
Abstract
The safety and high efficiency of adeno-associated virus (AAV) vectors has facilitated their wide-scale use to deliver therapeutic genes for experimental and clinical purposes in diseases affecting the central nervous system (CNS). AAV1, 2, 5, 8, 9, and rh10 are the most commonly used serotypes for CNS applications. Most AAVs are known to transduce genes predominantly into neurons. However, the precise tropism of AAVs in the dentate gyrus (DG), the region where persistent neurogenesis occurs in the adult brain, is not fully understood. We stereotaxically injected 1.5 × 1010 viral genomes of AAV2, 5, or rh10 carrying green fluorescent protein (GFP) into the right side of gerbil hippocampus, and performed immunofluorescent analysis using differentiation stage-specific markers 1 week after injection. We found that AAV5 showed a significantly larger number of double-positive cells for GFP and Sox2 in the DG, compared with the AAV2 and rh10 groups. On the contrary, AAVrh10 presented a substantially larger number of double-positive cells for GFP and NeuN in the DG, compared with AAV2 and AAV5. Our findings indicated that AAV5 showed high transduction efficiency to neural stem cells and precursor cells, whereas AAVrh10 showed much higher efficiency to mature neurons in the DG.
Introduction
Adeno-associated virus (AAV) vectors are widely used to deliver therapeutic genes for experimental and clinical purposes in diseases affecting the central nervous system (CNS). 1,2 AAV is a nonenveloped, single-stranded DNA virus with a diameter of 20–25 nm, and its extensive use in animal and human studies show that the virus is nonpathogenic. 3 In addition to their low immunogenicity and integrity, AAVs do not replicate in the absence of helper viruses such as adenovirus or herpesvirus. 4 These features make AAVs ideal vectors to transduce genes into the CNS.
The hippocampus plays a critical role in learning and memory 5 ; hence, it is a therapeutic target in many neurodegenerative diseases, such as Alzheimer's disease, 6,7 traumatic brain injury, 8,9 transient global ischemia, 10,11 and epilepsy. 12,13 In addition, the dentate gyrus (DG) in the hippocampus is a discrete region where neurogenesis occurs in the adult brain. 14 In 1965, Altman and Das reported a study using a tracking technique of [ 3 H]-thymidine incorporation into adult rat brains to suggest that neurogenesis takes place in the DG. 15 Evidence from numerous later studies also showed that neural stem cells in the subgranular zone (SGZ) of the DG continuously produce new neurons throughout life in different species, such as humans, mammals, and rodents. 16 Neural stem cells in the SGZ functionally integrate into the hippocampal circuit after some stages of proliferation and differentiation, and ultimately contribute to learning and memory. 17 Furthermore, it is well known that neurogenesis in the DG increases or decreases in some neurodegenerative diseases. For instance, several studies show that neurogenesis in the DG decreases in Alzheimer's disease, 18,19 but increases in traumatic brain injury, 20,21 transient global ischemia, 22,23 and epilepsy. 24 Much remains unelucidated about the mechanisms and the roles of adult neurogenesis.
The increasing demand of AAVs in both clinical and experimental settings makes the precise tropism of AAVs in the DG an area of interest. At present, AAV1, 2, 5, 8, 9, and rh10 are the most commonly used serotypes for CNS applications. However, no previous report has shown the detailed tropism of AAVs in the DG thus far. Therefore, we compared transgene expression in the DG 7 days after focal injection of AAV2, 5, or rh10, especially focusing on transduction to the neural stem cells in the SGZ of the DG.
Materials and Methods
Plasmids and recombinant AAV production
To compare the tropism of recombinant AAV vectors derived from different AAV serotypes, AAV-CMV-humanized recombinant green fluorescent protein (hrGFP) (Agilent Technologies, CA) plasmid was used (Fig. 1A). Recombinant AAV-CMV-hrGFP (serotypes 2, 5, and rh10) was prepared as previously described. 25 In brief, 60% confluent human embryonic kidney cells (HEK293; Agilent Technologies) were incubated in large culture vessels and cotransfected with the AAV vector plasmid, adenoviral helper plasmid pHelper (Agilent Technologies), and one other helper plasmid, including pAAV-RC (Agilent Technologies) for AAV2, pAAV5 Rep/AAV5 Cap for AAV5, or pAAV2 Rep/AAVrh10 Cap for AAVrh10. Crude viral lysates were purified twice on a CsCl two-tier centrifugation gradient. Viral titers were determined by quantitative PCR of the simian virus 40 poly(A) sequence, and the viral stock in this study was dissolved in HN buffer [50 mmol−1 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, pH 7.4, 150 mmol−1 NaCl] and adjusted to the final concentration of 7.5 × 109 viral genomes (vg)/μL.
