
Abstract
Several neurodegenerative disorders are characterized by oligodendroglial pathology and myelin loss. Oligodendrogliopathies are a group of rare diseases for which there currently is no therapy. Gene delivery through viral vectors to oligodendrocytes is a potential strategy to deliver therapeutic molecules to oligodendrocytes for disease modification. However, targeting oligodendroglial cells in vivo is challenging due to their widespread distribution in white and gray matter. In this study, we aimed to address several of these difficulties by designing and testing different oligodendroglial targeting vectors in rat and mouse brain, utilizing different promoters, serotypes, and delivery routes. We found that different oligodendroglial promoters (myelin basic protein [MBP], cytomegalovirus-enhanced MBP, and myelin-associated glycoprotein [MAG]) vary considerably in their ability to drive oligodendroglial transgene expression and different viral vector serotypes (rAAV2/7, rAAV2/8, and rAAV2/9) exhibit varying efficacies in transducing oligodendrocytes. Different administration routes through intracerebral or intraventricular injection allow widespread targeting of mature oligodendrocytes. Delivery of rAAV2/9-MAG-GFP into the cerebrospinal fluid results in GFP expression along the entire rostrocaudal axis of the spinal cord. Collectively, these results show that oligodendrocytes can be targeted with high specificity and widespread expression, which will be useful for gene therapeutic interventions or disease modeling purposes.
INTRODUCTION
Oligodendroglial dysfunction and demyelination are apparent in leukodystrophies, multiple sclerosis, and several proteinopathies, such as multiple system atrophy (MSA) and progressive supranuclear palsy (PSP). 1 Leukodystrophies are juvenile monogenic diseases, where a defective gene leads to myelin pathology. 2 Proteinopathies are age-related protein-misfolding diseases where accumulation of protein leads to axonal loss and neuronal death, and in some cases oligodendroglial dysfunction and demyelination. 3 In MSA, the protein α-synuclein (αSyn) accumulates and forms inclusions, primarily in oligodendrocytes, 4 whereas in PSP, the tau protein forms insoluble inclusions in glial cells and causes oligodendroglial dysfunction. 5
Although no disease-modifying treatments are available for these diseases, clinical trials are underway for leukodystrophies using viral vectors or cell transplantation to deliver potentially therapeutic proteins to the brain. 5 Because these approaches do not use vectors that target oligodendrocytes, in both cases, for ex vivo or in vivo leukodystrophy gene therapy, the appropriate protein needs to be secreted from either microglia or neurons to compensate for the faulty gene in oligodendrocytes. These approaches have weaknesses when it comes to specificity and efficacy of the delivery of the protein and could potentially be improved if the gene of interest is targeted directly to oligodendrocytes. In the case of proteinopathies, clinical trials are testing if the administration of antisense oligonucleotides can lower the expression of pathogenic αSyn and tau, respectively, and counteract their toxic effects. 6,7 These therapies are again not specifically targeted to oligodendrocytes and could therefore still benefit from precision gene delivery strategies.
Recently, some efforts were already reported to selectively transduce oligodendrocytes with viral vectors in vivo. By using specific oligodendroglial promoters, such as the myelin basic protein (MBP) promoter or a novel chimeric oligodendroglial-specific serotype (olig001), it was possible to selectively target oligodendrocytes and promote protein expression. 8,9 Specifically, oligodendroglial expression of αSyn through viral vectors was sufficient to cause MSA-like neuropathology in vivo. 9,10 In addition, gene targeting of oligodendrocytes through the usage of a human CNP promoter in a self-complementary AAV, was used to suppress an oligodendrocyte-specific gene (Plp1), responsible for the leukodystrophies Pelizaeus/Merzbacher disease, resulting in an ameliorated pathology. 11 However, upscaling efforts would benefit from the availability of more efficient and specific viral vectors.
In this study, we produced and characterized viral vectors based on a combination of promoters, serotypes, and administration routes aiming to target oligodendrocytes with robust and widespread expression in rodent brain. More specifically, we compared different oligodendroglial-specific promoters, including MBP, myelin-associated glycoprotein (MAG), and a chimeric promoter carrying the early gene enhancer element of the cytomegalovirus (CMV) promoter fused to the MBP promoter (CMVieMBP). With the aim of achieving widespread transgene expression, we combined these expression constructs with different serotypes, rAAV2/7, rAAV2/8, and rAAV2/9, and administered the vectors through different routes, intrastriatally or intraventricularly (see Table 1 for a summary of all tested combinations).
Summary of different administration routes and vectors used in this study
CMVieMBP, cytomegalovirus promoter fused to the MBP promoter; MAG, myelin-associated glycoprotein; MBP, myelin basic protein.
We found that different oligodendroglial-expressing viral vectors have markedly different expression profiles. The expression pattern of oligodendroglial targeting vectors was also influenced by the administration route. Notably, when we administered the rAAV2/9-MAG GFP vector intracerebrally or intraventricularly, we achieved the most robust and widespread eGFP expression in oligodendrocytes throughout the rodent brain and spinal cord.
MATERIALS AND METHODS
Cloning of AAV plasmids
The plasmids for producing recombinant rAAV viral vector were generated as described previously. 12 These plasmids include the constructs for the different AAV serotypes 2/7, 2/8, and 2/9, the AAV transfer plasmid (pZac 2.1 eGFP3 SEED) encoding the eGFP transgene under the control of the oligodendroglial-specific promoters MBP, CMVieMBP, or MAG and the pAdvDeltaF6 adenoviral helper plasmid. The MBP promoter was generated as described previously 13 and the MAG promoter was created by PCR amplification of the MAG promoter in human HeLa cells (sense primer, ATATATGCTAGCCCTCAGAAGGAACCAACACTGCCAGCACTT and antisense primer, ATTTATGATATCGCCCCCACTTGCCAGCCCCTCCCCTCCC). Initially, the eGFP transgene was replaced by a multiple cloning site inserted between the NotI and BamHI sites in the original transfer plasmid. The eGFP was inserted into this multiple cloning sites. This construct will further be referred to as the transfer plasmid. The CMVie enhancer was generated as described previously 14 and cloned into the transfer plasmid.
