
Abstract
Lentiviral vectors are one of the most commonly used viral delivery systems for gene therapy. Vesicular stomatitis virus-G envelope glycoprotein (VSV G)-pseudotyped lentiviral vectors have been widely used in clinical studies for treatment of virus infections and genetic deficient diseases. However, the efficiency of lentiviral vector transduction has been long recognized as a limiting factor in clinical gene therapy application, especially in transducing hematopoietic stem cells. MARCH8 (membrane-associated RING-CH 8), an E3 ubiquitin ligase, has been reported to target and downregulate VSV G. Results in this study show that MARCH8 induces ubiquitination and lysosome degradation of VSV G, and knockout of MARCH8 in virus-producing cells increases lentiviral vector transduction by elevating the level of VSV G protein. We then engineered VSV G mutant that has the lysine residues in the cytoplasmic domain substituted for arginine, and showed that this G mutant resists degradation by MARCH8, and allows the enhancement of transduction efficiency of lentiviral vector particles than the parental VSV G protein. This engineered VSV G mutant thus further advances the lentiviral vector system as a powerful tool in gene therapy.
Introduction
Lentiviral vector is one of the most prominent tools for stable gene delivery. It has been approved in clinical applications as gene therapeutics. Because of their capacity of transducing dividing and nondividing cells, and accommodating relatively large transgene fragments, lentiviral vectors have become one of the few gene delivery systems commonly used in research and in clinics. 1 –6 The recent generation of lentiviral vectors show relatively safe integration profiles in host chromosomes, and generate long-term expression of the delivered genes, while induce only moderate immune responses. 7 –11 Therefore, they have been widely used for treatment of virus infections, cure of genetic deficient diseases, and generation of chimeric antigen receptors or epitope-specific transgenic T cell receptors T cells for cancer immune therapies. 12 –29
Lentiviral vector system consists of a set of plasmids, the packaging DNA construct encoding Gag/Gag-Pol and HIV regulatory protein Rev, the envelope protein, and a transgene transfer vector. 4,6,30,31 Stable producer cell lines were also used to produce batches of high titer LV for large clinical trials. 32 –36 VSV G (vesicular stomatitis virus-G envelope glycoprotein) is one of the most commonly used viral envelope proteins. It binds to the low-density lipoprotein receptor (LDLR), a ubiquitously expressed cell surface protein, thus warrants a broad spectrum of target cells. 37 –39 VSV G is stable with virus particles, and allows the application of ultracentrifugation and column chromatography to produce high-titer virus stocks. 38,40
While having a number of advantages for gene delivery, lentiviral vectors still have several weaknesses that await further improvement. One issue is the low transduction efficiency of resting T lymphocytes, B cells and hematopoietic stem cells (HSCs) due to the low level of LDLR, cell cycle quiescence, proteasome-mediated inhibition, and high level of restriction factors such as IFITM3. 41 –54 Increasing transduction efficiency presents one strategy to deliver genes into these cell types. Potential solutions include cytokine stimulation, increasing multiplicity of infection (MOI), sequential transductions, and genetic modification of VSV G. 53,55 –59 Recent studies found that agents such as mammalian target of rapamycin (mTOR) inhibitor, caraphenol A, cyclosporine H could overcome IFITM3 antiviral activity and enhance the transduction of stem cells. 49,60 –62
VSV G was recently reported to be downregulated by human MARCH8 (hMARCH8). 63 MARCH8 belongs to the MARCH (membrane-associated RING-CH) family, which were first discovered in virus as E3 ubiquitin (Ub) ligases and downregulated MHC class I molecule (MHC I). 64,65 Mammalian MARCH proteins were subsequently shown to modulate plasma membrane proteins turnover. 66 –70 For example, cell surface MHC I, MHC II, CD44, and CD98 were reduced by hMARCH8. 66,67,71 –73
MARCH8 was shown to inhibit the incorporation of HIV-1 Env and VSV G into viral particles. 63 A recent report demonstrated that MARCH8 targeted HIV-1 Env and VSV G by two different mechanisms: MARCH8 downregulated HIV-1 Env from cell surface through tyrosine motif-dependent pathway, while induced VSV G lysosome degradation by ubiquitination-dependent pathway. 74 In this study, we report that hMARCH8 inhibits lentiviral vector transduction through downregulating VSV G expression, and knockout of hMARCH8 in virus-producing cells enhances lentiviral vector transduction efficiency. Furthermore, we engineered the hMARCH8-resistant VSV G mutant V5R, which enhances transduction efficiency of lentiviral vector. Our findings thus provide one tool to further advance the clinical application of lentiviral vectors.