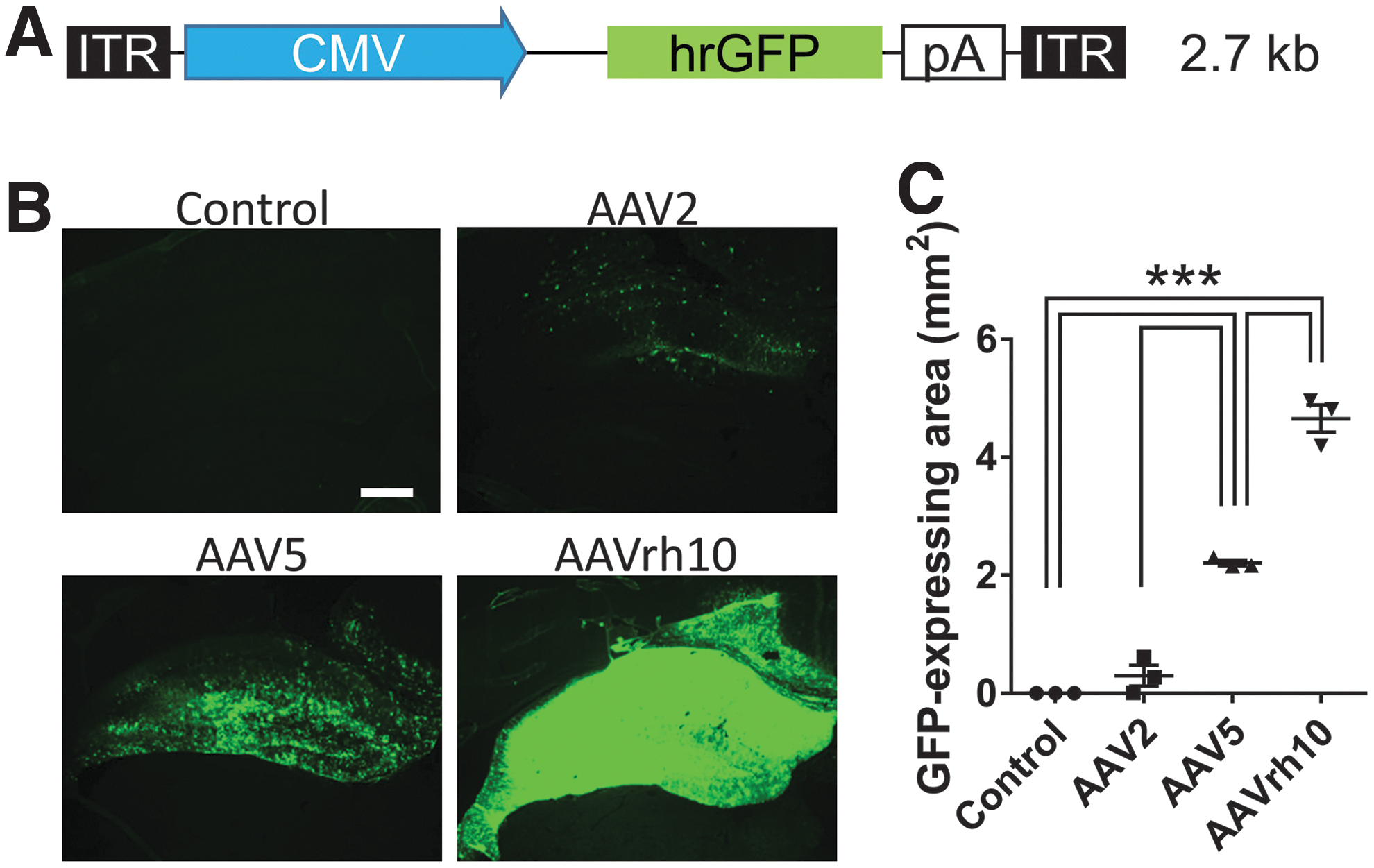
Vector construction and transduced gene expression of the hippocampus on the injected side.
