model achieved high transduction in human hepatocytes. 20,21 These studies provide an effective strategy for developing personalized AAV capsids that exhibit high transduction efficacies; however, AAV capsid engineering strategies may potentially change the original structure and tropism of AAV. 22,23 Most importantly, no reliable model to test these AAV mutants and predict their transduction efficiency in humans currently exists. Respecting these concerns, other approaches have been exploited to effectively increase rAAV vector transduction without engineering the capsid. Some pharmacological agents have been found to boost AAV transduction by interfering with the life cycle of rAAV vectors, especially with their intracellular trafficking and synthesis of the genomic second strand. 24 –27 The proteasome inhibitors bortezomib, MG132, and carfilzomib can increase the transduction of multiple AAV serotypes both in vitro and in vivo through proteasome inhibition. 26 Notably, MG132 has been reported to enhance canine factor VIII (FVIII) activity and reduce blood loss in hemophilia A dogs with the coadministration of MG132 and AAV8 encoding canine FVIII. 27 Several other FDA-approved chemicals, such as teniposide and arsenic trioxide, have also been reported to enhance AAV transduction. 28 Moreover, we recently demonstrated that polyploid rAAV vectors, generated from the cotransfection of AAV helper plasmids of different serotypes or mutants, are able to enhance AAV transduction in the liver and muscle and escape the pre-existing Nabs. 28,29 Since polyploid vectors contain subunits from different AAV capsids, they potentially acquire advantages from parental serotypes or mutants for enhanced AAV transduction. Additional approaches without capsid modification involve proteins and peptides, which have attracted considerable attention for a wide range of applications in the fields of gene therapy and biotechnology. 30,31 Herein, this review primarily highlights the recent status of protein- and peptide-based strategies to enhance AAV transduction and provides a valuable insight regarding the design of effective strategies for clinical trials.
Effect of Serum Proteins on AAV Transduction
Without capsid engineering, some naturally existing proteins can affect the efficacy of AAV transduction. These natural serum or nonserum proteins, which may interact with rAAV vectors, have been identified using mass spectrometry and two-dimensional gel analysis. 32 –35 Some of these proteins increased AAV transduction, while several others showed negative effects. In addition, enhanced AAV transduction endowed by binding antibodies without detectable neutralizing activity was also observed. 35 All the proteins that have been reported to affect AAV transduction in this review are listed in Table 1.
List of proteins that regulate adeno-associated virus transduction
CRP, C-reactive protein; G3BP, galectin 3 binding proteins; HAS, human serum albumin; LDL, low-density lipoprotein; PF4, platelet factor 4; vWF, von Willebrand factor.
Serum protein enhancement of AAV transduction
Previous studies have found that some serum proteins directly interact with AAV virions to enhance AAV transduction. The incubation of rAAV vectors with human serum increased the transduction of serotype 1–9 AAV vectors in vitro, and enhanced liver and muscle transduction was observed after administering rAAV8 preincubated with human serum via either retro-orbital or muscle injection, respectively. 34 After rAAV8 binding protein pull-down and mass spectrometry analysis, human serum albumin (HSA) was found to directly interact with AAV virions for transduction enhancement. Notably, rAAV8 vectors preincubated with HSA boosted FIX expression and improved correction in hemophilia B mice, which suggested the therapeutic possibility of using a low dose of rAAV vectors for hemophilia and other diseases. 34 Further investigation indicated that HSA, transferrin, and low-density lipoprotein (LDL) could interact with AAV8 and enhance the mouse liver transduction of rAAV8 vectors by increasing the cell binding ability of rAAV8. 36 Interestingly, no further increase was observed when rAAV8 was incubated with a combination of the serum proteins, which suggests that these serum proteins may competitively bind to the same locations on rAAV8 virions. 36 Strikingly, when we performed the Nab analysis with intravenous immunoglobulin (IVIG), we found that a high concentration of IVIG was able to block AAV transduction, while a low concentration of IVIG without neutralizing activities actually increased AAV transduction. 34 The phenomenon suggested that non-neutralizing IgG could increase AAV transduction. Indeed, another study confirmed that binding antibodies induced by rAAV8 without any neutralizing function enhanced the uptake of rAAV8 vectors and increased AAV transduction. 35
Among all of the serotypes, AAV9 is superior for viral distribution and is able to efficiently transduce brain, muscle, and other tissues via peripheral vein injection 37 ; however, HSA increases the transduction of rAAV9 in the liver but not in the other tissues. 32 Thus, it is possible that HSA does not have the ability to impact vascular permeability. In the following studies, we observed that human serum increased the vascular permeability of rAAV9 vectors in vitro. Some serum proteins, such as fibrinogen, fibronectin, alpha-1-acid glycoprotein, and plasminogen, that are involved in vascular permeability have been identified to have the potential to increase AAV9 global transduction. After systemic injection in mice, it was observed that these serum proteins enhanced the global transduction of rAAV9 vectors in mice. 32 Interestingly, platelet factor 4 (PF4) and von Willebrand factor (vWF) increased rAAV9 transduction in most of the major organs, but not in the liver.