Materials and Methods
Cells
293T cells and HeLa cells were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS; Gibco), 1% penicillin (100 U/mL) and streptomycin (100 μg/mL) (Solarbio). SupT1 cells and THP1 cells were cultured in complete RPMI 1640 supplemented with 10% FBS (Gibco). Peripheral blood mononuclear cells (PBMCs) isolated from healthy donors were purchased from AllCells (Shanghai, China), and cultured in RPMI 1640 media containing 10% FBS (Gibco). Frozen granulocyte colony stimulating factor-mobilized peripheral blood (mPB) CD34+ cells were purchased from AllCells (Shanghai, China). This research is exempt from approval to allow use for research purposes only. mPB-CD34+ cells were cultured in StemSpan™ SFEM II (STEMCELL), supplemented with recombinant human cytokines thrombopoietin, Flt3L, IL3, and stem cell factor (STEMCELL). hMARCH8 knockout 293T cells were created using the CRISPR-Cas9 system. 293T cells were transfected with lentiCRISPR v2 (Addgene) control vector or CRISPR-Cas9 carrying guide RNAs that target hMARCH8 (g1, GTAAGACCAAAGAAAAGGAG; g3, GTGCGAGAGAAGGAGGACAC), followed by selection with puromycin (0.6 μg/mL; Sigma-Aldrich). Pools of puromycin-resistant cells were diluted and plated in 96-well plates to obtain single-cell clones. The genotype of knockout clones was confirmed by sequencing.
Plasmids and antibodies
The cDNAs of hMARCH8 and W114A mutant, pLenti-GFP (green fluorescent protein) were kindly provided by Dr. Zhaohui Qian. 75 Nine different Ub expression plasmids were obtained from Addgene. Lysine mutants of VSV G were generated with KOD-Plus-Mutagenesis Kit (TOYOBO). Plasmid DNA was transfected into cells using polyethyleneimine (Sigma-Aldrich) in accordance with the manufacturer's instruction. The following antibodies were used in this study: anti-VSV G (V5507; Sigma-Aldrich), anti-Flag antibody (F3165; Sigma-Aldrich), anti-β-actin (A1978; Sigma-Aldrich), anti-p24 (11695-RP01; Sino Biological), anti-MARCH8 (14119-1-AP; Proteintech), anti-HA tag (hemagglutinin tag) (H6908; Sigma-Aldrich), and anti-GFP (50430-2-AP; Proteintech).
Viral vector production and transduction
293T cells were seeded in 10-cm dishes in DMEM with 10% FBS, then transfected with 3:3:1 of psPAX2 encoding the structural polyproteins Gag-Pol, pLenti-GFP containing the GFP and luciferase reporters, and pMD.G encoding VSV G envelope, in the presence or absence of hMARCH8. After 48 h, supernatants were clarified by centrifugation, passed through a 0.45 μm filter, and measured with p24-antigen capture enzyme-linked immunosorbent assay (ELISA). Cells were transduced with 10 ng of p24 antigen of hMARCH8-expressing lentiviral vector or the control vector for 72 h, and then harvested for luciferase assay and flow cytometry. The relative luciferase activities were determined using a Luciferase Assay System (Promega) in a Modulus Microplate Reader (Turner Biosystems). GFP signal was determined with FACS Canto II (BD).
Flow cytometry
Cells transfected with VSV G and hMARCH8 or control vector were suspended in phosphate-buffered saline containing 1% paraformaldehyde (Solarbio), permeabilized with 0.1% Triton X-100, and immunostained with the antibody against VSV G (Sigma-Aldrich), followed by incubation with anti-mouse Alexa Fluor® 488-conjugated antibody (Thermo Scientific). VSV G expression was analyzed with FACS Canto II (BD).
Immunofluorescence staining and confocal microscopy
HeLa cells transfected with VSV G and Flag-hMARCH8 or control vector were fixed in 4% paraformaldehyde, and permeabilized with 0.1% Triton X-100 for 10 min. Then cells were stained with anti-VSV G antibody (Sigma-Aldrich) and anti-Flag antibody (Sigma-Aldrich) for 1 h, followed by incubation with anti-mouse IgG conjugated with Alexa Fluor 488 (Thermo Scientific), and anti-rabbit IgG conjugated with Alexa Fluor 594 (Thermo Scientific). Nuclei were stained by DAPI. A Leica TCS SP5 inverted microscope (Leica Microsystems) were used to acquire images.
Western blotting and immunoprecipitation
Western blotting was performed as described previously. 76 Cells were lysed in RIPA buffer containing 25 mM Tris/HCl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1% NP-40, 0.25% sodium deoxycholate, and a proteinase inhibitor cocktail (Sigma-Aldrich). After centrifugation, proteins in lysates were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred onto the nitrocellulose membranes (Millipore). Membranes were probed with indicated antibodies, followed by incubation with IRDye secondary antibodies (Li-Cor). Then membranes were scanned with Odyssey Infrared Imaging System (Li-Cor).