Stereotaxic injection
All experiments were carried out in full compliance with the guidelines from the Institutional Animal Care and Use Committee of Jichi Medical University. Twelve male gerbils (4 weeks old, body weight: 28–40 g, from Japan SLC, Japan) were anesthetized with 2% isoflurane and placed in a stereotaxic apparatus (Narishige, Japan) (three animals per group). Viral stocks or phosphate-buffered saline (PBS) were injected at a rate of 0.2 μL/min through a 36-gauge glass cannula connected to a 10 μL Hamilton syringe mounted to a microinjection pump (Univentor, Malta). The stereotaxic injection coordinates for DG of hippocampus were 2.0 mm caudal to the bregma, 1.5 mm right to the midline, and 2.5 mm below the pial surface with a 2 μL injection (1.5 × 1010 vg in total). The needle remained in place for 5 additional minutes before the cannula was slowly withdrawn to prevent reflux. The skin incision was closed with a suture and the animal was kept warm using a heat blanket before being returned to the cage.
Histology and immunohistochemistry
One week after injection of the viral stocks or PBS, gerbils were decapitated under deep anesthesia, and their brains were quickly removed and frozen with powdered dry ice. Then, 10 μm coronal sections of the brains were cut on a sliding microtome (Leica, Germany). The sections were incubated with primary antibody against NeuN (diluted at 1:1000; MAB377, Merck, Germany), Sox-2 (diluted at 1:1000, ab79351; Abcam, United Kingdom), glial fibrillary acidic protein (GFAP) (diluted at 1:1000, #840001; Biolegend, CA), Sox-1 (diluted at 1:1000, ab87775; Abcam, United Kingdom), and Ki-67 (diluted at 1:1000, ab15580; Abcam, United Kingdom). These were placed in PBS containing 0.3% Triton X-100 at 4°C overnight followed by incubation with a secondary antibody against Alexa Fluor 555 anti-mouse immunoglobulin G (IgG; diluted at 1:1000, #4409; Cell Signaling, MA) or Alexa Fluor 546 anti-rabbit IgG (diluted at 1:1000, A10040; ThermoFisher Scientific, MA) and Alexa Fluor 647 anti-mouse IgG (diluted at 1:1000, A21235; ThermoFisher Scientific) for 1 h at room temperature. After washing with PBS, the sections were coverslipped with mounting agent including 4′,6-diamidino-2-phenylindole (DAPI) (ThermoFisher Scientific). In Fig. 1, the GFP-positive area was automatically measured using the BZ-II analyzer software (Keyence, Japan). The GFP-positive area of one animal was defined as the mean of three sections at 1.7, 2.0, and 2.3 mm caudal to the bregma. In Figs. 2 and 4, immunoreactivity was assessed under a microscope (BZ-9000; Keyence, Japan) and the number of immunopositive cells in the DG of one animal was defined as the mean of three sections of whole DG at 1.7, 2.0, and 2.3 mm caudal to the bregma. Cell counts were carried out by an investigator who was blind to the animal groups. The SGZ was defined within 20 μm from the inner surface of the granule cell layer.
