
Abstract
Infants and older adults are especially vulnerable to infection by respiratory syncytial virus (RSV), which can cause significant illness and irreparable damage to the lower respiratory tract and for which an effective vaccine is not readily available. Palivizumab, a recombinant monoclonal antibody (mAb), is an approved therapeutic for RSV infection for use in high-risk infants only. Due to several logistical issues, including cost of goods and scale-up limitations, palivizumab is not approved for other populations that are vulnerable to severe RSV infections, such as older adults. In this study, we demonstrate that intranasal delivery of adeno-associated virus serotype 9 (AAV9) vector expressing palivizumab or motavizumab, a second-generation version of palivizumab, significantly reduced the viral load in the lungs of the BALB/c mouse model of RSV infection. Notably, we demonstrate that AAV9 vector-mediated prophylaxis against RSV was effective despite the presence of serum-circulating neutralizing AAV9 antibodies. These findings substantiate the feasibility of repeatedly administering AAV9 vector to the airway for seasonal prophylaxis against RSV, thereby expanding the application of vectored delivery of mAbs as an effective prophylaxis strategy against various airborne viruses.
INTRODUCTION
Respiratory syncytial virus (RSV) can cause both upper and lower respiratory tract infections. 1 Infants are especially vulnerable to RSV in their first 2 years of life. 1 In addition, older adults and patients with chronic lung or heart diseases or with immune deficiencies are highly susceptible to severe clinical sequelae following infection with RSV. 1 While most affected patients present with various disease symptoms, ∼2% of affected patients require hospitalization for symptom management. 2 Currently, intramuscular injection of the monoclonal antibody (mAb) palivizumab, a Food and Drug Administration (FDA)-approved treatment for high-risk infants, confers some level of protection against the virus during the RSV season. 1 –3 Palivizumab targets a highly conserved region of the RSV fusion glycoprotein (F0) and has demonstrated in vivo efficacy against both the RSV A and B strains. 2,3 However, palivizumab is not approved for use in adults. 4
The second-generation version of palivizumab is an anti-RSV glycoprotein F recombinant mAb known as motavizumab. Medimmune (acquired by AstraZeneca) developed motavizumab by affinity maturation, with this modification resulting in a significantly higher (∼70-fold) binding affinity for RSV. 2,3 In passive transfer studies conducted in the cotton rat model, motavizumab was highly efficient and resulted in a 100-fold greater neutralization of RSV compared with palivizumab. 2,3
In a Phase III noninferiority trial, subjects treated with motavizumab experienced 50% fewer RSV medically attended lower respiratory tract infections than subjects treated with palivizumab. 5 However, the FDA did not approve licensure of motavizumab due to significant safety concerns, which included a 3-fold increase in nonfatal hypersensitivity adverse reactions compared with treatment with palivizumab. 4 So, while there are available highly potent anti-RSV mAbs, there remains an unmet need for a safe and effective prophylactic vaccine against RSV.
Vectored delivery of monoclonal Abs is a promising effective prophylaxis strategy against airborne infectious viruses. 6,7 Skaricic et al. were the first to report that delivery of an adeno-associated virus (AAV) rh10 (AAVrh10) vector expressing a murine version of palivizumab into the pleural space of mice resulted in high and sustained levels of palivizumab expression in systemic circulation, which in turn substantially reduced RSV A2 replication following an intranasal (IN) challenge. 8 While this was an important proof-of-principle study, the reduction in RSV titer was moderate in magnitude.
Adeno-associated virus serotype 9 (AAV9) has since emerged as a favorable vector for respiratory-targeted immunoprophylaxis due to its notable transduction of cells of the nasal airway epithelium (i.e., the site of infection of most airway-tropic viruses) and its unique ability to be readministered because it can effectively transduce the airway epithelium even in the presence of high-level serum-circulating AAV9-specific neutralizing ABs (NAbs), which are elicited by the immune system in response to the IN-administered AAV9 vector. 6,9,10 As such, we reasoned that if AAV9 vector was used to express palivizumab or motavizumab at the nasal mucosa, it could provide a safe and effective defense against the major pathogenic strains of RSV A and B. In this study, we engineered an AAV9 vector to express the scFy-immunoadhesin (IA) version of palivizumab or motavizumab and hypothesized that localized delivery to the mouse airway would effectively protect against a challenge with RSV and enable vector readministration, thus supporting the clinical development of this vectored prophylactic vaccine for seasonal use.