Some human serum proteins have been used in the clinic. Our extended study found that clinical-grade HSA could increase liver transduction after systemic administration of the preincubated HSA and rAAV8 complex. 34 Cryoprecipitate is mainly composed of fibrinogen, fibronectin, and vWF and has been used to treat patients with hypodysfibrinogenemia, von Willebrand disease, hemophilia A, and bleeding related to thrombolytic therapy. 38 As mentioned above, we have also shown that cryoprecipitate can interact with rAAV9 and enhance AAV global transduction after systemic injection of the rAAV9 and cryoprecipitate complex. 32 These findings strongly suggest that albumin and cryoprecipitate could be used for enhancing the transgene delivery of rAAV8 or 9 vectors in future clinical trials.
It has been reported that mouse C-reactive protein (CRP) is able to increase the transduction of AAV1 and AAV6. In CRP-deficient mice, the transduction efficiency of AAV1 and AAV6 was lower than that in wild-type C57BL/6 mice. When the preincubated AAV6/CRP complex was injected into the CRP-deficient mice, AAV6 transduction was restored to a level similar to that in wild-type mice. 39 It is interesting to note that the human CRP fails to bind to either AAV1 or AAV6 for transduction enhancement. In addition, CRP from both mice and humans does not bind to the AAV8 and AAV9 virions. These results suggest that the CRP-mediated enhancement of transduction is species/serotype dependent.
Serum protein blocking of AAV transduction
The most common serum proteins that interact with rAAV vectors and block their transduction are Nabs. 13 Unfortunately, the high prevalence of pre-existing Nabs against AAVs in the general human population has highly influenced AAV-based gene therapy in the current clinical trials. 40 –42
Although several serum proteins have demonstrated the ability to increase the transduction efficacy of rAAV vectors in vivo, some other serum proteins may play negative roles in AAV transduction based on the AAV serotype and the species from which the sera are derived. For example, galectin 3 binding proteins (G3BPs) from human and dog sera, but not from macaque and mouse sera, can strongly interact with AAV6. Interestingly, human G3BP blocked AAV6 transduction efficiency in mice through the formation of aggregates between G3BP and AAV6. 33 Another study showed that PF4 from mouse and human sera interacts with both AAV8 and rAAV9 vectors. After administering rAAV vectors preincubated with human serum, lower heart transduction was observed in the wild-type mice compared with that in PF4-deficient mice. There was no difference in transduction in the other tissues, such as liver and muscle. 43 This result is inconsistent with our finding that the preincubation of AAV9 with human PF4 protein enhanced global transduction, including the heart. 32 Several possibilities can explain this discrepancy, including different approaches for testing (knockout mice vs. preincubation), interference of other contaminated proteins, different ratios of serum protein to AAV virions, and the sensitivity of quantitative polymerase chain reaction for the AAV genome.