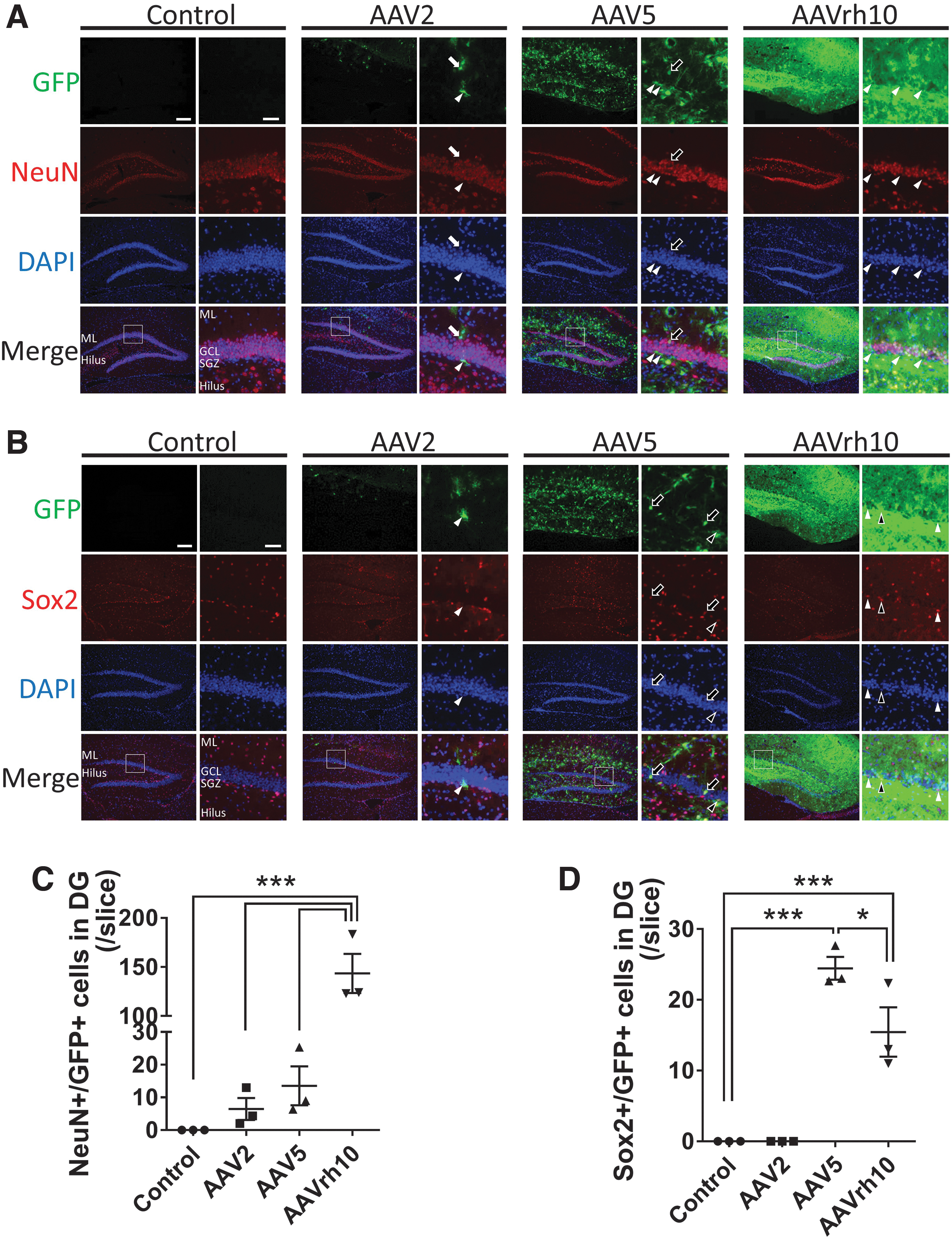
Different tropism of AAV2, 5, and rh10 in the DG.
Statistical analysis
The results are presented as the mean ± standard error of mean. Statistical analysis was performed using GraphPad Prism 6.0, and comparisons between groups were conducted using one-way analysis of variance (ANOVA) with Tukey's post hoc test for multiple comparisons. Statistical significance was defined as p < 0.05.
Results
Gene expression area of AAV2, AAV5, and AAVrh10 in gerbil hippocampus after stereotaxic injection
To investigate the gene transduction efficiency of three different AAVs, such as AAV2, AAV5, and AAVrh10, we stereotaxically injected 1.5 × 1010 vg of one of the AAVs carrying GFP into the right hippocampus. We then evaluated the GFP-expression pattern using three coronal sections of the same animal 1 week after stereotaxic injection (Fig. 1B). We found that the smallest expression area was that of AAV2, whereas the largest was expressed by AAVrh10, and AAV5 was in the middle (AAV2: 0.30 ± 0.17 mm2, AAV5: 2.21 ± 0.05 mm2, AAVrh10: 4.66 ± 0.23 mm2, p < 0.001, for AAV5 and AAVrh10, compared with the control; Fig. 1B, C). AAV5 and AAVrh10 both showed the GFP expression throughout the ipsilateral hippocampi with little variance between each animal within the same group. On the contrary, AAV5 showed a patchy pattern, whereas AAVrh10 was homogeneous with almost no gap of the GFP expression.