MATERIALS AND METHODS
Viral vector construction
We constructed expression cassettes encoding the variable regions of palivizumab [Protein Database (PDB): 2HWZ_H and 2HWZ_L, GenBank Identifiers (GI): 157835115 and 157835114 or motavizumab (PDB: 3IXT_H and 3IXT_L, GI: 284055645 and 284055644), linked to the constant (CH2 and CH3) domains of human immunoglobulin G1 (IgG1) to generate an Ab architecture known as an IA. A human IgG1 signal peptide was engineered to drive the cellular secretion of the IA. The sequences were synthesized de novo (Geneart, Life Technologies, Grand Island, NY) and cloned downstream of the hybrid cytomegalovirus (CMV) enhancer/chicken β-actin 7 (CB7) promoter and upstream of the rabbit beta-globin poly A signal. For the AAV9.201Ig-IA vector, we used the light and heavy chain sequences of an anti-SIVsmF236 Ab isolated from macaque to construct an IA using macaque IgG secretion signal and Fc domains. 11 We also generated an AAV9.ffLuc vector, which encodes the reporter gene firefly luciferase instead of an IA. This vector was used in mice to elicit anti-AAV9 NAbs before delivering the AAV9.IA vector. AAV9 vector stocks were produced as previously described. 6
RSV production
The RSV strain A2 (VR-1540; ATCC, Manassas, VA) was grown and quantified on Hep-2 cells as previously described. 8
Animal studies
Six-week-old female BALB/c mice (Jackson Laboratories, Bar Harbor, ME) were housed in the barrier side of the Animal Facility of the Translational Research Laboratories at the University of Pennsylvania. To express the various IA constructs by AAV9, mice were anesthetized by an intraperitoneal injection of ketamine/xylazine as previously described, and AAV9 vector, at doses varying from 1 × 109 to 1 × 1011 genome copies (GC) as described below, was delivered IN at a 50 μL volume.
6
For the RSV challenge, mice were inoculated IN with 2 × 106 plaque-forming units (PFU) of RSV strain A2 at a 50 μL volume, at either 14 days or 4 months after the vector treatment. Four days after the RSV challenge, at the peak of viral replication in control nonvector-treated animals, the mice were sacrificed and the lungs were harvested for RNA analysis. In one experiment, mice were first treated IN with AAV9.ffLuc, at either 1 × 108 or 1 × 109 GC, then assayed for NAb and were administered with
Bronchoalveolar lavage fluid, nasal lavage fluid, and serum collection
We harvested bronchoalveolar lavage fluid (BALF), nasal lavage fluid (NLF), and serum from mice at the time of necropsy, as previously described. 6,9
NAb assay
We evaluated serum-circulating NAbs against the AAV9 vector using the in vitro transduction inhibition assay on Huh7 cells as previously described. 12 The limit of detection of the NAb assay was a 1:5 serum dilution.
Enzyme-linked immunosorbent assay for IA expression
Enzyme-linked immunosorbent assay (ELISA) plates (Costar, high-binding EIA plate, cat.3369) were coated with 100 μL of 5 μg/mL protein A (Protein A from Staph aureus, Sigma, P6031-5MG; Thermo Fisher Scientific, Wilmington, DE and Sigma-Aldrich, St. Louis, MO) and stored overnight at 4°C. Plates were washed with 0.05% Tween-20/phosphate-buffered saline (PBS) and blocked with 10% bovine serum albumin (BSA)/PBS (Kirkegaard & Perry Laboratories). Samples were diluted in PBS and added to the plate for 1 h at 37°C. We used purified palivizumab (Synagis, MedImmune, Gaithersburg, MD) to generate a standard curve to determine the Pal-IA and Mot-IA concentrations. We detected binding using a biotin-SP-AffiniPure goat anti-human IgG Fcg-specific Ab (Jackson ImmunoResearch Laboratories, West Grove, PA) and a streptavidin-HRP conjugate (Abcam, Cambridge, MA). We developed plates with 3,3′,5,5′-Tetramethylbenzidine (TMB) substrate (Sigma-Aldrich) and measured absorbance at 280 nm in a NanoDrop 2000 (Thermo Fisher Scientific). ELISA Ab expression levels were normalized to total protein in the sample.