Effect of Nonserum Proteins on AAV Transduction Enhancement
In addition to serum proteins, it was also found that nonserum proteins have the ability to augment AAV transduction. Our recent study demonstrated that the FerA domains, derived from diverse Ferlin proteins (dysferlin, myoferlin, and otoferlin), increase the transduction of multiple AAV serotypes (AAV6, 8, and 9) through direct interaction of the FerA domain with AAV virions. 44 FerA is a membrane-associating α-helical bundle domain that is ∼15 kD and has been shown to possess Ca2+-dependent phospholipid binding activity. 45 The application of different rAAV vectors preincubated with various FerA domains dramatically augmented AAV transduction via either systemic or intramuscular administration. Furthermore, both mouse- and human-derived FerA domains can increase AAV transduction. We administered AAV8/hFIX vectors preincubated with the human dysferlin-derived FerA domain in a hemophilia B mouse model and found that the FerA domain boosted FIX expression and activity compared with AAV8/hFIX alone, subsequently improving hemostasis in hemophilia B mice. 44 Unlike some serum proteins with a serotype-specific or species-specific dependency, FerA domains enhance AAV transduction independent of the AAV serotype and species.
Therapeutic Potency of Peptides in AAV Gene Therapy
As highly selective and biologically versatile compounds, peptides have also drawn considerable interest as therapeutics in the development of pharmaceuticals because of their functionality being similar to that of proteins. 46 –49 Compared with proteins, peptides exhibit relatively safer, better tolerated, and more efficacious properties in view of their great pharmacodynamic profiles, such as shorter length, higher chemical versatility, and lower immunogenicity. 31,47 Consequently, peptides have enabled the delivery of various proteins, antibodies, small interfering RNAs (siRNAs), liposomes, and nanoparticles in vivo. 31,50 –52 Furthermore, peptides are typically characterized by lower production complexity and corresponding cost-effectiveness. Thus far, over 60 peptide-based drugs have been approved for clinical use, and hundreds of clinical trials for additional peptide-based therapeutics are currently underway. 53,54
Many steps are required for effective viral delivery and transduction, and interactions between AAV capsid proteins and cellular proteins are critical for this process. In theory, peptides could structurally and functionally intervene in a reciprocal manner to further affect AAV transduction efficacy.
Cell-penetrating peptides for the enhancement of AAV transduction
Cell-penetrating peptides (CPPs) are protein-derived, chimeric, or synthetic short oligopeptides (usually <40 amino acids) with high variable conformations. 55 They are characterized by an intrinsic ability to penetrate cell membranes in a noninvasive and nontoxic manner. Therefore, CPPs have been utilized to deliver various cargoes (nanoparticles, small molecules, and siRNAs). 56 A classic example is the identification of the human immunodeficiency virus type 1-derived TAT peptide, which is one of the best-studied CPPs for brain delivery. 57,58 Liu et al. have shown that several CPPs, such as TAT-HA2, Antp, and LAH4, can significantly enhance AAV2 and AAV8 transduction in primary cells/tissues by facilitating vector internalization into and escape from endosomal membranes, and even more importantly, the incubated complex of CPPs and AAV8 endows higher muscle transduction after intramuscular administration. 59 We also found that the incubation of LAH4 or TAT with AAV8 enhances liver transduction. 60
Blood–brain–barrier shuttle peptides for the enhancement of AAV transduction
It is well known that the key challenge in the systemic administration of AAV for neurological diseases surrounds how to surmount the blood–brain–barrier (BBB). BBB shuttle peptides can transport various cargoes into the brain and maintain BBB integrity; moreover, unlike endogenous proteins, they do not compete with the endogenous substrates. 31 Notably, several targeted peptides with the ability to cross the BBB for brain delivery have been identified. For example, TAT has been suggested as a BBB shuttle peptide for reaching the brain parenchyma in an active form. 61 Angiopep-2 exhibits excellent transcytosis capacity and effectively transports a wide variety of small molecules across the BBB. 62 The THR peptide can be conjugated with gold nanoparticles to increase the permeability of the conjugate in the brain. 52 Our study demonstrated that several BBB shuttle peptides could effectively bind to AAV8 and enhance its transduction in the brain, especially in neuron cells following systemic administration. 60 Furthermore, we identified a novel BBB shuttle peptide using a combination of in vivo and in vitro phage library screenings. The customized peptide could bind to AAV9 and specifically enhance its brain transduction by traversing the BBB (unpublished data).