Quantification of number of transduced mature neurons and neural stem cells in DG
To reveal the reason why each AAV showed a different gene expression pattern in the hippocampus, we first analyzed immunohistochemistry for two markers, including NeuN, a marker for mature neurons, and Sox2, a marker for neural stem cells to quantify double-positive cells with GFP in the DG. AAVrh10 showed a significantly larger number of double-positive cells for NeuN and GFP, compared with AAV2 and AAV5 in the granule cell layer of the DG (AAV2: 6.4 ± 3.3, AAV5: 13.6 ± 5.9, AAVrh10: 143.4 ± 20.0, p < 0.001 for AAVrh10 compared with the control; Fig. 2A, C). On the contrary, AAV5 showed a significantly larger number of double-positive cells for Sox2 and GFP, compared with AAV2 and AAVrh10 in the granule cell layer and SGZ of the DG (AAV2: 0.0 ± 0.0, AAV5: 24.4 ± 1.6, AAVrh10: 15.4 ± 3.5, p < 0.001 and p < 0.05 for AAV5 compared to the control and AAVrh10, each; Fig. 2B, D).
In addition, we further evaluated immunohistochemistry for GFAP and Sox1, both of which are also markers for neural stem cells (Fig. 3). 26 AAV2 showed few GFP-positive cells and no triple-positive cells for GFP, Sox2, and GFAP or Sox1 in the SGZ. AAV5 and AAVrh10 showed triple-positive cells for GFP, Sox2, and GFAP or Sox1, but not all the double-positive cells for Sox2 and GFAP or Sox1 were also positive for GFP in the SGZ. AAV5 showed a significantly larger number of double-positive cells for GFAP and GFP, compared with AAV2 and AAVrh10 in the SGZ of the DG (AAV2: 0.0 ± 0.0, AAV5: 9.4 ± 1.9, AAVrh10: 4.2 ± 0.6, p < 0.001 and p < 0.05 for AAV5 compared with the control and AAVrh10, each; Fig. 3A, C). Concerning Sox1, AAV5 showed a significantly larger number of double-positive cells for Sox1 and GFP, compared with AAV2, but not with AAVrh10, in the in the SGZ of the DG (AAV2: 0.0 ± 0.0, AAV5: 12.6 ± 2.1, AAVrh10: 15.3 ± 3.7, p < 0.05 for AAV5 compared with the control, p < 0.01 for AAVrh10 compared with the control; Fig. 3B, D).

Double immunofluorescent staining of makers for neural stem cells and neural progenitor cells in the DG.
Quantification of number of transduced proliferating neural stem cells in SGZ
We further investigated the transduction efficiency of each AAV to the proliferating neural stem cells of the SGZ. We analyzed immunohistochemistry of Ki-67, which is expressed during all active phases of the cell cycle (late G1, S, G2, and M phases) except the resting phase and has been used as a proliferation marker. 27,28 AAV2 showed no double-positive cells for GFP and Ki-67 in the SGZ. AAV5 and AAVrh10 showed a significantly larger number of double-positive cells for GFP and Ki-67 in the SGZ (AAV2: 0.0 ± 0.0, AAV5: 5.2 ± 1.1, AAVrh10: 3.2 ± 0.3, p < 0.01 and p < 0.05 for AAV5 and AAVrh10 compared with the control, each; Fig. 4). AAV5 showed a larger number of double-positive cells for GFP and Ki-67 in comparison with AAVrh10, but there was no statistical significance between AAV5 and AAVrh10.