RNA preparation and cDNA synthesis
RNA was isolated with TRIzol (Life Technologies), treated with DNase I at 37°C for 30 min (Roche Applied Science, Indianapolis, IN), and eluted using an RNeasy spin column (QIAGEN, Valencia, CA) in 50 μL H2O. We quantified the RNA by spectrophotometry using a NanoDrop 2000 (Thermo Fisher Scientific) with cDNA synthesized using RT Random Primers provided with the Applied Biosystems® High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA).
Absolute qPCR for RSV RNA
To quantify the RSV RNA load, we performed real-time PCR with an Applied Biosystems 7500 using 1 μL cDNA in a total volume of 50 μL master mix with the following run conditions: 1 cycle of 2 min at 50°C and 10 min at 95°C followed by 40 cycles of 15 s at 95°C and 60 s at 60°C (Life Technologies). Primers (Forward: 5′-AAGACCAAAAACACAACAACAACCA-3′; Reverse: 5′-AGTGAAAATCATTATTGGGTTTGTTTGGTG-3′) amplified a 100-bp region in the RSV-G gene containing the TaqMan® FAM™-labeled probe (6FAM-AAGCCCACCACAAAAC-MGBNFQ). Reaction mixtures were set up in duplicates in 96-well plates. The standard was prepared by gene-specific reverse transcription of the hRSV Glycoprotein G gene from hRSV-infected mouse lung. This was followed by near end-to-end PCR amplification (918 bp) using a high-fidelity DNA polymerase (Phusion®, Thermo Fisher Scientific). We purified and cloned the PCR fragment into the PCR®-Blunt-TOPO plasmid of the Zero Blunt® TOPO Cloning Kit (Life Technologies). We confirmed both the PCR and the cloning products by sequencing and restriction digestion analyses. The TOPO-cloned Glycoprotein G gene was expanded, purified, and used as the real-time PCR standard following plasmid linearization with HindIII. We used eight serial dilutions of RSV-G at known concentrations to derive the standard curve. Standards and negative controls were run together with each qPCR assay.
Statistical analyses
We performed statistical analyses using the Prism 6 software (GraphPad Software, San Diego, CA). The unpaired two-tailed Student's t-test, assuming equal variance, was used for experiments comparing two groups. We used an analysis of variance (ANOVA) followed by the post-hoc Tukey's test to assess multiple (more than three) group comparisons. Statistical significance was determined at p < 0.05.
RESULTS AND DISCUSSION
AAV9-mediated airway expression of modified RSV antibodies
First, we characterized the expression profiles of two candidate AAV9 anti-RSV-IA vectors in mice. Specifically, BALB/c mice (n = 3/group) were dosed IN with 1 × 1011 GC of AAV9 vector expressing either the IA version of palivizumab (

AAV9-mediated expression of anti-RSV IA inhibits RSV replication in mouse airways.
AAV9 dose-dependent prophylaxis against RSV
We next evaluated the level of prophylaxis conferred by the IN-delivered AAV9 anti-RSV-IA vectors against an airway challenge with the RSV A2 virus. The RSV RNA load in the respiratory secretions of infants and adults, as measured by qPCR, has been shown to correlate with (1) RSV disease severity and (2) the number of viable virions in the airways.
13
–15
Moreover, changes in the level of the RSV RNA load in the lung of the challenge model can serve as an indicator of the efficacy of prophylaxis. We reasoned that AAV-mediated prophylaxis would result in lower viral RNA load in the lungs. BALB/c mice (n = 5/group) were administered with 1 × 10,
9
1 × 10,
10
and 1 × 1011 GC of
As a control, BALB/c mice were administered IN with 1 × 1011 GC of AAV9.201Ig-IA vector (a vector expressing an IA against simian immunodeficiency virus and irrelevant to RSV). Four days later, we harvested the lungs to analyze RSV load. We observed a significantly reduced viral load in all AAV9 treatment groups, with a maximal ∼17-fold decrease of RSV load between the untreated and
Longevity of AAV9-mediated protection against RSV infection
The RSV season typically ranges from 3 to 5 months, 1,2 so it is important that a vector-based prophylaxis confers sustained protective Ab expression in the airway. If an AAV9-based prophylaxis strategy for RSV could confer sustained protection for an entire season after a single administration of the vector, it would be an improvement over the repeated (up to five per season) monthly palivizumab injections currently administered to high-risk infants. This prophylaxis strategy would be an accessible prophylactic for the older at-risk populations that have no approved drug. Since the AAV9 vector does not integrate into the genome of transduced cells, we expected that the airway expression of Pal-IA and Mot-IA would steadily decline with the natural cell turnover of vector-transduced airway cells. 9 Therefore, central to the successful development of a sustained AAV vector-mediated prophylaxis for an entire RSV season is the ideal balance of Ab expression from the AAV-transduced cells and the turnover of transduced cells with time.