For immunoprecipitation and ubiquitination assay, 293T cells in 10-cm dishes were transfected with 2 μg HA-Ub, 3 μg VSV G, and 1 μg hMARCH8 or W114A mutant. After 48 h, cells were harvested and lysed in RIPA buffer. After clarification with centrifugation, cell lysates were incubated with anti-VSV G Ab and Dynabeads™ magnetic beads (Invitrogen) at 4°C for 4 h. After washing with RIPA buffer, immunoprecipitates were denatured by heating in RIPA buffer containing 1% SDS for 5 min. The elutes were diluted with RIPA buffer and reimmunoprecipitated with anti-VSV G Ab and Dynabeads magnetic beads. After extensive wash, agarose was boiled in SDS-PAGE sample buffer, and samples were analyzed by Western blotting as described above.
BlaM-Vpr assay
BlaM-Vpr pseudoviral particle fusion assay was performed as previously described. 77,78 In brief, cells were transfected with VSV G, psPAX2, and BlaM-Vpr. Forty-eight hours after transfection, 293T cells were incubated with BlaM-Vpr pseudoviral particles for 8 h, and then loaded with CCF2-AM substrate (Invitrogen) for 2 h according to the manufacturer's instruction. Following substrate loading, cells were collected for flow cytometry analysis.
Statistical analysis
All data were analyzed using Microsoft Excel or GraphPad Prism 5. Statistical significance was determined by Student's t-test, *p < 0.05; **p < 0.01.
Results
hMARCH8 affects VSV G levels and diminishes the transduction efficiency of lentiviral vectors
VSV G is an essential component of the lentiviral vector system, and is critical for the transduction efficiency of lentiviral vectors. Consistent with the previous report, 63,74 VSV G was downregulated by hMARCH8 in a dose-dependent manner, no effect was observed for GFP (Fig. 1A). Immunofluorescence imaging of VSV G further showed that VSV G expression was decreased in hMARCH8-expressing cells (Fig. 1B), which was confirmed by flow cytometry analysis of VSV G levels (Fig. 1C). Given that hMARCH8 reduces VSV G levels, we tested whether hMARCH8 diminishes the transduction efficiency of lentiviral vector by producing VSV G-pseudotyped lentiviruses from 293T cells that transiently expressing hMARCH8. As shown in Fig. 1D, levels of VSV G in virus-producing cells and in the progeny viruses decreased as a result of hMARCH8 expression (Supplementary Fig. S1A), with similar levels of viral p24 proteins as shown by the result of p24-antigen capture ELISA (Fig. 1E and Supplementary Fig. S1B). 