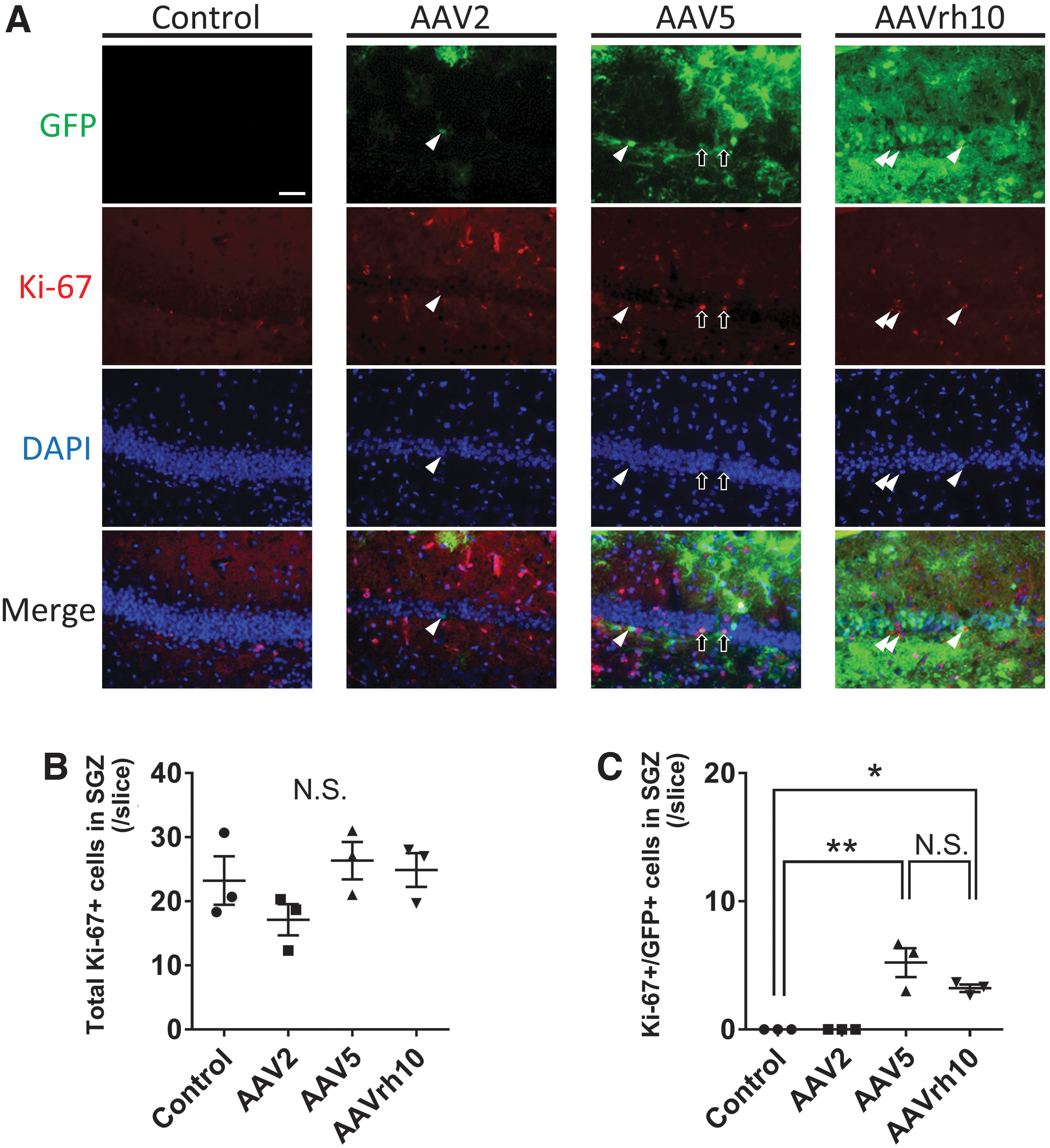
Immunoflorescent staining of Ki-67 in the DG.
Discussion
AAV vectors are promising tool for gene replacement in neurodegenerative diseases owing to their long-term efficacy and safety. 2,3,29 The pathophysiology of each neurodegenerative disease is different; hence, it is necessary to choose the appropriate AAV serotype that is ideal for the experiment or intervention. As no previous report has focused on the gene transduction efficiency to the neural stem cells and immature neurons in the DG of the hippocampus, we compared the tropism of the AAV2, 5, and rh10 7 days after stereotaxic injection into gerbil DG, which would be helpful for the safe and efficient gene transduction.
