
Abstract

CRISPR/C
They reported both humoral and cytotoxic T lymphocyte (CTL) responses to AAV-mediated Cas9 in normal and dystrophic canines, which were only observed against Cas9 and no other transgenes (Fig. 1). Although further characterization of the immune responses to editing machinery is needed, these results suggest that the immune response to AAV-CRISPR may be a critical barrier to the advancements of CRISPR-based therapies.
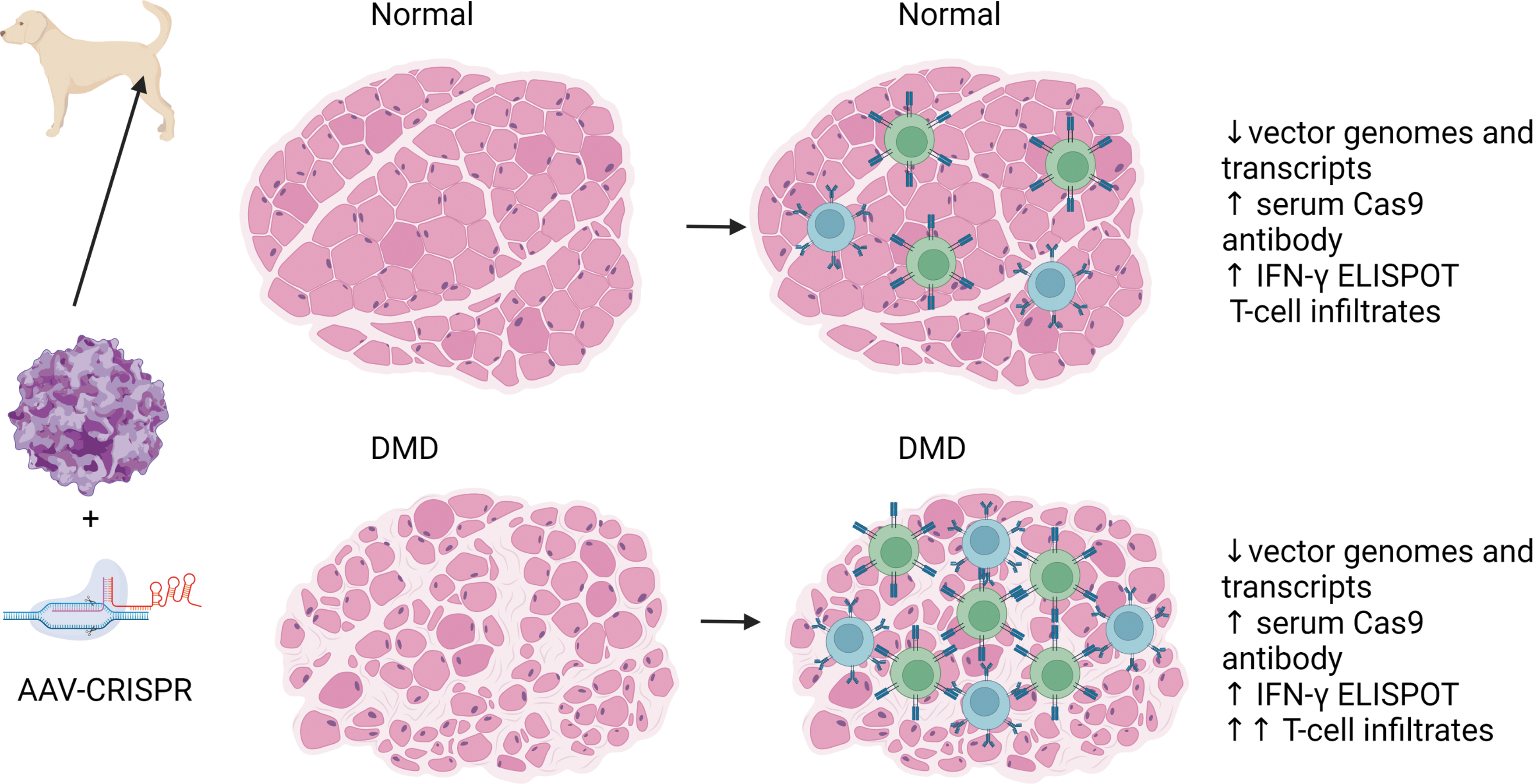
In the Hakim et al. study, 1 when AAV capsids are used to deliver Cas9 proteins in dog models, both normal and DMD models result in loss of vector genomes and transcripts, and an increase in serum Cas9 antibodies and IFN-gamma ELISPOTs. Considerable T-cell inflammation was observed in the DMD dogs treated with AAV-Cas9. AAV, adeno associated virus; DMD, Duchenne muscular dystrophy; IFN, interferon; ELISPOT, enzyme-linked immune absorbent spot.