293T cells and SupT1 cells were transduced with lentiviral vector of the same viral p24 amounts, and transduction efficiency was evaluated with luciferase assay and flow cytometry. The results showed that hMARCH8 strongly inhibited lentiviral vector transduction of 293T (Fig. 1F, G) and SupT1 cells (Fig. 1H, I) in a dose-dependent manner (Supplementary Fig. S1C–F). The lowest dose of hMARCH8 DNA (0.05 μg) already diminished the transduction efficiency by 70% (Supplementary Fig. S1), consistent with previous report that hMARCH8 profoundly reduced infectivity of VSV G pseudotyped virus. 63 This decrease of transduction was caused by the inhibition of VSV G expression and impaired incorporation into virions in MARCH8-expressing producer cells.
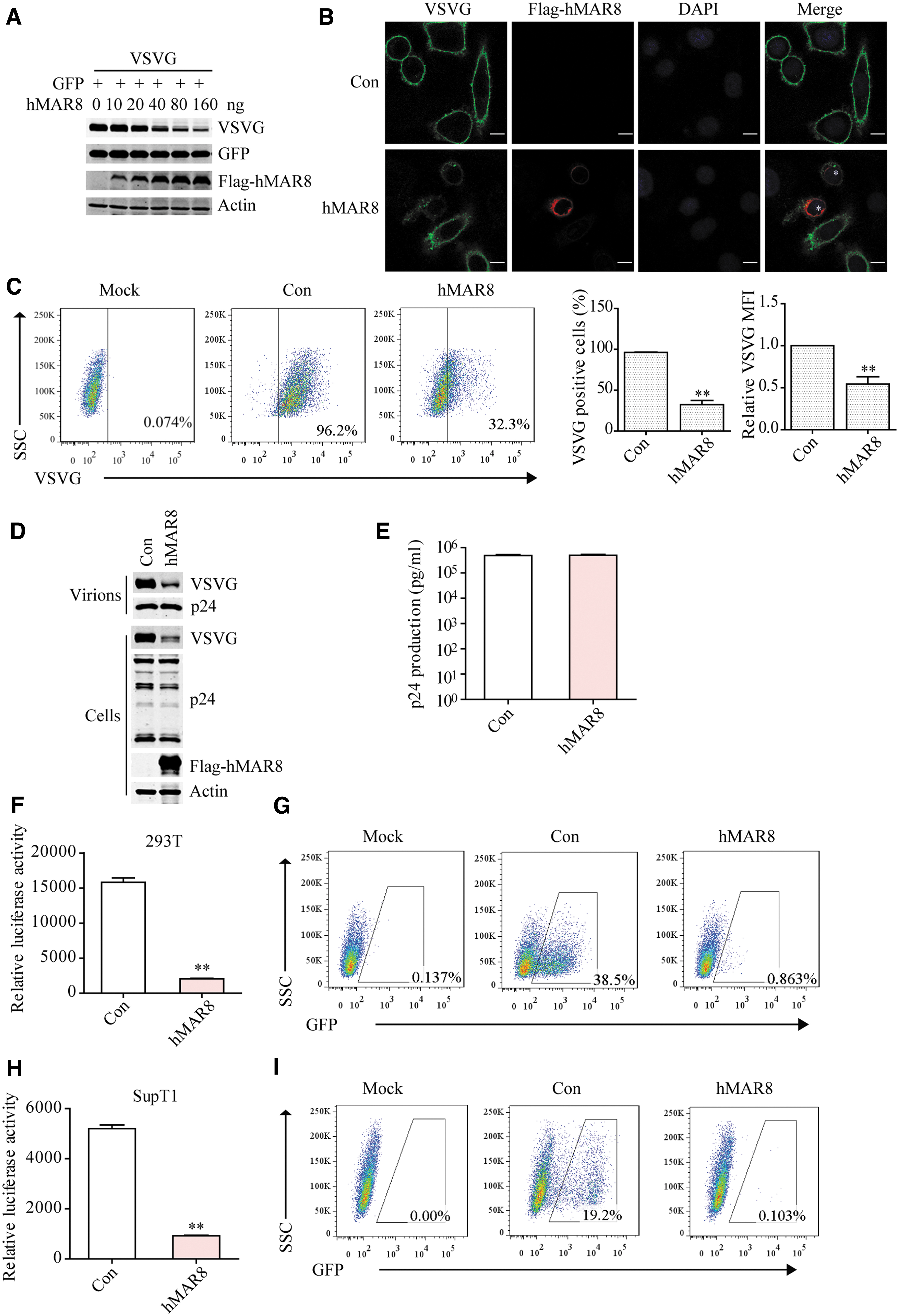
hMARCH8 reduces VSV G expression.
Knockout of hMARCH8 in virus-producing cells enhances the transduction property of lentiviral vector
We next created hMARCH8-knockout 293T cells using CRISPR-Cas9 and investigated the effect of endogenous hMARCH8 on the transduction activity of lentiviral vectors. The genotype of knockout clones was analyzed by sequencing (Supplementary Fig. S2A). VSV G-pseudotyped lentiviruses were produced in hMARCH8-knockout 293T cells. As shown in Fig. 2A, the expression of MARCH8 was depleted in hMARCH8-knockout 293T cells, and the level of VSV G increased. hMARCH8 knockout did not affect virus production (Fig. 2B). Results of luciferase assay showed that knockout of hMARCH8 in virus-producing cells enhanced the transduction efficiency of lentiviral vectors in different cell types, including 293T cells (Fig. 2C), SupT1 cells (Fig. 2D), THP1 cells (Fig. 2E), PBMCs (Fig. 2F), and human mPB-CD34+ cells (Fig. 2G) by twofold to fourfold. The enhancement of transduction efficiency using hMARCH8-knockout cells was still observed with exogenous hMARCH8 expression (Supplementary Fig. S2B–D). Together, these data support the inhibitory effect of hMARCH8 on VSV G levels and the transduction property of lentiviral vectors.