Serum-free recombinant rAAV production
Subconfluent, low (o50) passage-adherent HEK 293T (293T) cells (ATCC, Manassas, VA) were transfected using a 25-kDa linear polyethylenimine solution using the AAV-TF, AAVrep/cap, and pAdbDeltaF6 plasmids in the ratio of 1:1:1. Productions were performed using a Hyperflask Cell Culture Vessel (1,720 cm2 growth area, Corning Life Sciences, Kennebunk, ME). The 293T cells were seeded in the Hyperflask at 108 cells per production in Opti-MEM (Invitrogen, Merelbeke, Belgium) with 2% fetal calf serum. The next day, 400 μg of pAAVTF plasmid, 400 μg of rep/cap construct, and 400 μg of pAdbDeltaF6 plasmid were mixed in 20 mL of 150 mM NaCl. An equal volume containing 4 mL of 10 mM polyethylenimine solution and 16 mL of 150 mM NaCl was added slowly to the DNA mixture. Following 15 min incubation at room temperature, the DNA–polyethylenimine complex was added to the 293T cells in Opti-MEM with 0% fetal calf serum. After 24 h of incubation at 37°C in a 5% CO2-humidified atmosphere, the medium was replaced with Opti-MEM with 0% fetal calf serum. The supernatant was harvested 3 days after transient transfection and concentrated using tangential flow filtration.
Recombinant rAAV purification
The concentrated supernatant was purified using an iodixanol step gradient. Iodixanol was diluted with phosphate-buffered saline (PBS) containing a final concentration of 1M NaCl in 20, 30, and 40% (w/v) solutions. A discontinuous gradient was made by carefully underlayering the concentrated supernatant with 5 mL of 20% iodixanol, 3 mL of 30% iodixanol, 3 mL of 40% iodixanol, and 3 mL of 60% iodixanol. The gradient was centrifuged in a Beckman Ti-70 fixed angle rotor (Analis, Gent, Belgium) at 27,000 r.p.m. for 2 h. Gradient fractions were collected in 250 mL aliquots and all fractions with a Refraction Index between 1.39 and 1.42 were pooled. The pooled fractions were centrifuged in a Vivaspin 6 (PES, 100,000 MWCO, Sartorius AG, Goettingen, Germany) using a swinging bucket rotor at 3,000 g. The iodixanol of the pooled fractions was exchanged five times with PBS using this procedure. The final sample was aliquoted and stored at −80°C. Characterization of the rAAV stocks included real-time PCR analysis for genomic copy determination and silver-stained sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis for vector purity. Briefly, real-time PCR was performed using a primer probe set for the polyA sequence (primer sequences, 5′-TCTAGTTGCCAGCCATCTGTTGT-3′ and 5′-TGGGAGTGGCACCTTC CA-3′; probe sequence, 5′-TCCCCCGTGCCTTCCTTGACC-3′). Genome copies were adjusted to ∼3 × 1012 GC/mL for all viral vector productions.
Stereotactic injections
All animal experiments were carried out in accordance with the European Communities Council Directive of November 24, 1986 (86/609/EEC) and approved by the Bioethics Committee of the KU Leuven (Belgium). The KU Leuven Institutional Review Board approved animal experiments (project number P070/2017). Young adult female Sprague–Dawley rats (8 weeks old, n = 4/group), (Janvier, Le-Genest-Saint-Isle, France), weighing about 200–250 g, were housed under a normal 12-h light/12-h dark cycle with free access to pelleted food and tap water. All surgical procedures were performed using aseptic techniques and ketamine (60 mg/kg i.p. Ketalar, Pfizer, Puurs, Belgium) and medetomidine (0.4 mg/kg and Dormitor, Pfizer, Belgium) anesthesia. Following anesthesia, the rodents were placed in a stereotactic head frame (Stoelting, Wood Dale, IL). Intrastriatal injections were performed with a 30-gauge needle and a 10-μL Hamilton syringe. All animals were injected unilaterally with 4 μL of vector divided over two locations, 1.5 μL according to the stereotactic coordinates anteroposterior (AP): +0.07 cm, lateral (LAT): −0.28 cm, and dorsoventral (DV): −0.64 cm; and 2.5 μL AP: +0.07 cm, LAT: −0.28 cm, and DV: −0.49 cm for a total of 1.2 × 1010 GC. The injection rate was 0.25 μL/min; the needle was left in place for an additional 5 min before being retracted. For cerebellar injections all animals were injected bilaterally with 1.5 μL vector in the left and right middle cerebellar peduncle by using the following coordinates: AP): −/+0.34 cm, LAT: −0.34 cm, and DV: −0.65 cm for a total of 9 × 109 GC. The injection rate was 0. 25 μL; the needle was left in place for an additional 5 min before being retracted.
Intracebroventricular injections
Newborn pups (n = 3/group) were injected intracerebroventricularly (ICV) as close as to their birth at P0 (6–12 h), P1/2 (48–60 h), or P3 (72–84 h). Naive pups were isolated from their mother when milk spots were visible and were kept warm on a heating pad while awaiting surgery. Only half of the pups were taken from their mother during nursing. Before surgery, pups were cryoanesthetized on a metal plate on ice to cool down body temperature to 4°C and subsequently transferred to a cooled neonatal surgery frame that was maintained between 2°C and 6°C with dry ice. Hypothermia anesthesia was observed by a change of body color from pink to purple and a complete stop of body movement and respiration. The pup head was wiped with 70% ethanol, leveled parallel with the injection frame, and fixed gently between two ear bars. Cranial blood vessels were used to determine the lambda sutures for ICV injection coordinates. A 34G 10 μL Hamilton needle was placed at +0.165 cm A/P, ±0.080 cm M/L from lambda. The bevel of the needle was used to pierce the skull and positioned just below the flattened skull, after which it was lowered to −0.170 D/V and retracted to −0.130 D/V for ventricular perfusion. Injection was performed bilaterally for a volume of 2 μL and a total volume of 4 μL at a perfusion rate of 0.5 μL/min for a total of 1.2 × 1010 GC. After the injection, the skull was flattened by gently retracting the needle to open the ventricles and the needle was left for another 2 min to allow the injected bolus to spread throughout the ventricles. The needle was slowly retracted over 1 min. Viral vector was supplemented with 0.5% Trypan Blue dye to achieve a 0.05% Trypan Blue solution to assess successful ventricular injection. After injection, pups were allowed to recover on a 37°C heating pad and returned to their home cage. The procedures as described above for IVC surgery were approved by the Van Andel Institute Institutional Animal Care and Use Committee (IACUC).