It is important to note that the mechanisms of gene transduction through AAV are not fully elucidated; however, most AAVs are known to transduce genes predominantly into neurons. Furthermore, the tropism of AAVs is determined by the vector capsid (serotype), vector purification method, administration route, and the maturity of the host. 30 –33 The use of a cell-specific promoter can be a possible strategy to transduce genes into a particular type of cells, such as oligodendrocytes and astrocytes. 32,34
The reason why we chose three different serotypes of AAV, such as AAV2, 5, and rh10, in this study is that these serotypes have been reported to have different features: (1) AAV1, 9, and rh10 show the widespread and strong transgene expression with neuronal tropism 30,33,35 ; (2) AAV5 has strong glial tropism as given in Supplementary Fig. S1 36,37 ; and (3) AAV2 is suitable to transduce genes in the limited region because of its less diffusion in brain tissue compared with other serotypes. 33,38
In this study, we chose gerbils to investigate the tropism to neural stem cells. Gerbils are often used to investigate the mechanisms of neurogenesis in the DG after transient ischemia because we can stably reproduce ischemia of the hippocampus because of their insufficient collateral blood supply from posterior circulation. 39,40 We can also use rats and mice for global ischemia, although a 2-day operation is necessary to occlude the blood flow from both the carotid (anterior) and the vertebral-basilar (posterior) system. As a limitation of this study, we should also note that there are species-specific differences of AAV tropism between rodents, nonhuman primates, and humans. 41
In this study, we found distinctly different expression patterns of transduced GFP between AAV5 and rh10 in the DG 1 week after stereotaxic administration. There was little variance of GFP-expressing areas from animal to animal in the same group, which means the administered vectors of 1.5 × 1010 vg were sufficient to transduce genes into the gerbil hippocampus of the injected side (Fig. 1). Furthermore, immunohistochemical analysis revealed a significantly higher number of double-positive cells for GFP and Sox2 in the SGZ and granule cell layer in AAV5, compared with AAV2 and rh10. On the contrary, AAVrh10 had robustly higher efficiency of gene transduction to mature neurons in the granule cell layer, in comparison with AAV2 and AAV5 (Fig. 2). In short, our study showed transduction preference of AAV5 to neural stem cells and progenitor cells, whereas preference of AAVrh10 to mature neurons was displayed in the DG.
In the adult brain, neurogenesis occurs in two restricted regions that include the subventricular zone of the lateral ventricle and the SGZ of the DG. 42 With regard to DG, radial glia-like stem cells in the SGZ give rise to the granule cells and astrocyte throughout life. 16 It is important to note that neurogenesis in the DG generates only granule cells. In the original publications, these neural stem cells were identified only through the detection of their proliferative activity, the absence of the mature morphological characteristics, and specific markers of mature neurons, such as NeuN. 15,43 To date, a set of marker antigens has been identified at each different stage of adult neurogenesis, as summarized in Fig. 5. 43,44 The marker proteins have different functions, such as cytoskeleton (doublecortin [DCX], 45 GFAP, 46 and nestin 47 ), cell adhesion (polysialic acid-neural cell adhesion molecule [PSA-NCAM], 48 ), intracellular signaling, RNA splicing (NeuN, 49 ), and transcription factor (Sox2, 50,51 ). Hence, the markers do not necessarily overlap although the cells are on the same differentiation stage as was shown in the double immunostaining in Fig. 3.

Graphical summary of this study. Radial glia-like stem cells in the SGZ of the DG persist cell division even in the adults with some daughter cells, which give rise to the granule cells that functionally incorporate into the hippocampal circuit and contribute to learning and memory. Our immunofluorescent analysis using stage-specific markers found that AAV5 showed higher transduction efficiency into neural stem cells and progenitor cells compared with AAVrh10 in the gerbil DG. On the contrary, AAVrh10 showed far higher transduction efficiency into mature neurons compared with AAV5.
