
Abstract
Tuberculosis (TB) has been for many years a major public health problem since treatment is long and sometimes ineffective favoring the increase of multidrug-resistant mycobacteria (MDR-TB). Gene therapy is a novel and effective tool to regulate immune responses. In this study we evaluated the therapeutic effect of an adenoviral vector codifying osteopontin (AdOPN), a molecule known for their roles to favor Th1 and Th17 type-cytokine expression which are crucial in TB containment. A single dose of AdOPN administration in BALB/c mice suffering late progressive pulmonary MDR-TB produced significant lower bacterial load and pneumonia, due to higher expression of IFN-γ, IL-12, and IL-17 in coexistence with increase of granulomas in number and size, resulting in higher survival, in contrast with mice treated with the control adenovirus that codify the green fluorescent protein (AdGFP). Combined therapy of AdOPN with a regimen of second line antibiotics produced a better control of bacterial load in lung during the first days of treatment, suggesting that AdOPN can shorten chemotherapy. Taken together, gene therapy with AdOPN leads to higher immune responses against TB infection, resulting in a new potential treatment against pulmonary TB that can co-adjuvant chemotherapy.
INTRODUCTION
M
During the start of Mtb infection, the immune system activates a Th1-type response with the expression of IFN-γ, TNF-α, and IL-12, polarizing macrophages to M1 phenotype which in some individuals is efficient to control Mtb growth, eliminating the infection or inducing the bacteria to a latent state with the formation of granulomas. 2 In contrast, during late progressive disease, chronic inflammation drives the immune system to a Th1/Th2 cytokine profile, macrophage exhaustion, impaired activation of dendritic cells, and FoxP3+ Treg activation and proliferation. 3 –5 All these responses lead to an upregulated expression of the anti-inflammatory cytokines IL-4, IL-13, IL-10, TGF-β and in consequence the decrease of pro-inflammatory cytokines like TNF-α, IL-1α, IFN-γ which in turn favors granuloma disruption and mycobacterial growth leading to pulmonary tissue damage that can cause death. 6 –8
Many regulatory proteins with diverse functions also participate in the immune and inflammatory response against Mtb. One of them is osteopontin (OPN), which is a multifunctional protein involved in biomineralization, wound healing, and inflammation, that has been found in several types of cancer, diabetes, and nephrolithiasis. Different types of cells express OPN, including macrophages and T cells; however, OPN can modulate functions in these cells, as well as in dendritic cells, NK cells, B cells, and neutrophils.
The main role of OPN in inflammatory processes is inducing chemotaxis of these immune cells. 9 It has been reported that OPN also induces maturation of dendritic cells, activation of T cells expressing IFN-γ, and IL-12 prompting a Th1 or Th17 polarization. 10 –12 Indeed, all these immune responses are protective in pulmonary TB. 4 Thus, mice OPN null infected intraperitoneally with Mycobacterium bovis BCG (bacillus Calmette-Guérin) showed higher bacterial load compared to wild type mice 13 ; macrophages derived from monocytes infected with Mtb strain H37Ra and incubated with OPN had fewer bacteria (colonies forming units [CFU] count). 14 Thus, it seems that OPN has a protective activity in pulmonary TB.
To seek for an improvement treatment of progressive TB, trying to shorten the time of treatment that can prevent or decrease the toxic effects and abandonment of chemotherapy, we proposed a gene therapy approach, by increasing the expression of OPN using recombinant adenoviral vector (Ad5). Theoretically, this kind of treatment should increase the expression of pro-inflammatory cytokines (IFN-γ, IL-12, and IL-17) and polarization of macrophages and T cells to M1 and Th1 phenotype, respectively, that efficiently contain and eliminate Mtb. Thus, in the present work we used an experimental model of TB with BALB/c mice infected with MDR-TB.
In the first part of this work, we determined the kinetics of OPN gene expression during the course of the infection by reverse transcriptase polymerase chain reaction (RT-PCR) and the OPN cellular sources by immunohistochemistry. Then infected mice after 2 months of infection were treated by intratracheal route with a single dose of the recombinant adenoviral vector that expresses OPN (AdOPN) alone or in combination with second line antibiotics scheme (intragastric) and determined bacillary loads and extension of tissue damage (pneumonia) to evaluate its therapeutic efficiency, as well as protective cytokine production, induced by AdOPN treatment.