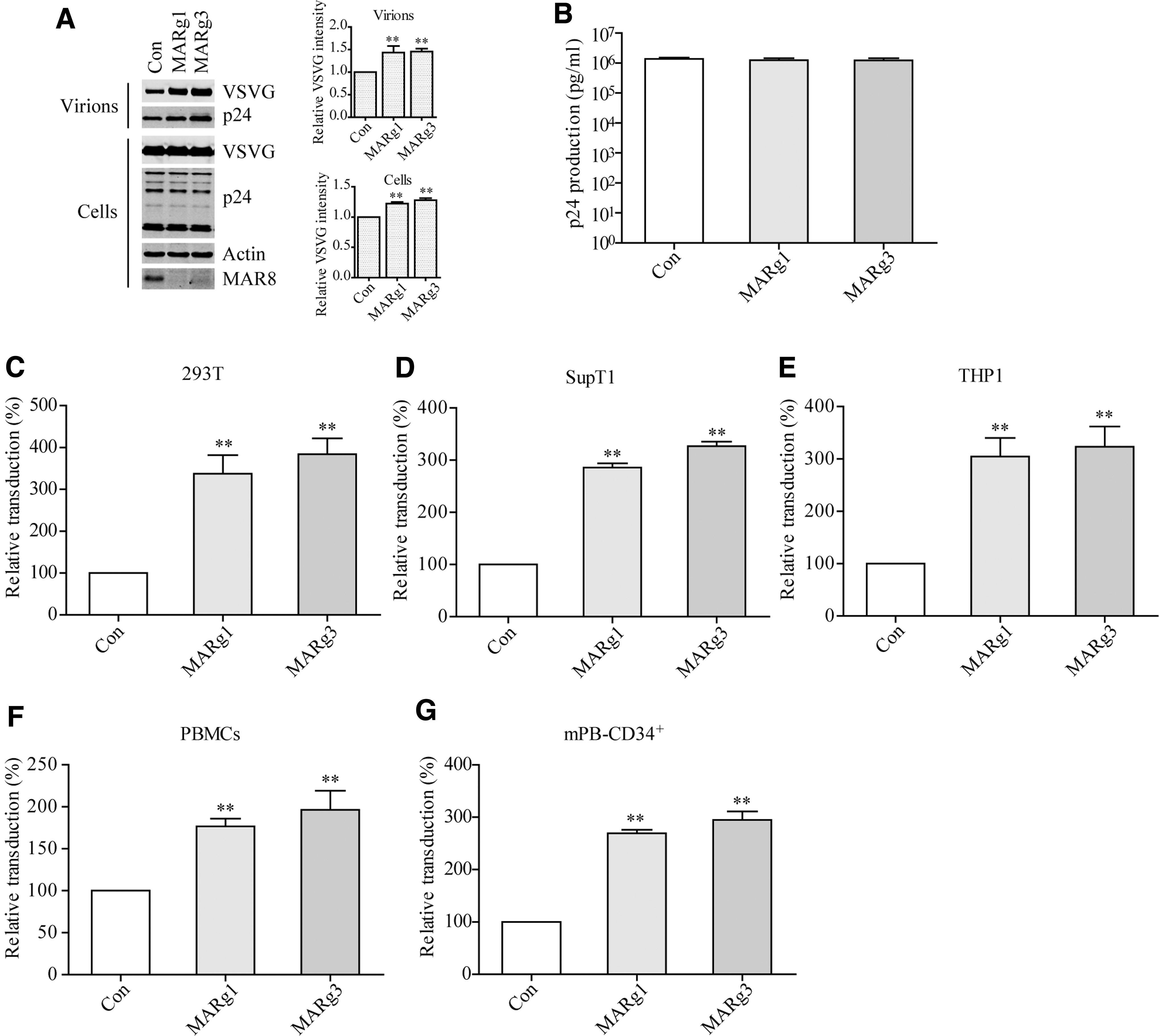
Knockout of hMARCH8 in virus-producing cells enhances the transduction efficiency of lentiviral particles.
hMARCH8 induces ubiquitination and lysosome degradation of VSV G
MARCH8 downregulates several immunomodulatory proteins, such as MHC II, CD44, CD88, and CD98, through the proteasome or lysosome pathway via ubiquitination of the target proteins. 66,67,71 –73 To determine which degradation pathway hMARCH8 uses to degrade VSV G, we treated hMARCH8 and VSV G transfected cells with proteasome inhibitor MG132 or the lysosome inhibitor chloroquine (CQ). CQ but not MG132 inhibited degradation of VSV G by ectopic hMARCH8 (Fig. 3A) and endogenous hMARCH8 (Fig. 3B), indicating that hMARCH8-mediated degradation of VSV G depends on the lysosome pathway. hMARCH8 is an E3 ubiquitin ligase, and its W114A mutant loses the E3 ligase activity. When hMARCH8 or its W114A mutant DNA was transfected into HeLa cells together with VSV G, results of Western blotting and immunofluorescence imaging showed that VSV G was downregulated by wild-type hMARCH8 but not by the W114A mutant (Fig. 3C, D), supporting the critical role of the E3 ligase activity of hMARCH8 in VSV G degradation. Immunoprecipitation assay showed that ubiquitination of VSV G was caused by hMARCH8, not W114A mutant (Fig. 3E), consistent with a recent report. 74 Since different types of polyubiquitination occur to protein substrates, we investigated which type of ubiquitination hMARCH8 adds to VSV G. MARCH8, VSV G, and wild-type ubiquitin, or ubiquitin mutants each has only one lysine (K6, K11, K27, K29, K33, K48, K63), or K0, which has no lysine transfected in 293T cells. Then VSV G was immunoprecipitated and ubiquitin was probed with HA antibody. As shown in Fig. 3F, hMARCH8 mainly induced K27-linked ubiquitination of VSV G. Then ubiquitination assay was performed in hMARCH8 knockout 293T cells, and the results showed that ubiquitination of VSV G was decreased in hMARCH8 knockout cells (Fig. 3G), indicating ubiquitination of VSV G by endogenous hMARCH8. Together, these results demonstrate that hMARCH8 causes K27-linked ubiquitination and lysosome degradation of VSV G.

hMARCH8 causes ubiquitination and lysosome degradation of VSV G.