Thus far, tremendous countermeasures have been devoted to surmounting the various intrinsic drawbacks of peptide-based therapeutic applications, such as variable solubility, in vivo instability, proteolytic degradation, and reduced bioavailability. 31,54,63 These parameters substantially influence the ability of peptides to bind to AAV virions for AAV transduction enhancement and merit further investigation.
Mechanistic Insights Regarding the Application of Proteins and Peptides in AAV Transduction
During AAV transduction, following attachment to the receptors/coreceptors on the cell surface, the virus undergoes internalization and subsequent intracellular trafficking from the early (EEs)/late endosomes (LEs) to the nucleus. Therein, the AAV virions are uncoated, and the viral genome is released. In consideration of the intrinsic and diverse properties of proteins and peptides, the mechanisms affecting AAV transduction may vary. At present, proteins and peptides involving several mechanisms that may be related to the multiple steps of the AAV life cycle as well as other strategies have predominantly been exploited (Fig. 1).
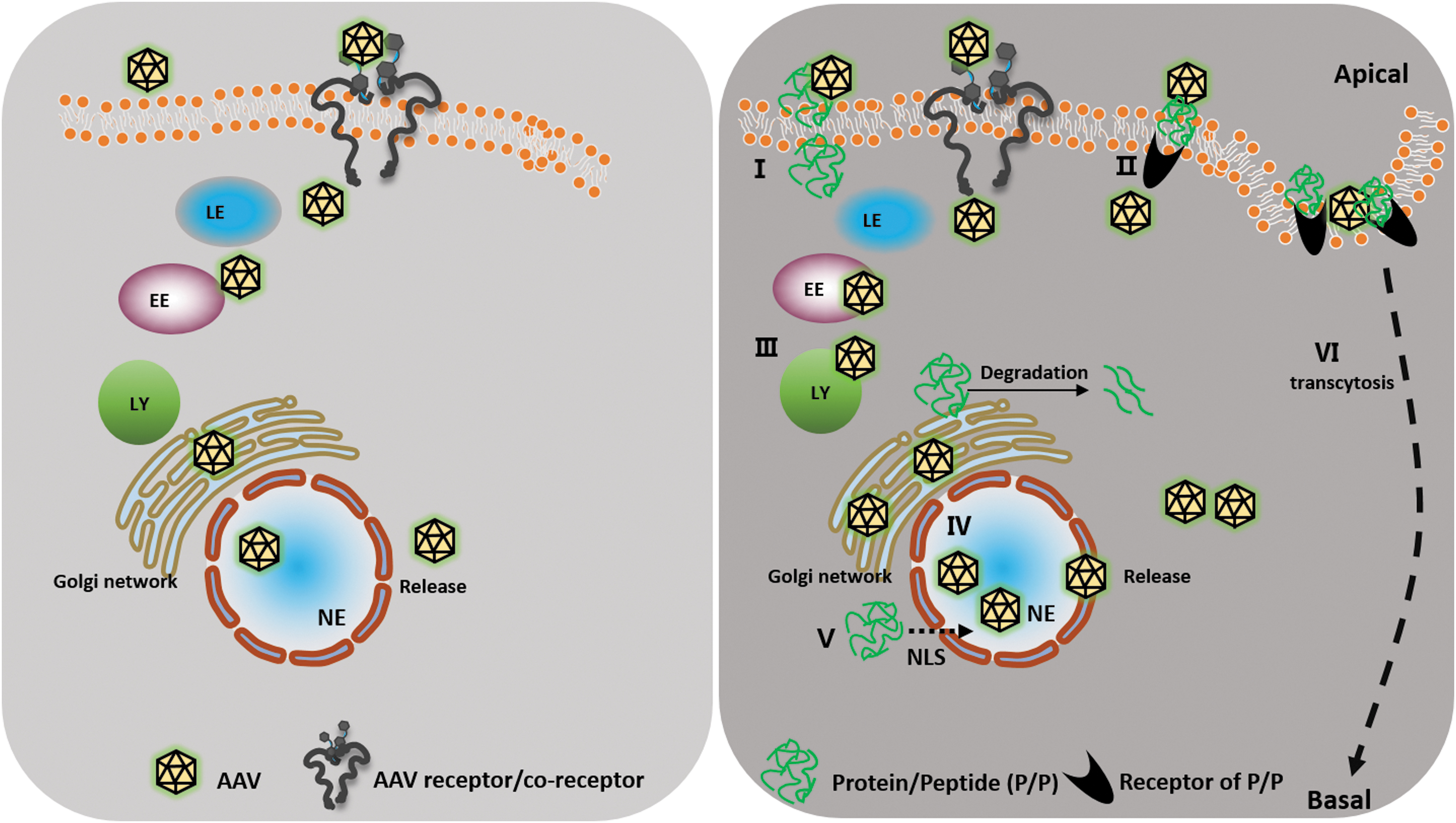
A schematic outlining of the potential mechanisms of protein/peptide-promoted rAAV vector transduction. (I) P/P directly enhances rAAV binding via the AAV receptor or coreceptors. (II) P/P indirectly increases rAAV binding via the respective P/P receptors. (III) P/P facilitates the rAAV trafficking pathway through the endosome/lysosome/Golgi network transport. (IV) P/P promotes the perinuclear/nuclear accumulation of rAAV. (V) P/P acts as an NLS and potentially promotes the nuclear transport of rAAV. (VI) P/P upregulates the rAAV transcytosis ability. EE, early endosome; LE, late endosome; LY, lysosome; NE, nucleus; NLS, nuclear localization signal; P/P, protein/peptide; rAAV, recombinant adeno-associated virus.
Binding