Adult animals (described in Supplementary Fig. S1) were ICV at 8–12 weeks of age. Animals were anesthetized with a cocktail solution of ketamine (60 mg/kg i.p. Ketalar, Pfizer, Puurs, Belgium) and medetomidine (0.4 mg/kg and Dormitor, Pfizer, Belgium) and positioned in a stereotaxic head frame (Stoelting, Wood Dale, IL). Viral vectors were administered by stereotaxic surgery into the right lateral cerebral ventricle with a Hamilton 34G needle, in the following coordinates from Bregma: anteroposterior = −0.2 mm; mediolateral = −1.0 mm; dorsoventral = −2.3 mm. With the aid of a pump, 2.2 × 1010 viral vector particles (7.5 μL) were injected at a rate of 0.5 μL/min during 15 min. The needle was left for additional 5 min to allow ventricle spreading of the vector. The needle was slowly retracted over 3 min. Animals were sutured and placed in a heating pad until recovery and returned to their home cage.
Histology
Two weeks after rAAV vector injection, rats were euthanized with an overdose of sodium pentobarbital (60 mg/kg, i.p., Nembutal, Ceva Sante, Belgium) followed by intracardial perfusion with 4% paraformaldehyde in PBS. After fixation overnight, 50-mm-thick coronal brain sections were made with a vibrating microtome (HM 650V, Microm, Walldorf, Germany). IVC-injected mice were euthanized after 20 days followed by intracardial perfusion with 4% paraformaldehyde in PBS. Immunohistochemistry was performed on free-floating sections using an in-house antibody raised against eGFP (rabbit polyclonal 1:10,000). Sections were pretreated with 3% hydrogen peroxide for 10 min and incubated overnight with primary antibody in 10% normal goat serum (DakoCytomation, Heverlee, Belgium). As a secondary antibody, we used biotinylated anti-rabbit IgG (1:300, DakoCytomation), followed by incubation with a streptavidin–horseradish peroxidase complex (DakoCytomation). eGFP immunoreactivity was visualized using Vector SG hydrogen peroxidase substrate kit (SK-4700; Vector Laboratories, CA, USA) as a chromogen. For fluorescent double staining, sections were rinsed thrice in PBS and then incubated overnight in PBS–0.1% Triton X-100, 10% donkey serum, chicken anti-eGFP (1:1,000, 1020; Aves Laboratories, OR, USA) in combination with rabbit anti-NeuN (1:1,000, ABN78; Sigma-Aldrich, MO, USA), rabbit anti-GST-π (1:1,000, 311; MBL Life, MA, USA), goat anti-Iba-1 (1:1,000, ab107159; Abcam, Cambridge, United Kingdom), or rabbit anti-S100β (1:1,000, ab41548; Abcam). After three rinses in PBS–0.1% Triton X-100, the sections were incubated in the dark for 1 h in fluorochrome-conjugated secondary antibodies: goat anti-rabbit Alexa 488 (1:500, Molecular Probes; Invitrogen) and goat anti-mouse Alexa (1:500, Molecular Probes; Invitrogen). After being rinsed in PBS and mounted, the sections were coverslipped with Mowiol mounting medium (Calbiochem®, San Diego).
Quantification of transgene expression
The volume of eGFP-immunoreactive transduced cells in the brain was determined by stereological volume measurements using the Cavalieri method in a computerized system as previously described 15 (StereoInvestigator; MBF Bioscience, Magdeburg, Germany) in a total of 10 sections/animal. The number of positive cells, as well as the type of cells obtained, was measured in a section adjacent to the injection site and divided by that area, in which condition the coefficients of error, calculated according to the procedure of Schmitz and Hof as estimates of precision 16 varied between 0.05 and 0.14.
Quantification of GFP-positive cells for relative GFP expression in oligodendrocytes was performed through the MBF stereologer (StereoInvestigator; MBF Bioscience) and MBF software (MBF Bioscience). The rat striatum was equally divided into seven fields (20 × magnification) along a mediolateral axis and the average GFP expression per cell was quantified separately for each field. Within each field of view, individual GFP-positive cells were outlined and selected as a region of interest. From each cell, the total fluorescence was determined. After collecting the fluorescent values of every cell within a field of view, the average fluorescence intensity per cell was made. Presented values are from four different animals and all values were normalized against the lowest value measured between experimental conditions. The lowest measured fluorescent value was set to 1. Results are plotted as relative GFP expression, or fold increase versus the lowest measured value.
For fluorescent analysis of GFP expression in spinal cord segments, the total fluorescence intensity per slide was determined after correcting for background with sliding paraboloid background subtraction in ImageJ.
Statistical analysis
For each serotype, the mean expression volume and the mean expression efficiency were compared using one-way analysis of variance (ANOVA) followed by Bonferroni post hoc testing. To compare the total number of positive cells, we used one-way ANOVA followed by Tukey's post hoc test. Statistical analysis was performed using GraphPad Prism. Statistical significance level was set as follows: *p < 0.05, **p < 0.01, and ***p < 0.001.
RESULTS
Oligodendroglial promoters in rAAV capsids drive specific transgene expression in oligodendrocytes in vivo
To achieve specific expression in oligodendrocytes in rat brain, we compared three promoters specific for mature oligodendrocytes (Table 1). Since it was previously shown that adenoassociated viral vectors of the 2/8 serotype (rAAV2/8) can transduce oligodendrocytes in vivo, 1 we designed rAAV2/8 viral vectors in combination with the oligodendrocyte-specific promoters of the MBP and the MAG. In addition, we tested if we could enhance transgene expression by adding the immediate early gene enhancer element of the CMV promoter to the MBP promoter, thereby generating a hybrid construct, which we termed CMVieMBP (Fig. 1A).