Sox2 is a well-characterized marker that is highly expressed in neural stem cells and progenitor cells and is usually downregulated in differentiating cells. 50 Sox2 is a member of the Sox gene family that acts as a transcriptional factor in neural stem cells, and is required for their maintenance of self-renewal capacity. 52 Although less reports have been published, Sox1 is also a member of the Sox gene family and is expressed in the GFAP-positive radial glia-like stem cells and Sox2-positive nonradial cells. 53 In contrast to Sox2, Sox1 shows a much more restricted pattern than other previously reported markers, as summarized in Fig. 5. Also in our results of Fig. 3, the number of Sox1-positive cells were less than Sox2-positive cells, which did not show significant difference between AAV5 and rh10.
GFAP, an established marker for mature astrocytes in the adult brain, is also positive in radial glia-like stem cells in the SGZ. 54 The majority of GFAP-positive cells in the whole hippocampus are mature astrocytes as illustrated in Supplementary Fig. S1. Hence, this study viewed the need to determine tropism in the DG separately. Aschauer et al. analyzed transduction profile of six different AAV serotypes, including AAV1, 2, 5, 6, 8, and 9, in the whole hippocampus of mice 3 weeks after stereotaxic administration. 4 They showed that areas of transgene expression were the widest in AAV5 and 9, which were significantly wider than AAV2. However, transduction efficiency of specific cell types such as NeuN- or GFAP-positive cells did not show significant differences between each serotype, although AAV8 exceptionally displayed higher efficiency for GFAP-positive cells in comparison with AAV2. Burger et al. found that AAV5 presented almost exclusive patterns of transduction into the neurons of a rat hippocampus 4 weeks after stereotaxic injections. 33 On the contrary, our group previously showed higher transduction efficiency of AAV2 to MAP2-positive cells and AAV5 to GFAP-positive cells using rat cortical primary culture, although the results in vitro does not necessarily imply the same tropism of AAVs in vivo. 36 The findings mentioned previously suggest that AAV5 prefers neural stem and progenitor cells and the difference between each serotype may be underestimated if quantitative analysis is performed as the unit of the whole hippocampus. In our results given in Supplementary Fig. S1, AAV5 showed substantial GFP transduction to mature astrocytes outside the granule cell layer and SGZ, which was not a focal point in this study. The common underlying mechanisms may include AAV5 transduction of genes into neural stem cells and mature astrocytes because neural stem cells have astrocytic features. 55 In addition, we should note that the difference of gene promoters, virus purification methods, and animal species might affect the tropism of AAVs.
Ki-67 is a marker for cell proliferation that is expressed in dividing cells and is strongly downregulated or absent in quiescent cells. 28 Although neurogenesis in the SGZ is promoted in some neurodegenerative disorders such as traumatic brain injury, transient global ischemia, and epilepsy, it is predominantly regulated to a basal level under physiological conditions. The animals in this study were under healthy condition; therefore, we found not many Ki-67-positive cells (Fig. 4B). The number of double-positive cells for GFP and Ki-67 were significantly larger in the AAV5 and rh10 groups in comparison with the control and AAV2 groups (Fig. 4C). Although not significant, AAV5 showed higher number of double-positive cells for GFP and Ki-67, compared with AAVrh10. This finding was consistent with the immunostaining for Sox2.
Conclusion
AAV5 showed higher transduction efficiency into neural stem cells compared with AAVrh10 in the gerbil DG. On the contrary, AAVrh10 showed far higher transduction efficiency into mature neurons compared with AAV5. Although the tropism of AAVs is known to be mostly forwarded to neurons, we should note the precise tropism for the safe and efficient gene transduction.
Footnotes
Authors' Contributions
Y.S. conceived and planned the experiments. Y.S., R.U., and K.S. carried out the experiments. Y.S. performed analysis of the data. Y.H., K.O., M.U., A.I., K.K., and H.M. contributed to the interpretation of the results. Y.S. prepared figures and drafted the article. All authors reviewed and approved the final version of the article.
Acknowledgments
The authors thank Miyuki Kondo, Miyoko Mitsu, and Satomi Fujiwara for their technical assistance.
Author Disclosure
No competing financial interests exist.
Funding Information
This work was supported by JSPS KAKENHI (Grant No. 17K10846), Taiju Life Social Welfare Foundation, and Japan Brain Foundation (to Y.S.).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.