Binding is the first step of AAV infection in cells. Proteins/peptides can facilitate the AAV binding process by multiple means. Some proteins or peptides, such as LDL, transferrin, and THR, functionally possess their own receptors on the cell surface. Therefore, the proteins/peptides can utilize their respective receptors and increase AAV binding to the cells for enhanced transduction. Our recent studies have shown that the preincubation of AAV with THR peptide, serum protein albumin, LDL, transferrin, or FerA protein enhances AAV transduction by increasing viral binding ability. 36,44,60 It has been also shown that the binding of peptides or proteins with virions does not interfere with AAV infection biology, 44,60 which suggests that the peptide binding sites on the AAV surface may be not the same binding regions/domains for target cell interactions. It is also possible that the AAV virion-binding proteins will facilitate AAV interactions with target cells via the AAV receptor or coreceptors. There is currently no direct evidence to support this mechanism; thus, it requires future investigation.
Internalization and intracellular trafficking
Internalization is the subsequent step after viral binding. Typically, CPPs are characterized by an internalization ability to deliver their conjugates by translocation across the cell membrane. 64 Liu et al. have shown that the LAH4 peptide can increase the internalization of AAV2 by facilitating energy-independent/-dependent cellular uptake. 59 Furthermore, proteins/peptides can also facilitate AAV intracellular trafficking from the EEs/LEs to the nucleus. For example, we showed that FerA proteins fueled the intracellular trafficking process of AAV, which contributed to the enhancement of AAV transduction. 44 In addition, several CPPs possess a nuclear localization signal and may assist with AAV nuclear entry. 65,66 In summary, peptides/proteins may potentially be used in conjunction with AAV to enhance transduction.