In this next set of studies, we assessed the longevity of AAV9-mediated protection against an RSV challenge. Mice were given IN 1 × 1011 GC of
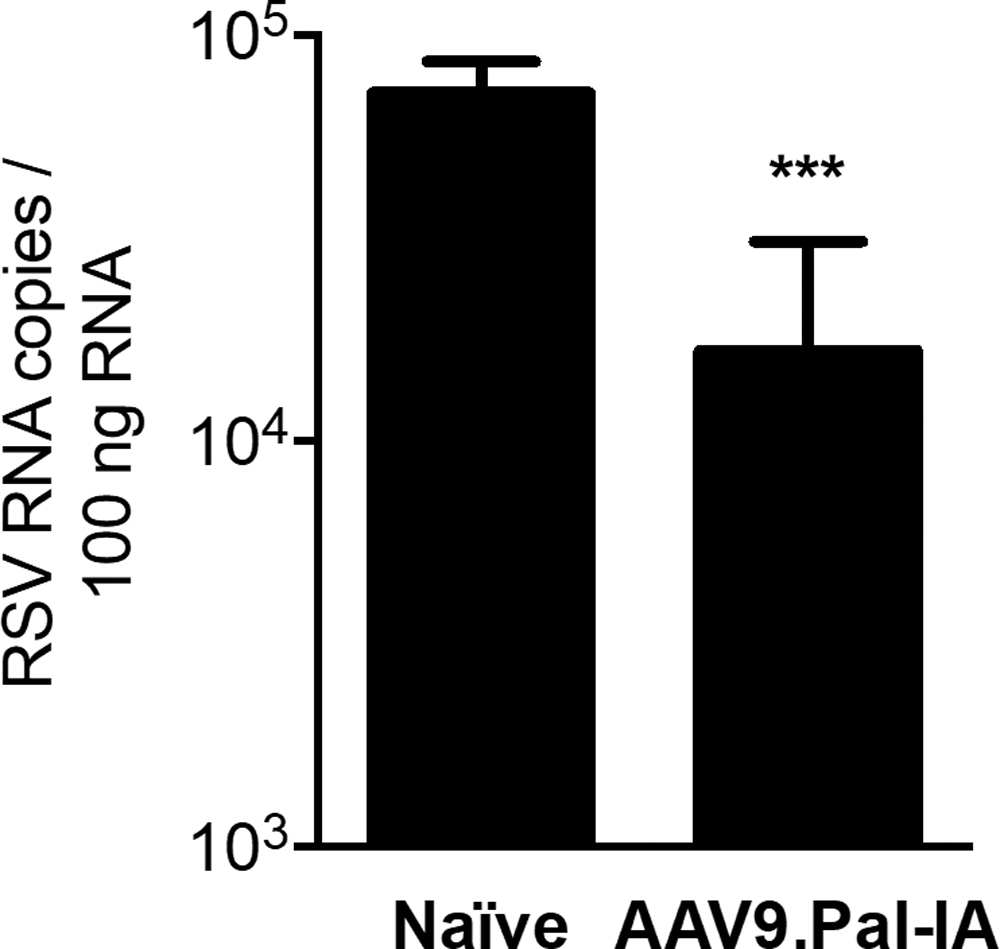
Sustained
The apparent loss of potency of the AAV9-Pal-IA vector may simply reflect the natural turnover of AAV9-transduced airway cells that contribute to the secretion of the protective Ab into the airway surface liquid layer. 9 However, the turnover of transduced cells with time adds to the safety profile of our proposed prophylaxis approach, as it minimizes sustained Ab production following the end of each RSV season.