MATERIALS AND METHODS
Adenoviral construction, production, and titration
Mus musculus OPN-like sequence was obtained from NCBI GenBank (Accession No. X13986). The complementary DNA (cDNA) was purchased from Research Genetics, Inc.; OPN sequence was subcloned in pBSK vector with EcoRI and HindIII. For adenoviral vector construction OPN sequence was subcloned into pAdTrack-CMV, which was linearized with PmeI and cotransformed in Escherichia coli BJ5183 along with pAdeasy-1 vector, kanamycin resistant bacteria were selected, and recombination was confirmed by enzyme restriction with PacI. Finally, to obtain viral particles linearized plasmid was transfected to packaging HEK-293 cell line (ATCC No. CRL-1573) with lipofectamine (Invitrogen). Control vector encoding only green fluorescent protein (AdGFP) was reported previously. 15
To produce high titers of adenovirus (AdGFP and AdOPN), HEK-293 cells were infected at a multiplicity of infection of 0.1 in 150 mm cell culture dishes (Corning, Lowell, MA) and incubated at 37°C for 14 days. Then 3 mL of NP-40 (Invitrogen) was added and mixed for recovery of cells and supernatants, which were frozen at −20°C until posterior purification of adenoviral vectors. Purification was made precipitating the vectors with PEG 8000 (Sigma-Aldrich) and NaCl 2.5 M, followed by ultracentrifugation with CsCl gradient and filtration through a sepharose column with isosaline water.
For virus titration, six-well plates (Corning) were seeded with HEK-293 cells and growth until 90% confluency and then infected with the adenoviral vectors in serial dilutions of the stock in DMEM 2% SFB (Gibco/BRL, Grand Island, NY) for 1 h. Supernatant was discarded and monolayer cells were incubated at 37°C in noble agar 1.6% with MEM 2 × (1:1). Plaque forming units (PFU) were counted by fluorescent microscopy after 14 days of incubation.
Determination of production and biological activity of recombinant OPN coded by AdOPN
HEK-293 cells in 100 mm cell culture dishes (Corning) at 90% confluency were infected with AdGFP or AdOPN, supernatants were recovered, and total protein was quantified (BCA Protein Assay Kit; Thermo Fisher Scientific). OPN production by AdOPN was verified by western blot using specific antibodies and comparing with a positive control of recombinant OPN (Cat. No. 441-OP; R&D Systems).
A chemotaxis assay was performed to verify biological activity of OPN, for this 1 × 105 macrophages J744A.1 (ATCC TIB-67) were plated in the upper compartment of 24-well Transwell inserts with 8 μm pore size polycarbonate membranes (Costar; Corning) and allowed to attach for 1 h at 37°C. Then, in lower compartments the equivalent for 5 or 10 μg/mL of OPN from AdOPN supernatants was added. For negative control, wells with the same amount of supernatant of AdGFP were used. Positive controls consisted of recombinant macrophage colony stimulating factor (M-CSF) 1.32 nM (Cat. No. 315-02; Peprotech) or recombinant purified OPN (Cat. No. 441-OP; R&D Systems) 5 μg/mL in DMEM 10% SFB (Gibco/BRL). Migration was allowed for 8 h of incubation at 37°C. Nonmigrating cells were removed, and cells in the lower surface were fixed with methanol 70% and stained with Giemsa for quantification. Five random fields were chosen, and number of cells per field is reported from four different experiments.
Determination of gene therapy dosage
Healthy male 8-week-old BALB/c mice were anesthetized with sevoflurane vapors (Abbott Laboratories, IL), and adenoviral vectors (AdGFP or AdOPN) were administered in 100 μL of isotonic sterile endotoxin-free saline solution intratracheally, with a stainless-steel cannula (Thomas Scientific, Swedesboro, NJ). Three different doses were evaluated: low (5 × 106 PFU), medium (5 × 107 PFU), and high (5 × 108 PFU) to determine the best dosage for OPN expression and safety in lung. Groups of three mice per dose were euthanized by exsanguination under anesthesia with pentobarbital after 1, 7, and 21 days postadministration; right lungs were frozen immediately in liquid nitrogen and left lungs were perfused with absolute ethanol for gene expression and histological analysis, respectively.
Experimental model of progressive TB
We used the murine model of intratracheal (i.t.) infection described previously. 16 Briefly, the virulent clinical strain of Mtb CIBIN-99, which is an isolate from a patient with advanced pulmonary TB that is resistant to all the first-line antibiotics (MDR-TB), was cultured in Middlebrook 7H9 broth (Difco Laboratories, Detroit, MI). After 1 month of culture, Mtb was harvested and adjusted to 2.5 × 105 bacteria in 100 μL of phosphate-buffered saline (PBS), aliquoted and maintained at −70°C until use.
Pathogen-free male BALB/c mice (6–8 weeks age) were anesthetized with sevoflurane (Abbott Laboratories), and 2.5 × 105 bacteria suspended in 100 μL of sterile PBS with 0.02% of Tyloxapol were inoculated intratracheally (i.t.) using a rigid stainless-steel cannula. Infected mice were maintained in groups of five in cages fitted with microisolators connected to negative pressure in a P-3 biosecurity level facility.
To determine the OPN gene expression kinetics and cellular source, infected animals were randomly selected in groups of five and euthanized by exsanguination under anesthesia with pentobarbital at days 1, 3, 7, 14, 21, 28, 60, and 120 postinfection. Lungs were collected immediately in 2 mL cryotubes and frozen in liquid nitrogen for bacterial load determination and quantitative PCR or fixed by perfusion with absolute ethanol for conventional histology and immunohistochemistry.