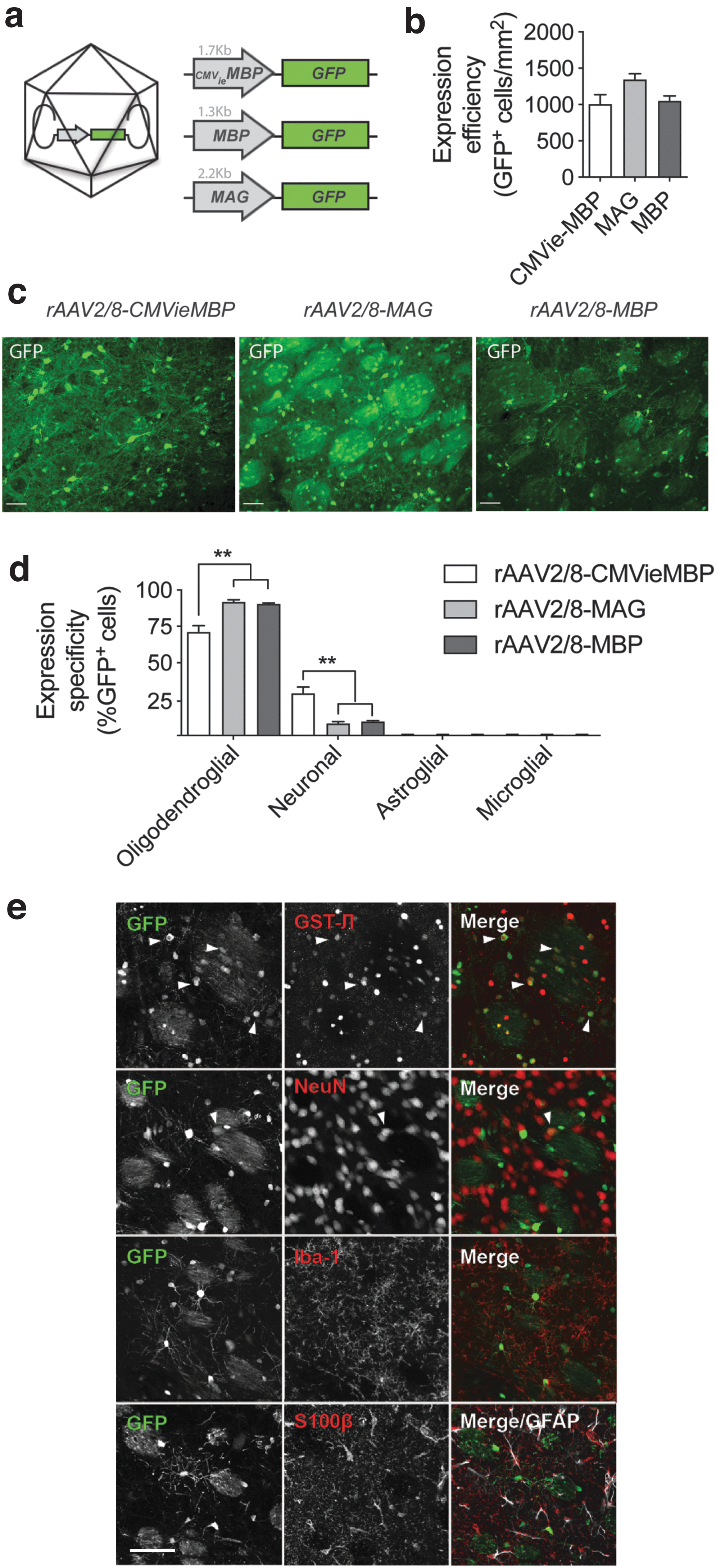
rAAV2/8-mediated expression of GFP under control of different oligodendroglial promoters yields variable expression patterns.
For characterization purposes, we injected 8-week-old rats, separately, with rAAV2/8-MBP, rAAV2/8-MAG, or rAAV2/8-CMVieMBP viral vectors, all of which express the GFP reporter gene. We chose to inject into the striatum, where the viral vectors can target mature oligodendrocytes in the fibers of the internal capsule that traverse the striatum, as well as in the corpus callosum that is immediately adjacent to the striatum. Two weeks after injection, we analyzed GFP expression in different cell populations by dual immunostaining for GFP and cell type-specific markers.
We evaluated both expression specificity and staining intensity (Fig. 1C). Expression efficiency calculated at the relative number of GFP-positive cells was similar for the three promoter constructs (Fig. 1B). Through double immunostaining, we observed that in all conditions GFP was expressed in oligodendrocytes (Fig. 1D, E), although with varying specificities. Both the MBP and MAG promoters drove expression in oligodendrocytes with a specificity of ∼90% in rat striatum (91.9% for the MBP promoter and 90.4% for the MAG promoter, n = 4 with standard error of the mean [SEM]). In contrast, the CMVieMBP induced less specific expression in oligodendrocytes with 71.1% of GFP-positive cells colabeling with the oligodendroglial marker GST-π. The remaining fraction of GFP-positive cells was identified as NeuN positive, indicating low expression in neurons. We did not find GFP expression in astrocytes or microglial cells (Fig. 1E, lower panels).
To determine the relative transgene expression levels induced by the different promoters, we analyzed GFP fluorescence in individual oligodendroglial cells in the striatum along a mediolateral axis perpendicular to the injection tract. The obtained heat maps indicate that the hybrid promoter CMVieMBP expressed relatively higher levels of GFP and over a larger brain area followed by the MAG and MBP promoters (Fig. 2A).

Relative transgene expression levels in oligodendrocytes using different cell-type-specific promoters.