Ubiquitination of VSV G occurs to the five lysine residues in the cytoplasmic region
Since the ubiquitin chain is attached to lysines exposed to the cytoplasm, we mutated the five lysine residues in the cytoplasmic region of VSV G, either individually or in combinations (Fig. 4A). The results showed that hMARCH8 still drastically downregulated the 491R, 493R, 496R, 497R, and 511R VSV G mutants, whereas moderately decreased the V4R mutant (491R/493R/496R/497R) (Fig. 4B). The V5R (491R/493R/496R/497R/511R) mutant completely resisted hMARCH8 degradation (Fig. 4B, C). Immunofluorescence (Fig. 4D) and flow cytometry analysis (Fig. 4F) confirmed that VSV G expression was reduced by hMARCH8, while expression of V5R was not affected. Furthermore, the V5R VSV G mutant was refractory to hMARCH8-mediated ubiquitination (Fig. 4E).
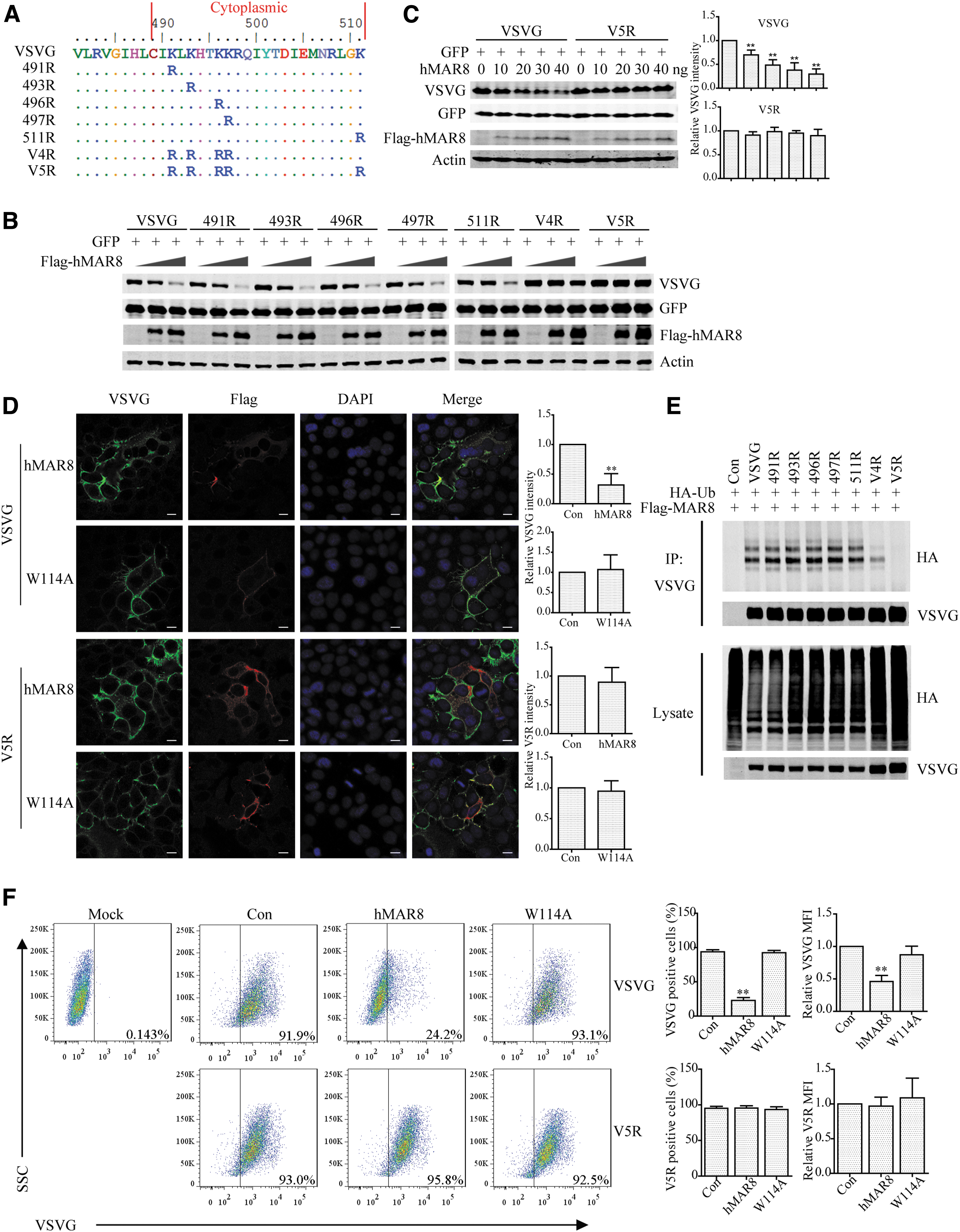
hMARCH8 causes VSV G ubiquitination at five lysines in the cytoplasmic region.