Readministration of AAV9 to confer sustained prophylaxis against RSV
We propose that AAV9-based prophylaxis for airborne infectious viruses would be best suited for seasonal use. As such, it would be important to be able to readminister this product on a yearly basis. Whereas infants require protection for at least their first 2 years of life, older adults will likely need sustained protection for several years. The presence of preexisting NAbs to AAV9 may be problematic as high levels of circulating NAbs can hamper the efficacy of AAV gene transfer. 9,12,16 Previous reports have shown that up to 80% of healthy anti-AAV9 seropositive adults have NAb titers <1:200, with the majority of these titers being <1:20. 16 We selected AAV9 for our studies as this capsid effectively transduces cells of the mouse and nonhuman primate airways, 6 and can circumvent neutralization from a high level of serum-circulating AAV9-specific NAbs when administered IN. 9,10
We established a mouse model of preexisting anti-AAV9 NAbs by administering mice an AAV9 vector expressing a secretory product that is irrelevant to RSV. First, to determine the dose of AAV9 vector that elicits a clinically relevant level of anti-AAV9 NAbs, we administered IN mice (n = 3/group) with AAV9.201-IA vector at doses ranging from 1 × 108 to 1 × 1011 GC. Twenty-five days later, we observed a dose-dependent generation of serum-circulating anti-AAV9 NAb levels ranging from 1:5 to 1:5,120 (Fig. 3A). Based on this dataset, we administered IN mice (n = 5/group) with 1 × 108 or 1 × 109 GC of AAV9.ffLuc vector to produce clinically relevant levels of serum-circulating AAV9 NAbs. Seventy days later, we observed serum-circulating AAV9-specific NAbs ranging from 1:5 to 1:10 for mice administered IN 1 × 108 GC of AAV9.ffLuc vector, and 1:40 to 1:320 for mice administered IN 1 × 109 GC of AAV9.ffLuc vector (Fig. 3B, C). We administered these mice IN with 1 × 1011 GC of
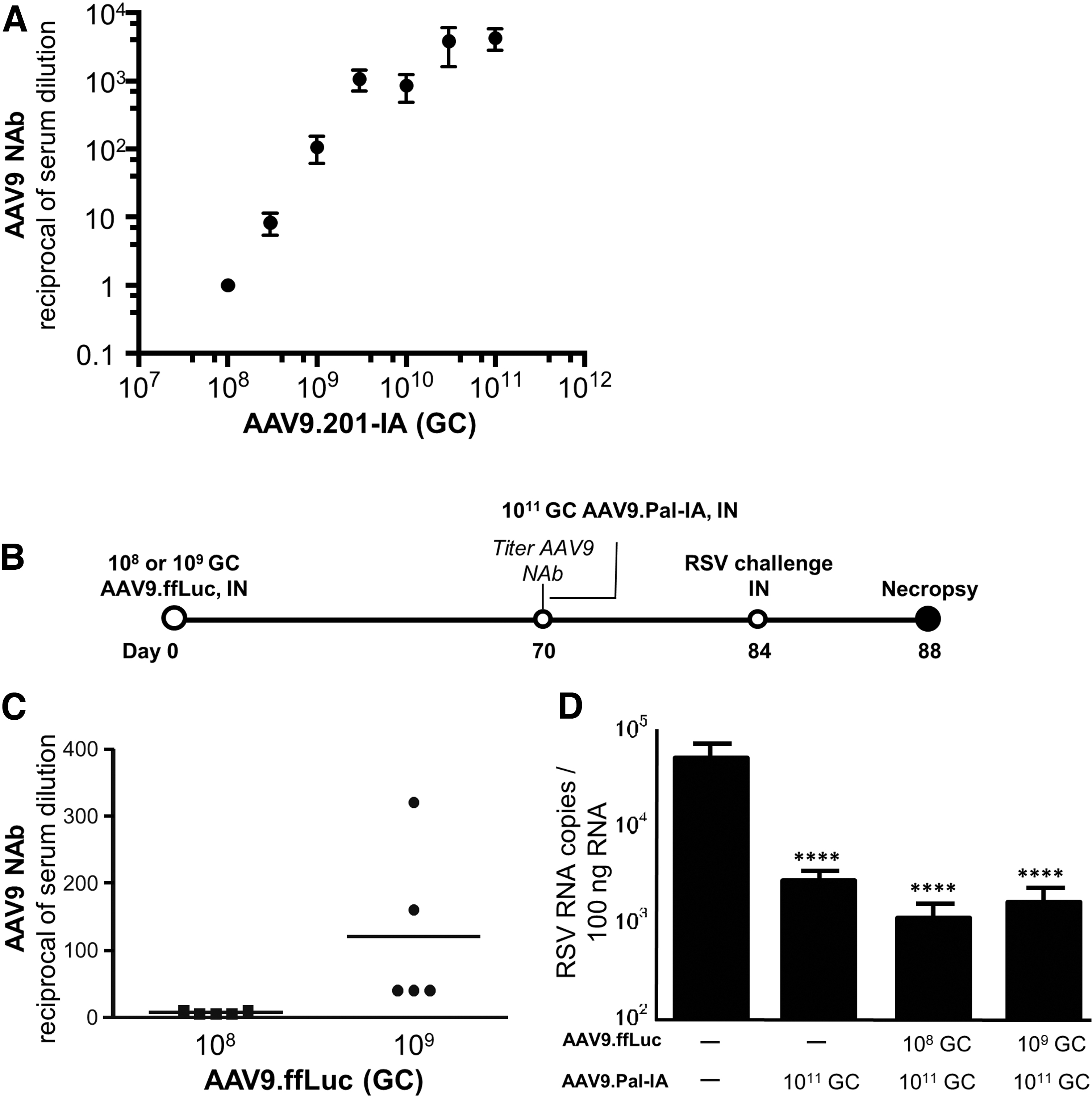
Effective AAV9-mediated prophylaxis against RSV despite presence of serum-circulating AAV9-specific NAb.
We observed a significant reduction in viral load for all mice treated with the
CONCLUSION