Transgene expression efficiency and pattern is further determined by viral vector serotype
Given the greater specificity of oligodendroglial expression of the MAG promoter compared with the hybrid promoter CMVieMBP and its relatively higher expression efficiency compared with the MBP promoter (Figs. 1C and 2), we selected the MAG promoter for subsequent experiments, where we wanted to compare the effects of viral vector capsids on oligodendroglial transduction. Viral vector serotypes can influence transduction efficiency through the presence of different capsid receptors and their differential binding to various cell types. 17,18 In an attempt to further improve the expression capacity of our MAG-driven viral construct, we tested if substituting the viral vector serotype could further reinforce transgene expression in oligodendrocytes and its expression volume. We focused on the three following serotypes, rAAV2/7, rAAV2/8, and rAAV2/9 (Fig. 3A), since we previously demonstrated that they yield widespread transgene expression in rodent brain. 14
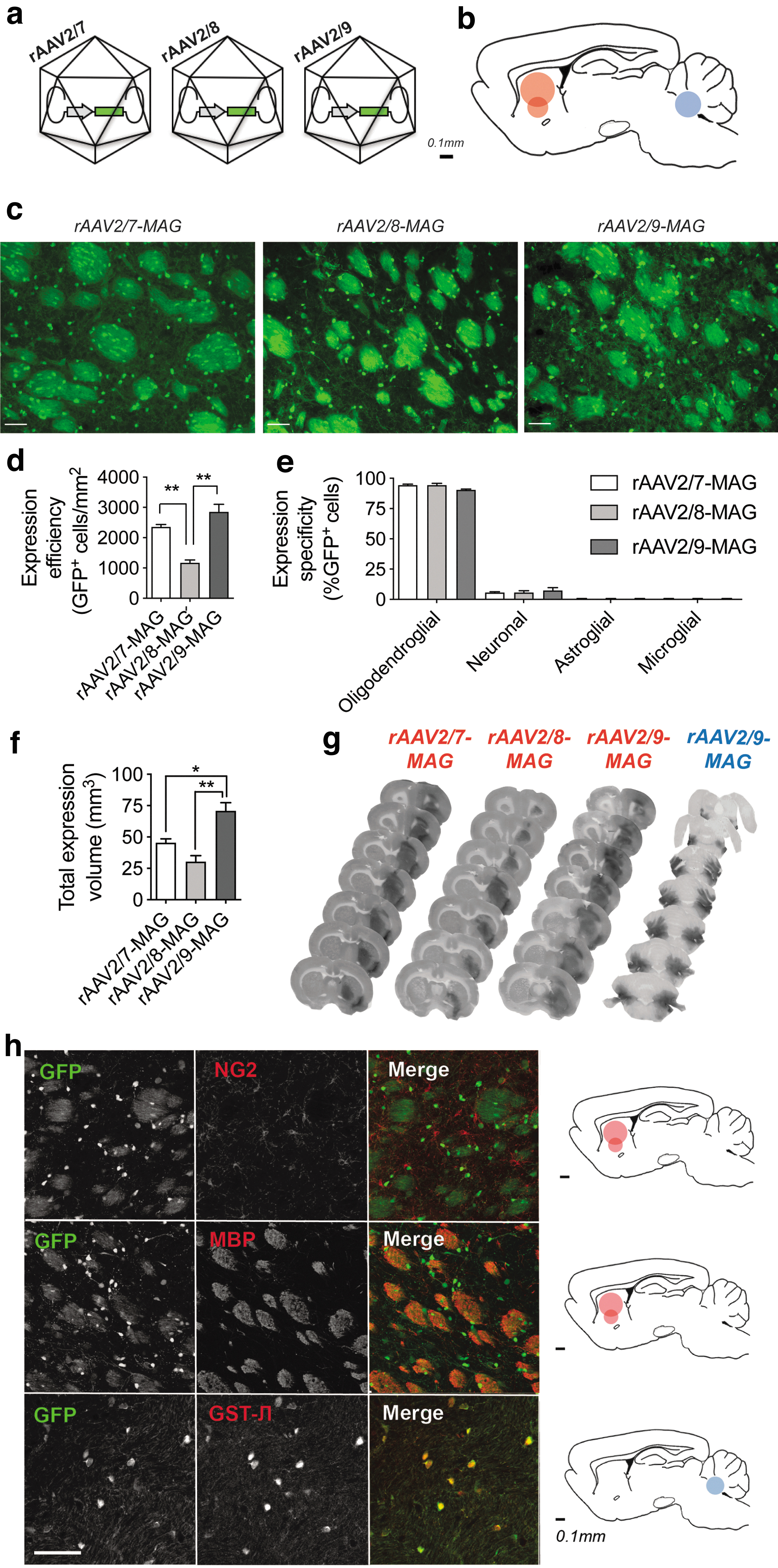
Expression pattern and specificity in oligodendrocytes using different viral vector serotypes.
We injected the viral vectors into rat striatum and allowed the expression for 2 weeks, after which the animals were analyzed (Fig. 3B). We first confirmed that the serotypes, rAAV2/7 and rAAV2/9, did not alter expression specificity compared with rAAV2/8 (Fig. 3C). To this aim, we characterized GFP expression through GFP fluorescence in oligodendrocytes and found that substituting viral serotypes did not change transgene expression specificity (Fig. 3E). Interestingly, both rAAV2/7 and rAAV2/9 yielded 50% and 59% more GFP-positive cells in rat striatum compared with rAAV2/8 (one-way ANOVA, **p = 0.0013, n = 4 with SEM) (Fig. 3D).
When examining the whole brain, we observed that rAAV2/9-MAG-mediated oligodendroglial GFP expression was superior to rAAV2/7 and rAAV2/8 and extended well beyond the corpus callosum into cortical areas with an average expression volume of 70.7 mm3 (Fig. 3F, G). Next, we further characterized expression specificity of rAAV2/9-MAG within different brain regions and in terms of oligodendrocyte cell types. Double staining for the marker NG2, which labels oligodendroglial precursor cells (OPCs), revealed no overlap between GFP-expressing cells and OPCs (Fig. 3H). In contrast, the MBP marker revealed extensive overlap with GFP in rat striatum, indicating expression in mature oligodendrocytes (Fig. 3H). When the rAAV2/9-MAG-GFP viral vectors were injected in the middle cerebellar peduncle of the cerebellum, another region rich in oligodendrocytes, we also exclusively observed oligodendroglial expression (Fig. 3G, H).
Lastly, we asked if different serotypes induced different expression distribution within oligodendrocytes. By measuring GFP fluorescence along a lateromedial axis, perpendicular to the injection site, we did not observe any significant differences between serotypes (Fig. 4A, B). However, rAAV2/7-MAG-GFP showed a gradual loss of expression with increasing distance from the injection site. This was in contrast to the rAAV2/9-MAG-GFP that showed a more even expression pattern from the point of injection toward more lateral areas.
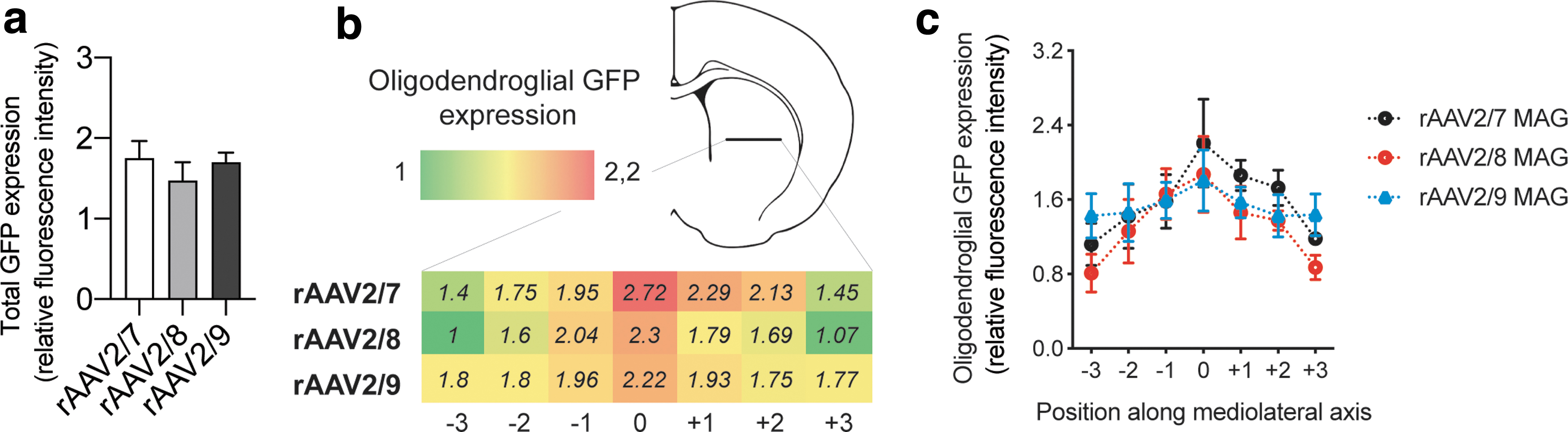
Relative transgene expression levels in mature oligodendrocytes using different viral vector serotypes.