VSV G mutant V5R resists hMARCH8-mediated impairment of lentiviral vector transduction
Since the V5R mutant was resistant to degradation by hMARCH8, we asked whether V5R allows the production of hMARCH8-resistant lentiviral vectors from hMARCH8-expressing cell. We therefore produced VSV G or V5R-pseudotyped lentiviruses from 293T cells transiently expressing hMARCH8. Levels of VSV G in cell lysates and virions dramatically decreased by hMARCH8, whereas no such effect was observed for V5R (Fig. 5A). Results of p24-antigen ELISA assay showed that hMARCH8 did not affect the virion production (Fig. 5B). Results of luciferase assay showed that hMARCH8 in producer cells reduced VSV G levels and the transduction efficiency of VSV G-pseudotyped lentiviral vector (Fig. 5C, D). In contrast, transduction by the V5R-pseudotyped lentivirus was not affected by hMARCH8 (Fig. 5C, D). These results indicate that the V5R VSV G mutant resists hMARCH8-mediated inhibition of lentiviral vector transduction, consistent with this recent report showing that virus pseudotyped with VSV G K/R mutant was resistant to MARCH8. 74

V5R resists hMARCH8-mediated impairment of lentiviral vector transduction.
The V5R mutant produces lentiviral vectors of higher transduction efficiency than VSV G
Finally, we assessed whether the V5R mutant is superior to VSV G in producing more efficient lentiviral vectors. We thus produced VSV G, V4R, or V5R-pseudotyped lentiviruses by transfecting 293T cells. The levels of V4R and V5R were moderately higher than VSV G in virus particles, while p24 levels were the same (Fig. 6A, B). Results of transduction experiments showed that lentiviral vectors bearing V4R or V5R exhibited higher transduction efficiency in 293T cells (Fig. 6C), SupT1 cells (Fig. 6D), THP1 cells (Fig. 6E), PBMCs (Fig. 6F), and mPB-CD34+ cells (Fig. 6G), compared to lentiviral vector having VSV G. BlaM-Vpr assay showed that V4R or V5R allows a better entry of lentiviral vectors than VSV G (Fig. 6H). These data support the advantage of using VSV G variant to enhance the transduction efficiency of lentiviral vectors.
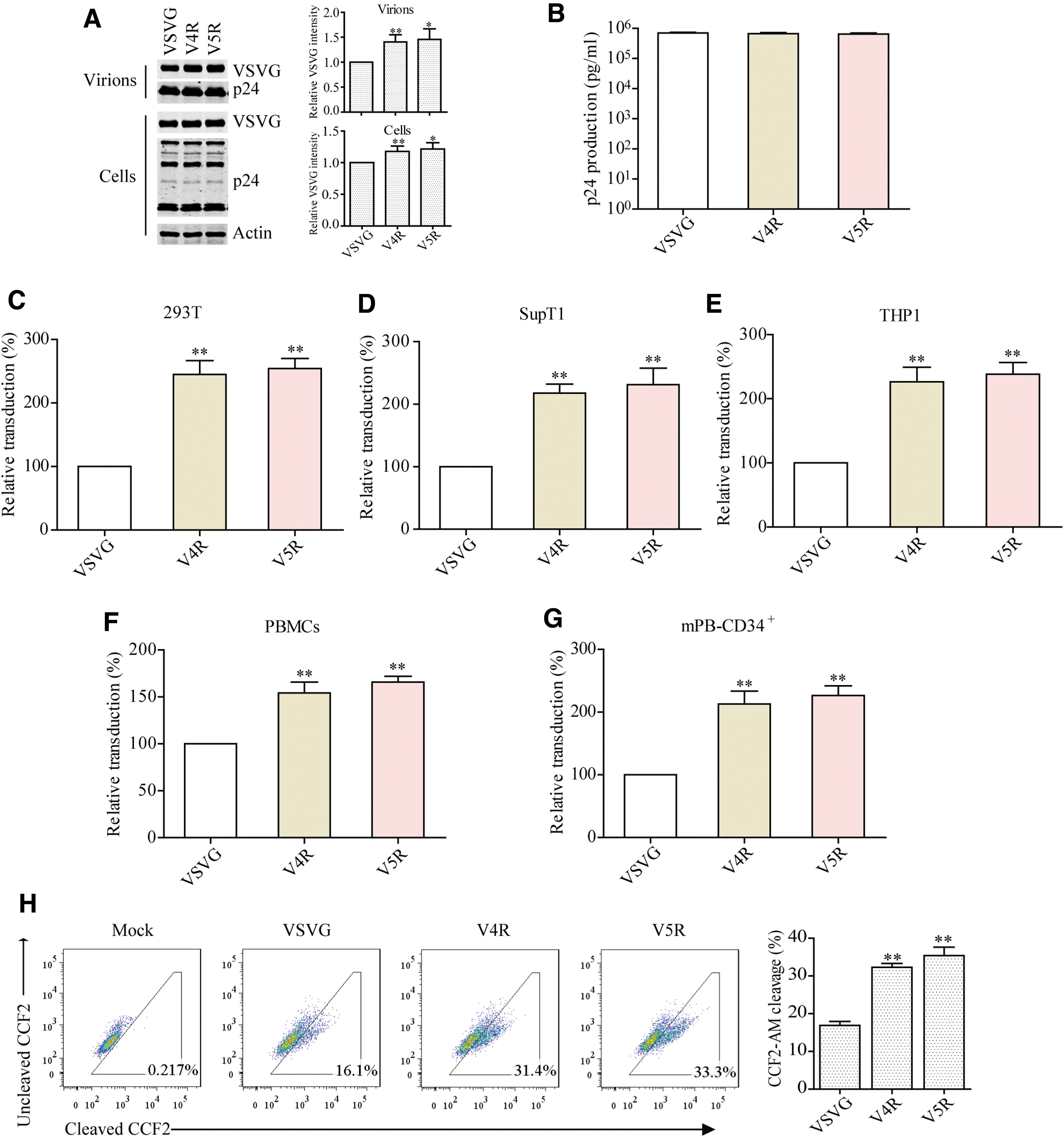
Transduction with lentiviral particles carrying VSV G mutant.