Transcytosis
As the process for transferring molecules across various cell types, transcytosis may play an important role in AAV transduction. Our study has shown that the BBB shuttle peptide THR increased the ability of AAV8 to cross the endothelial cell barrier through transferrin receptor 1-mediated transcytosis so that rAAV vectors transduced more neuron cells in the mouse brain. 60 Moreover, it was shown that the serum protein fibrinogen enhanced the global transduction of AAV9 by increasing the transcytosis of rAAV9 vectors. 32 Therefore, the use of transcytosis-enhancing peptides and proteins may be a promising approach for enhancing AAV transduction, especially for boosting transduction in the brain.
Parameters that Impact the Effect of Native Proteins/Peptides in AAV Transduction
Natural proteins/peptides have been reported to increase the transduction of rAAV vectors. However, the details of protein/peptide treatments and rAAV vectors have rarely been investigated. Here, we summarized some important parameters that have been reported to impact AAV transduction. The information will be very helpful for the application of peptides/proteins with rAAV vectors.
Incubation duration and temperature of proteins/peptides and rAAV vectors
The proper incubation of proteins/peptides and rAAV vectors may form an envelope-like structure for AAV transduction. Therefore, it is necessary to determine an appropriate duration and temperature for the incubation of proteins/peptides with AAV. It has frequently been observed that AAV transduction enhancement is only achieved in the groups for which the rAAV vectors are preincubated with proteins/peptides before injection into the mice; therefore, effective incubation is a prerequisite for enhancing AAV transduction. Liu et al. performed the incubation of CPP and AAV virions at 37°C for 30 min and achieved enhanced AAV transduction. 59 Denard et al. preincubated several proteins and AAVs at room temperature for 1 h before administration in vivo. 33,43 Wang et al. found that the enhancement of AAV transduction could be observed with a short incubation at 4°C for 15 min. Nevertheless, the highest enhancement was achieved when albumin and AAV8 virions were incubated at 4°C for 2 h. 34 In our subsequent studies, the stable complexes of AAV virions and proteins/peptides incubated at 4°C for 2 h could bolster AAV transduction. 32,44,60 These findings suggest that the duration and temperature of incubation should be considered based on the properties of the selected proteins or peptides.
Molecular ratios of proteins/peptides to AAV virions
It is worth noting that the number of proteins or peptides on the AAV surface is a pivotal property for enhanced transduction. The uptake of AAV particles is nonlinear in accordance with increasing the number of proteins/peptides. We speculated that both AAV serotype and the molecular ratio between AAV virions and proteins/peptides could influence the enhancement of AAV transduction. For instance, Liu et al. reported that the average ratio of AAV2 and CPP peptides ranges from 1:2,000 to 1:3,000. 59 In our studies, the optimal ratio of AAV8/9 and peptides or proteins was ∼1:2,000 to 1:20,000, and a molecular ratio of rAAV vectors to proteins/peptides greater than 1:20,000 did not appear to further enhance AAV transduction. Thus, an inadequate number of protein/peptide molecules could not achieve AAV transduction enhancement. These results suggest that AAV enhancement reaches a peak when the concentration of proteins/peptides becomes too saturated. The optimal ratio of AAV virions to proteins/peptides may be attributable to the affinity between the protein/peptide and the AAV capsid and their respective distinct structures. In addition, polarity/electrostatic forces also play a role in the interactions of proteins/peptides with AAV virions. 59
Concluding Remarks and Future Considerations
Therapeutic proteins and peptides have a wide range of applications in modern medicine and biotechnology. Meanwhile, substantial preclinical studies and clinical trials have demonstrated that the rAAV vector is a promising vehicle for gene therapy. Thus, purposeful exploration of peptide- and small protein-based design strategies will undoubtedly accelerate and broaden the development of AAV-based gene therapy. Many drugs, such as triptolide T10, hydroxyurea, hydroxychloroquine, and proteasome inhibitors, have also been reported to enhance AAV transduction in vitro and in vivo. 27,67 –69 Compared with the nonprotein drugs, proteins/peptides could be explored as customized “adjuvants” for specific tissues or AAV serotypes without the unwanted side effects of pharmacological reagents. The elaboration of the AAV structure-directed design and the discovery of therapeutic proteins, peptides, and even their conjugates are emerging as attractive new tools to aid and enhance gene delivery. Proteins and peptides could be also explored as a “biobridge” to connect AAV and the related promising molecules to broaden the application in preclinical/clinical trials. Exploring how to mitigate the weaknesses of protein/peptide-based AAV gene therapy and enhancing specific targeted tropisms should be the next concerns and focuses.