Intracerebroventricular delivery in neonates results in widespread expression in rodent spinal cord
Widespread viral vector-based gene therapy in the central nervous system (CNS) is hampered by a lack of vector distribution after intracerebral administration. Intrathecal or intraventricular delivery are therefore sometimes used to achieve widespread expression throughout the CNS. It has been previously shown that the rAAV2/9 serotype can efficiently spread through the cerebrospinal fluid and diffuse into the brain parenchyma, thereby transducing neuron and astroglia after intraventricular delivery to neonates. 19 Here, we asked if we could achieve specific oligodendroglial transgene expression using the rAAV2/9-MAG viral vector under a similar paradigm.
We bilaterally injected 2 μL of rAAV2/9 MAG GFP (3 × 1012 GC/mL) into the left and the right lateral ventricles of neonatal mice. During the first 24 h after birth, the ependymal lining of the ventricles is still immature and closes several days after birth. By injecting at ∼24, 48, and 72 h after birth (P0, P1, and P2), we can further assess how ventricle maturation affects vector diffusion throughout the CNS. We allowed all animals to age for an additional 20 days postinjection, after which the cerebrum and spinal cord were analyzed.
In the cerebrum, GFP expression was detected in the cortex and was mostly apparent along the needle tract (Fig. 5A). This might be due to minor damage along the needle tract that facilitates diffusion of the vector within these cortical areas. We further observed transgene expression in areas adjacent to the lateral ventricles and the subarachnoid space, which include the hippocampus, olfactory bulb, pons, and throughout the entire length of the spinal cord white matter (Fig. 5A, B).
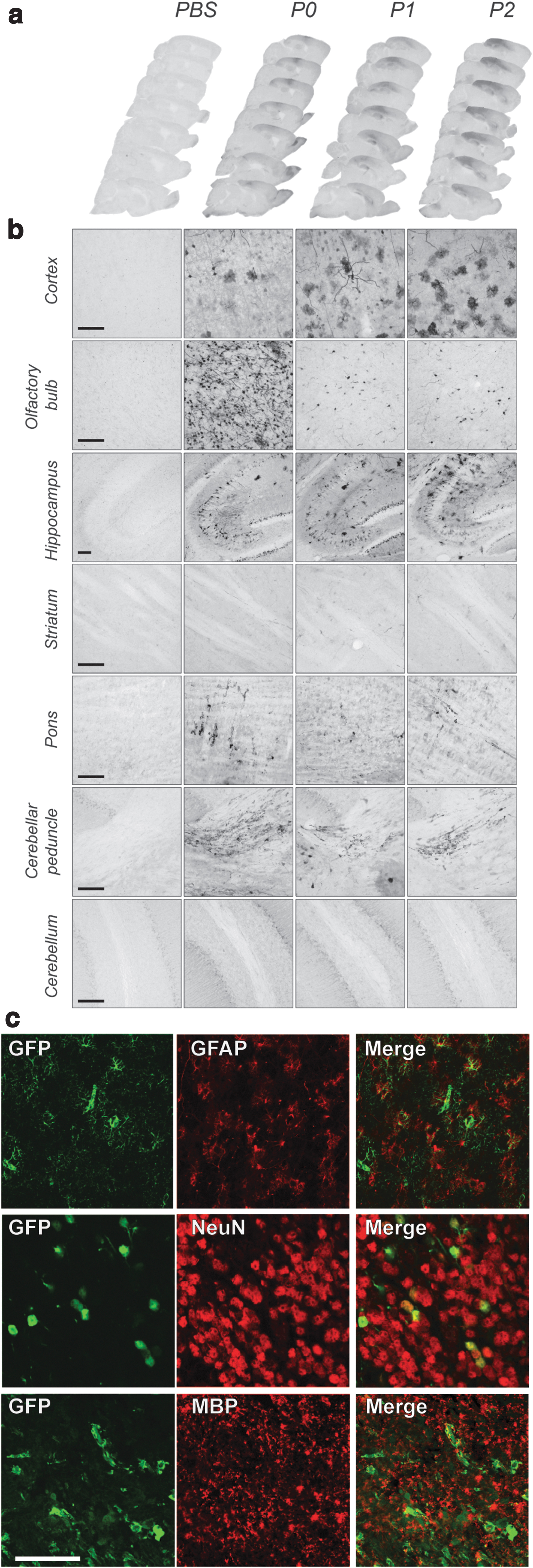
Intracerebroventricular delivery of rAAV2/9-MAG-GFP in mouse neonates results in oligodendroglial expression in the brainstem but lacks expression in the striatum or cerebellum. rAAV2/9-MAG-GFP viral vector was delivered intracerebroventricularly in mouse neonates 12 h (P0), 36 h (P1/2), or 60 h (P3) after birth.
Interestingly, the expression in cortical areas and hippocampus varied depending on the age of the neonate at the time of injection. In animals injected at P0, GFP expression was present in both neuronal and glial cells. However, in animals injected at later time points, we observed expression only in glial cells (Fig. 5B). In the olfactory bulb, expression was restricted to neurons in the P0 group and was less pronounced at P1 and P2 stages (Fig. 5B, C).
In the pons and spinal cord, we detected GFP exclusively in oligodendrocytes, as revealed in sections double labeled with GST-π and MBP markers (Fig. 5C). Transgene expression in spinal cord was distributed equally in myelinated regions of the cervical, thoracic, and lumbar regions and was primarily restricted to the outer ventral, lateral, and dorsal white matter columns (Fig. 6A, B). The expression was lower more caudal toward the sacral segments (Fig. 6B). Transgene expression was more apparent in the spinocerebellar and spino-olivary tracts where GFP was found in GST-π-positive cell bodies and myelinating fibers (Figs. 5C and 6B). We did not detect GFP expression in other cell types of the spinal cord white matter tracts. Adjusting viral titers from 1 × 1012 GC/mL to 3 × 1012 GC/mL resulted in a similar expression pattern but with two times higher expression levels measured through GFP fluorescence (Fig. 6C).