Pre-existing immunity to AAV capsid has been a hurdle for the clinical development of gene therapy. Furthermore, natural immunity to Cas9 in humans has been documented, including both B cell and T cell responses. 2 –4 Although pre-existing humoral response may not limit therapeutic potential of the intracellular Cas9 protein, patients with pre-existing CTLs to Cas9 could pose several issues for therapeutic potential. In addition, limiting treatments to patients without pre-existing immunity to both AAV and Cas would likely severely limit the number of patients available for treatment. Mouse studies revealed that if mice were sensitized to Cas9 before AAV-CRISPR/Cas9 delivery, edited cells were cleared by CTL responses. 5
In Hakim's study, they found that high levels of anti-Cas9 antibodies were detected in adult dogs, but puppies between 2 and 6 weeks of age had barely detectable titers, which suggested they were immunologically naive to Cas9. Most of the dogs dosed in this study were treated as puppies at 1 month of age and developed CTL responses that resulted in loss of transduced cells. Similarly, mouse studies have reported humoral and cellular immune responses in mice lacking pre-existing immunity. 6 –9 Taken together these data suggest that the antitransgene immunity to Cas9 may be an issue in clinical translation even in patients without pre-existing immunity to Cas9. However, in a mouse study, serial rounds of redosing of AAV-CRISPR were tolerated using divergent Cas protein homologs and AAV capsid variants. 8
Although several factors can contribute to antitransgene immunogenicity, the transgene itself plays an important role in the development of immune responses particularly if novel epitopes are introduced and there is a lack of self-tolerance to the delivered transgene. This is observed in cross-reactive immune material negative patients who have null mutations for Pompe disease and, therefore, have no immunological self-recognition of the acid α-glucosidase enzyme, which results in immune responses to both protein replacement therapy and AAV gene therapy through production of antitherapeutic antibodies. Antitransgene T cells responses were documented in intramuscular gene transfer in DMD after delivery of the mini-dystrophin transgene. 10
In addition, in an alpha-1-antityrpsin (AAT) deficiency clinical trial, immune responses resulting in reduced transgene expression were observed in one patient with a common single amino acid polymorphism that not represented in the wild-type transgene. 11 The immune response observed in this study in canines was specific for the Cas9 protein, which is bacterial derived and produces T cell response in humans. Induction of immune response and clearance of transcripts was not described in control animals injected with viruses containing the gRNA without co-injection of Cas9. Others have shown that engineering Cas9 to remove epitopes that are recognized by CTLs could potentially eliminate transgene immune responses to Cas94 and should be considered for further clinical development.
Many strategies have been deployed to minimize immune responses to AAV capsids and transgene. Targeting different tissues for expression by either the route of injection or promoter selection can alter the immunogenicity of an AAV gene therapy. Hakim et al. tested both intramuscular route and systemic route of injection and minimized off-target expression by using a muscle-specific promoter; however, under these conditions, cellular immune responses cleared vector genomes and transcripts.
Although in humans, muscle-directed targeting has been shown to be tolerogenic, inducing T-regulatory cells and providing long-lived transgene expression in clinical trials of AAT deficiency 12 –16 and lipoprotein lipase (LPL) deficiency, 17 –19 in proinflammatory environments, similar to those associated with dystrophies, muscle-specific delivery could lead to increased transgene immunogenicity. 10,20,21 However, T cell responses were observed in three dystrophic canine models, and also normal canines, even at a lower dose. 1
Immune-suppressive drugs have been widely used to control immune responses to AAV capsids and transgenes. Corticosteroids are used in the approved gene therapies for Spinal Muscular Atrophy 22 and inherited retinal dystrophy 23 to control cellular immune responses; however, several studies have indicated immune responses despite corticosteroid usage and additional immune suppression may be necessary especially for higher dose therapies. Prophylactic high-dose prednisolone treatment did not halt the clearance of edited cells in DMD dogs described in this study. 1
Immunosuppression has been achieved by treatment with a combination of Sirolimus and Rituximab to combat cellular and humoral immune responses to capsid and transgene, to suppress T cells, and to ablate B cells before AAV delivery in Pompe disease patients 24,25 and is being tested for readministration of AAV vectors. 26 However, humoral immune responses may return after immunosuppression is removed. 27 Other immunosuppressive regimens have been considered, including mycophenolate mofetil, calcineurin inhibitors such as tacrolimus, and proteasome inhibitors, and often these drugs are used in combination. Additional immunosuppressive drugs in combination with corticosteroids may be able to overcome the immune response to Cas9 in larger animal models.
Induction of suppression by targeting regulatory T cells (Tregs) may be an effective way to overcome immune responses to AAV-Cas9. Induction of endogenous Tregs in AAT and LPL deficiency have resulted in continued long-term transgene expression despite the development of T cell responses. Tolerance and long-lived expression was observed in Hakim et al. in control vectors expressing SERCA2a and micro-dystrophin, which persisted 84 weeks postinjection in DMD dogs and was associated with Treg infiltration. 1 Some immune-suppressive drugs, such as Sirolimus, inhibit cytotoxic and helper T cells, but also induces Tregs. 28,29 It has been suggested that either polyclonal or chimeric antigen receptor (CAR) Tregs could be beneficial in suppressing immune responses to AAV gene therapy. 30
CAR Tregs specific for AAV capsid have been shown to suppress both capsid and transgene immune responses in mouse models. 31 Others have suggested that Cas9-specific Tregs to mitigate immune responses. 32 A substantial proportion of Cas9-specific Tregs were discovered in human donors after Cas9 stimulation, 3 and these could potentially be expanded or a Cas9-specific CAR Tregs could additionally be generated to be used to suppress immune responses to Cas9.
Although immunogenicity to Cas9 may create a barrier to clinical translation of CRISPR-based editing, researchers have several tools to overcome those barriers, including immune orthologs or engineered Cas9 that removes immunogenic epitopes to prevent CTL responses as well as promoting tolerance by induction or expansion of specific Tregs or development of CAR Tregs.