Discussion
The VSV G is one of the few viral envelope proteins commonly used in lentiviral vector system. The E3 ubiquitin ligase hMARCH8 has been shown to reduce the infectivity of VSV G-pseudotyped viruses by downregulating VSV G expression. 63 We found that even a low level of hMARCH8 expression already profoundly reduces VSV G expression. Consistent with recent report showing that MARCH8 induces VSV G ubiquitination and downregulation, 74 we found hMARCH8 specifically induces K27-linked ubiquitination of VSV G and causes lysosome-dependent degradation of VSV G. Importantly, we engineered the VSV G mutant V5R, which has the five lysine residues in the cytoplasmic region of VSV G substituted for arginines, and this mutant resists hMARCH8-mediated ubiquitination and degradation, thus increases the transduction efficiency of lentiviral vectors without the need of depleting hMARCH8. Our data thus provide one strategy to increase the transduction efficiency of lentiviral vectors.
The relatively low transduction efficiency of unstimulated T cells, B cells, and HSCs has limited the application of lentiviral vectors in gene therapy, and in the meantime inspired efforts to discover solutions. For example, efficient transduction of these important target cell types has been attempted by preparation of lentivirus of high purity, transduction at high MOIs, sequential transductions, and prolonged ex vivo culture. 38,79 –81 Other strategies include the use of chimeric glycoproteins fused to ligands, IgG, or single-chain variable fragments, and the use of adjuvants including polycations or cationic liposomes. 55 –57,82 –85 Alternatively, small molecules have been tested to overcome the intrinsic immune restriction factors that deter the transduction of lentiviral vectors. These include mTOR inhibitor rapamycin, the IFITM3-modulating cyclosporine, and caraphenol A, which have been shown to improve transduction through reducing the expression of antiviral effector IFITM3. 49,60 –62
However, some of these strategies raise safety concerns because they require treatment of the targeting cells, which are to be transfused back to the patients. Although whether it will help overcome the several cell-intrinsic barriers to transduction (such as IFITM3) need to be investigated, our study provides a new strategy to increase the transduction efficiency of lentiviral vectors through modifying viral envelope protein VSV G that is used to pseudotype lentiviral particles, thus avoiding manipulation of targeting cells. This strategy can be implemented using MARCH8-knockout cells to produce lentiviral particles, and/or using the MARCH8-resistant VSV G variant that was generated in this study. Since MARCH8 not only targets and degrades VSV G but also other viral envelopes that have been used in pseudotyping lentiviral particles or other viral gene delivery vectors, the MARCH8-knockout cells are recommended for producing different viral vectors of high titers. It is also worthwhile considering to develop other MARCH8-resistant viral envelopes that can be used to improve the efficiency of gene delivery.
Footnotes
Authors' Contributions
F.G., C.L., F.X., and X.L. conceived and designed the study. F.X., X.L., D.Z., F.Z., Z.F., S.H., S.M., Y.H., H.S., L.W., and L.G. performed the experiments. F.G., C.L., S.C., and J.W. discussed the results. F.G., C.L., and F.X. composed the article. All authors read and approved the final article.
Acknowledgments
We thank Dr. Zhaohui Qian for providing hMARCH8 DNAs and Dr. Conghui Wang for technical assistance in performing flow cytometry.
Author Disclosure
F.G., F.X., and X.L. are inventors on pending and issued patents on lentiviral vector technology and gene transfer. The other authors declare no competing interests.
Funding Information
This work was supported by funds from the National Key Plan for Scientific Research and Development of China (2018YFE0107600, 2016YFD0500307, and 2020YFA0707600), from the Ministry of Science and Technology of China (2018ZX10301408-003, 2018ZX10731101-001-018), from the National Natural Science Foundation of China (82072288, 81401673), from the CAMS Innovation Fund for Medical Sciences (CIFMS 2018-I2M-3-004, CIFMS 2020-I2M-2-014), from the Canadian Institutes of Health Research (CCI-132561), and from the CAMS general fund (2019-RC-HL-012).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.