There is value to developing an AAV9-based prophylaxis strategy for RSV, which merits further efficacy testing in preclinical studies. Compared with the current standard of intramuscular palivizumab injection treatments, our vector-based prophylaxis strategy for RSV has the potential to confer longer lasting and more effective protection in a diverse subject population. Compared with the previously reported vector, our results suggest that IN administration of AAV9 may confer greater protection against RSV than intrapleural administration of AAVrh10, 8 perhaps because IN administration is a more clinically relevant route of delivery that provides protection along the route of infection.
Both palivizumab and motavizumab target a highly conserved region of the RSV fusion glycoprotein (F0), and both Abs have demonstrated in vivo efficacy against the RSV A and B strains. 2,3 Therefore, we predict these AAV9.IA vectors will be universally protective against major pathogenic strains of RSV. Moreover, the AAV9 vector has the necessary packaging capacity to potentially coexpress two different Abs against distinct respiratory infections caused by airborne infectious viruses, such as influenza 6,17 or coronaviruses, to provide a two-in-one defense strategy.
Footnotes
AUTHORS' CONTRIBUTIONS
J.T.: Data curation, formal analysis, investigation, writing—original draft preparation data; V.A., M.C., J.O., J.S.: data curation, formal analysis, investigation; A.P.T., S.R.: resources, S.W.: resources; J.M.W.: conceptualization, funding acquisition, M.P.L.: conceptualization, data curation, formal analysis, project administration, supervision, writing—final draft preparation.
ACKNOWLEDGMENTS
The authors thank the staff of the Program for Comparative Medicine and the Penn Vector, Immunology, and Cell Morphology Cores of the Gene Therapy Program at the University of Pennsylvania for their assistance with these studies. The authors thank Wendy Wu and Anurag Sharma at Cornell University for providing the RSV virus and Ronald Rubenstein at the Children's Hospital of Philadelphia for providing assay reagents.
AUTHOR DISCLOSURE
J.M.W. and M.P.L. have a pending patent application on AAV-mediated passive immunization of airborne pathogens. J.M.W. and M.P.L. are inventors on patents that have been licensed to various biopharmaceutical companies and for which they may receive payments. J.M.W. is a paid advisor to and holds equity in Scout Bio and Passage Bio; he also has sponsored research agreements with Amicus Therapeutics, Biogen, Elaaj Bio, FA212, Janssen, Passage Bio, and Scout Bio, which are licensees of Penn technology. He also has a sponsored research agreement with G2 Bio. J.M.W. holds equity in the G2 Bio-associated asset companies.
FUNDING INFORMATION
This work was supported by a research grant from DARPA/DOD #64047-LS-DRP.01 (J.M.W.).