In the meantime, the potential immune response induced by exogenous proteins/peptides needs to be considered. Most of native proteins that exist endogenously cannot elicit immune responses in human beings. It is also unlikely for peptides to mount a humoral immune response owing to their short length (usually ∼8–12 amino acids). However, when the complexes of AAV virions and exogenous proteins/peptides are taken up by antigen-presenting cells, it is possible to induce protein/peptide-specific CTL responses. The details of this process require additional investigation. Furthermore, it is also imperative to overcome the intrinsic weaknesses of proteins/peptides, such as the tendency for aggregation in vivo, a short circulating plasma half-life, and potential tissue toxicity. In addition, since the proteins or peptides will be synthesized in vitro, potential contamination by exogenous materials from bacterial/mammalian/yeast expression systems during the manufacturing process may present risks. As these concerns apply to all protein/peptide-based therapeutics, they should be resolved in accordance with the FDA guidelines before entering clinical trials.
Direct interactions of native proteins or peptides with AAV virions are essential for transduction enhancement. However, several issues require further investigation: the binding domains on the AAV virion surface and in proteins/peptides that have not been identified, the protein/peptide affinities for specific AAV serotypes, and the stability of the AAV/protein or AAV/peptide complexes in blood and during intracellular trafficking, especially in endosomes/lysosomes with low pH levels. Notably, the role of protein/peptide is only explored for five commonly used AAV serotypes in this review. Currently, 13 serotypes and over 100 variants or mutants of AAV have been developed as gene delivery vehicles. It is worth investigating whether diverse proteins/peptides could specifically affect the transduction of different AAV capsids, which will expand the pool of proteins/peptides for future clinical application. Recently, two groups have identified the lymphocyte antigen six-complex, locus A that could specifically bind to the neurotropic AAV9 variant AAV-PHP.B and dramatically promote viral transport across the BBB in a mouse strain-specific manner. 70,71 Another potential field requiring exploration involves the design of more effective molecules, such as fusion peptides or proteins with binding and intracellular trafficking capabilities, to further enhance AAV transduction. One recent study demonstrated that a synthetic CPP, consisting of the HIV-TAT sequence for nuclear entrance and an epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor peptide for blockage of EGFR signaling, effectively increased AAV2 transduction efficiency in the high EGFR-expressed fibroblast cell line RAT-1. 72
In summary, protein and/or peptide-based AAV transduction enhancement holds promise for broadening the field of AAV-based therapeutics in clinical applications, in the near future. This work presents a novel and effective strategy to enhance AAV efficacy without alterations in rAAV vector tropism. Future studies should focus on designing and further refining multifunctional proteins and peptides with the ability to bind to rAAV vectors for tissue-specific targeting or to prevent off-targeted AAV transduction.
Footnotes
Acknowledgments
The authors thank Amanda Lee Dobbins, Ellie Azure Frost, and Taylor Sheldon Ralph in our laboratory for their critical reviews of the article.
Author Disclosure
R.J.S. is the founder and a shareholder at Asklepios BioPharmaceutical and Bamboo Therapeutics, Inc. He holds patents that have been licensed by UNC-Chapel Hill to Asklepios BioPharmaceutical, for which he receives royalties. He has consulted for Baxter Healthcare and received payment for speaking. C.L. is a cofounder of Bedrock Therapeutics, Inc., He holds patents licensed by UNC-Chapel Hill and has received royalties from Bedrock Therapeutics and Asklepios BioPharmaceutical.
Funding Information
This work was supported by the National Institutes of Health grants R01AI117408, R01HL144661, and R01HL125749.