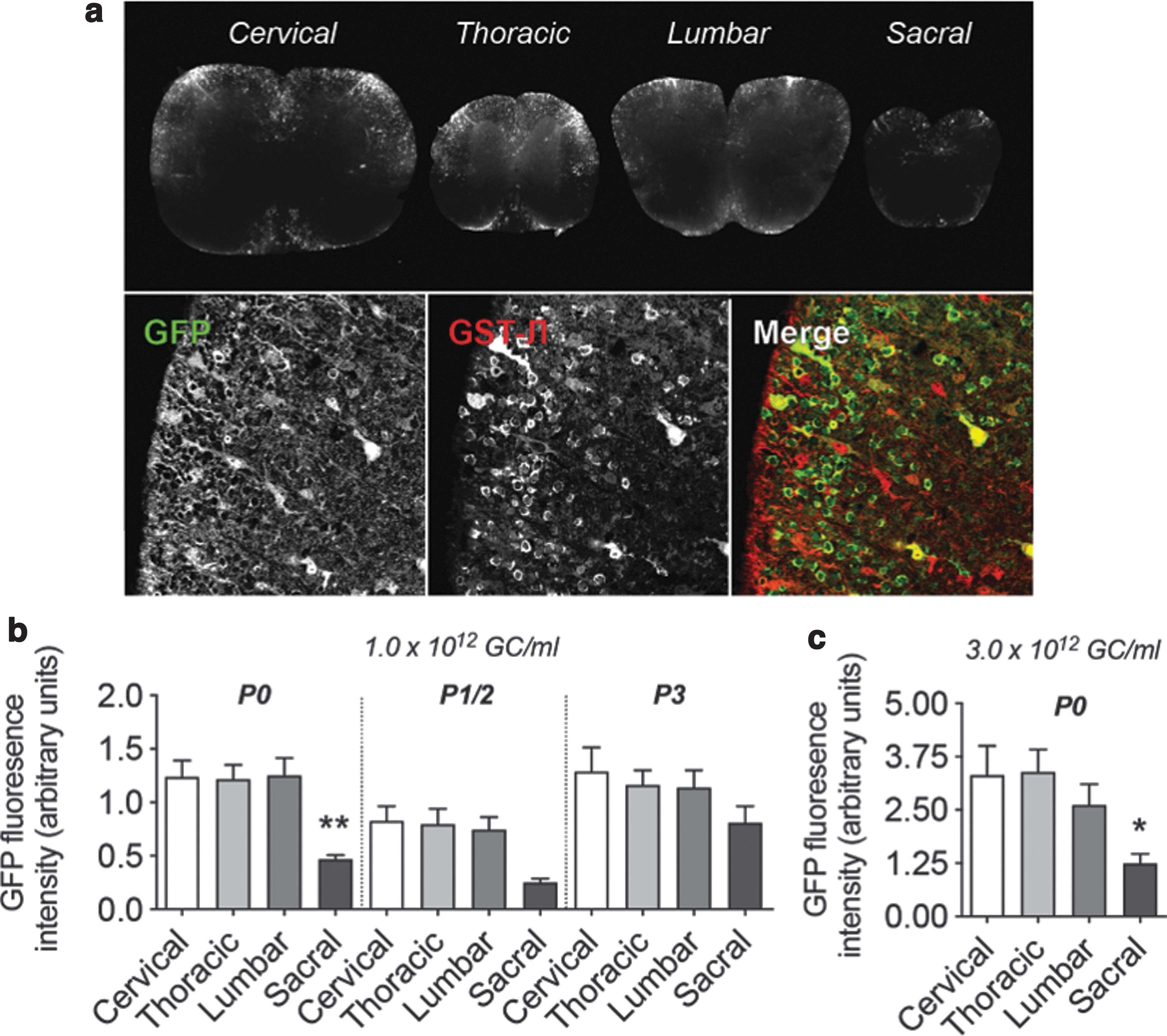
Intracerebroventricular delivery of rAAV2/9-MAG-GFP in mouse neonates results in oligodendroglial transgene expression in the spinal cord. rAAV2/9-MAG-GFP viral vector was delivered intracerebroventricularly in mouse neonates 12 h (P0), 36 h (P1/2), or 60 h (P3) after birth.
DISCUSSION
Gene delivery to oligodendrocytes might be a valuable and promising method for treatment of a number of diseases, such as MSA or some leukodystrophies. Therapies aimed at targeting oligodendrocytes instead of neurons in the CNS have rarely been explored. Recombinant AAV vectors have become a preferred method of gene delivery to the nervous system due to their low immunogenicity, tissue tropism, nonintegrating properties, long-term expression, and the possibility to administer systemically, centrally, or locally. 20 Targeting oligodendrocytes through viral vectors has proven difficult due to large promoter constructs that restrict the packaging capacity of viral vectors. In addition, the widespread pathology or systemic nature of oligodendrogliopathies ideally requires delivery and transduction of viral vectors throughout the whole CNS. In this study, we aimed to address several of these difficulties by designing and testing different oligodendroglial targeting vectors in vivo, utilizing different promoters, serotypes, and delivery routes.
An important consideration when designing packaging constructs for AAV vectors is their relatively small size of ∼5 kb. 21 Constructs any longer than this will inevitably result in lower titer yields, transgene recombination, or transgene truncation. The MBP and MAG promoters are 1.3 and 2 kb in size, respectively, leaving ∼3 kb for packaging transgenes into the cassette, one strategy to create more packaging capacity would be to truncate the promoter and use smaller versions of the MBP or MAG promoters to drive transgene expression. A previous study 22 examined this possibility by truncating the MAG promoter. It was shown that the truncated forms of 1.3 and 0.3 kb still expressed in oligodendrocytes but demonstrated a gradual loss of expression specificity.
In an attempt to further enhance transgene expression with the 1.3 kb MBP promoter, we also examined the usefulness of enhancer elements. The enhancer element of the CMV promoter is known to stimulate expression in human cells and cells of other species. 23 By combining the CMV immediate early enhancer (CMVie) in conjunction with the MBP promoter, we found that CMVie did indeed increase expression levels. However, it also reduced expression specificity and promoted transgene expression in neurons. Hence, promoter design and size could be chosen based on the cellular specificity and expression levels required for a specific transgene.
Next to promoter selectivity, capsid design can further modify transduction efficiency. 24 Viral vector serotypes strongly affect gene delivery in vivo, although this has not been tested systematically for oligodendrocytes. Different serotypes, such as the rAAV2/1 and multiple nonprimate nonchimeric serotypes, 9,22 have been shown to transduce oligodendrocytes in vivo, although to varying efficiencies. Recently, Powell et al. generated a chimeric capsid through capsid shuffling and directed evolution. 8 This capsid, which was termed Olig001, binds to oligodendrocytes with a much higher affinity compared with rAAV2/8 and transduces oligodendrocytes with over 90% specificity.
This allowed the authors to use constitutive promoters. 8 This strategy differs from our study, since we have used capsids allowing transducing both neurons and glial cells, but achieving oligodendroglial expression specificity by means of the promoter. A subsequent study using the Olig001 serotype reported an expression volume of 4 mm3 after intrastriatal injection in rat brain, which is considerably lower than what we observed in our study in rat brain (70 mm3). 10 Compared with other serotypes, the rAAV2/9-MAG viral vector showed a markedly efficient expression profile with high cell-type specificity and a large expression volume. Other capsids yielded significantly lower expression volumes, but with similar specificities.
The transduction profile of rAAV2/9 capsid is unique because of its galactose and laminin-binding receptors, 18 through which it is able to bind with multiple cell types and diffuse efficiently throughout the CNS. 25 It has furthermore been shown to cross the blood/brain barrier and enter the brain after intravenous administration. 26 Because of its transduction properties and translational purposes, we tested if the rAAV2/9-MAG oligodendroglial-expressing vector could drive gene expression in the CNS after delivery to the CSF (cerebrospinal fluid). We injected neonatal pups at different ages (P0, P1, and P2) and assessed oligodendroglial GFP expression when the MAG promoter is active for the production of myelin in juvenile mice (at P20). Interestingly, in the cerebrum, we mostly observed expression in nonoligodendroglial and sparse oligodendroglial-positive cells in the pons. However, over the entire rostrocaudal axis of the spinal cord, we detected expression in the white matter tracts of the anterior and dorsal spinal columns.
During early developmental stages, the ependymal lining of the lateral ventricles matures and restricts the flow of CSF to the brain parenchyma. Similar to our observations, it has been shown that during closure of the ventricles, rAAV2/9 vector expression shifts from neuronal to astroglial cells in cortical areas of young and adult rodents. 27 –29 Nevertheless, when delivering rAAV2/9 into the CSF through ICV (intracerebroventricular) injection, we did not observe any significant restriction of vector diffusion from the CSF to the spinal cord white matter. The expression of MAG during development is reported to increase after 12 days (Pedraza et al.) 30 and reach maximal expression at 1 month, which remains stable thereafter. Although at the time of neonatal intraventricular injection, the promoter activity is low (at P0–P2), animals, however, were analyzed at P20, leaving sufficient time for the transgene to become expressed. At this time point, we observe transgene GFP expression in oligodendrocytes of the spinal cord white matter tracts and also in the midbrain. This indicates that after intraventricular injection, and at the time points tested, the MAG promoter is active.
We further tested if the weaker expression seen after neonatal ICV administration might be due to developmental differences. We injected rAAV2/9 MAG GFP in the lateral ventricles of 8-week-old adult mice. With similar numbers of injected particles as used for the intrastriatal route, we again observe low overall GFP expression (Supplementary Fig. S1). Although we cannot strictly exclude that developmental changes in promoter activity might still affect protein expression levels, these experiments collectively suggest that dilution of the vector in the ventricles rather than weak promoter activity during development might be the reason of the relatively low transgene expression levels after ICV injection.
The highest transgene expression after ICV injection was observed in the ventral spino-olivary and spinocerebellar tracts. The pons, spino-olivary, and spinocerebellar tracts are among the first regions to be affected in the cerebellar variant of MSA (MSA-C). 31 The protein αSyn can accumulate in oligodendrocytes (to a lesser extent in neurons), and form deposits that trigger oligodendrogliopathy followed by axonal and neurodegeneration. 32 Targeting SNCA mRNA with these newly developed vectors to knock down αSyn levels in oligodendrocytes in CNS could be further explored as a potential useful strategy to slow disease progression in MSA. 33
CONCLUSION
In conclusion, by optimizing viral vector design we show that we can achieve efficient targeting of oligodendrocytes in vivo. We used different promoters that were specific for oligodendroglial expression. Choosing different viral serotypes and administration routes allowed us to further fine-tune gene delivery to different CNS areas. The viral vectors we used represent useful tools for delivery of therapeutic genes, gene silencing, or other tools for disease modeling and disease modification in animals. Finally, we propose that targeted gene delivery through viral vectors could lay the foundation for novel therapeutic approaches in oligodendrogliopathies.
Footnotes
AUTHORS' CONTRIBUTIONS
Conceptualization: W.P. and F.B.; Methodology: W.P, F.B., and C.V.H.; Investigation: W.P., F.B., and S.J.A.; Writing—Original Draft: W.P.; Writing—Review and Editing: W.P., F.B., J.A.S., P.B., and V.B.; Funding Acquisition: V.B.; Supervision P.B. and V.B.
AUTHOR DISCLOSURE
P.B. has received commercial support as a consultant from Axial Biotherapeutics, Calico, CuraSen, Fujifilm-Cellular Dynamics International, Idorsia, IOS Press Partners, LifeSci Capital LLC, Lundbeck A/S, and Living Cell Technologies LTD. He has received commercial support for grants/research from Lundbeck A/S and Roche. He has ownership interests in Acousort AB and Axial Biotherapeutics and is on the steering committee of the NILO-PD trial. The other authors declare no additional competing financial interests.
FUNDING INFORMATION
This work was supported by the FWO Flanders (Projects G.0927.14, G080517N, PhD fellowship to FB and postdoctoral fellowship to WP), Fulbright Program (fellowship to WP), the KU Leuven (C14/18/102), the ERA-NET JPco-fuND 2015 SYNACTION, the Medical Foundation Queen Elisabeth, the Vlaamse Parkinson Liga, and the Fund Sophia managed by the King Baudouin Foundation. J.A.S. and P.B. were supported by Van Andel Institute.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.