Gene therapy and antibiotic administration in an experimental model of progressive TB
The experimental model of progressive TB used previously to determine the expression of OPN was used. At day 60 postinfection, when infected animals are in active late progressive disease, each mouse received for gene therapy one dose of 5 × 107 PFU of AdOPN suspended in 100 μL of isotonic sterile saline by intratracheal route as described above. Groups of six mice were euthanized by exsanguination under anesthesia with pentobarbital at days 14, 28, and 60 post-treatment; lungs were collected immediately in 2 mL cryotubes and frozen in liquid nitrogen for bacterial load determination and quantitative PCR or perfused with absolute ethanol for histological analysis. For control groups, infected mice received intratracheally recombinant adenoviruses expressing AdGFP at the same dosage (5 × 107 PFU) or only the vehicle (isotonic saline solution).
To assess the effect of AdOPN combined therapy with second line antibiotics, the next day after adenoviral administration, infected mice received daily second-line antibiotics by gavage in 100 μL of distilled water with 0.275 mg of moxifloxacin, 0.1375 mg of ethionamide, and 0.4125 mg of pyrazinamide, as well as intramuscular amikacin (0.4125 mg dissolved in 10 μL of sterile water). Control groups with the same number of animals that received AdGFP or the vehicle were treated with the same scheme of antibiotics. Groups of six mice per experimental group were euthanized as previously described at days 7, 14, 28, and 60 post-treatment. Two independent experiments were performed.
Determination of bacterial load in lungs
Frozen left lungs were disrupted using ceramic beads in tubes with 1 mL of PBS-Tween 80 0.05% and homogenized in the FastPrepR-24 (MP Biomedicals). Four consecutive logarithmic dilutions of each sample were prepared, and 10 μL was plated in duplicate on Bacto Middlebrook 7H10 agar (Difco BD, Sparks, MD) enriched with OADC (Difco). CFU were counted after 21 days of incubation at 37° C with 5% CO2.
Gene expression determination by quantitative PCR
Right lungs frozen in 1.5 mL cryotubes were homogenized using ceramic beads in the FastPrepR-24 (MP Biomedicals), and RNA extraction was performed using the RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. The RNA was quantified and evaluated by spectrophotometry (A260/280; EPOCH 2; BioTek), and 100 ng of total RNA was retrotranscribed with Omniscript Kit (Qiagen) to obtain the cDNA.
Real-Time PCR for gene expression was performed using the Real-Time PCR system 7500 (Applied Biosystems) along with the QuantiTect SYBR Green Mastermix Kit (Qiagen). Specific primers for targets were designed (Primer-Blast, NCBI) and synthetized (Invitrogen) as following:
Cycling conditions were as follows: initial denaturation at 95°C for 15 min, followed by 40 cycles at 95°C for 20 s, 60°C for 20 s, and 72°C for 30 s. Ct values were determined, and relative quantification was calculated by the 2(−ΔCt) method to the endogenous “housekeeping gene” (Rplp0).
Morphological, immunohistochemical, and immunofluorescent analysis of lungs
Right lungs of mice were perfused with absolute ethanol by endotracheal route and fixed for 24 h; parasagittal sections were dehydrated and embedded in paraffin. Sections of 4 μm were obtained and stained with hematoxylin and eosin. For granuloma and pneumonia quantification was used as an automated image analyzer (Leica Q500/W Image Analysis System; Milton Keynes, United Kingdom) in which total lung area, areas of pneumonia, and empty spaces were selected, and the percentage of surface area and size of granulomas in square microns were determined.
For immunohistochemistry, the same paraffin blocks were sectioned and deposited on slides covered with poly
For quantification, 10 random fields in areas of pneumonia of each sample were analyzed; total number of cells and percentage of immunostained cells were determined using an automated image analyzer (QWin Leica) to acquire images and ImageJ 1.52a software (National Institute of Health) for positive cell counting. Negative control for antibody recognition was taken as baseline to determine cell positivity.
For immunofluorescence, sections from paraffin blocks were sectioned and deposited on slides covered with poly
Images were obtained with LSM 710 Duo (Carl Zeiss, GmbH) at 40 × , considering negative control for autofluorescence.
Statistical analysis
All data were analyzed with GraphPad Prism 6 (Graph Pad Software, Inc., La Jolla, CA). Statistical significance was defined using two-way ANOVA test for comparing between experimental groups at a 95% confidence interval (p < 0.05).
Ethical statement
Animal studies were approved by the Institutional Ethics Committee of Animal Experimentation of the National Institute of Medical Sciences and Nutrition, Salvador Zubirán, following the guidelines of the Mexican National Regulations on Animal Care and Experimentation (NOM 062-ZOO-1999).
RESULTS
Kinetics of osteopontin expression and cellular sources during experimental pulmonary TB
Infection (i.t. route) of BALB/c mice with a high number of a virulent clinical isolate of M. tuberculosis resistant to all first-line antibiotics (MDR-TB strain) induces a progressive pulmonary disease, in a similar way than in BALB/c mice infected with the reference drug sensible strain H37 Rv. 17
During the first 14 days after the infection with this MDR-TB strain there is a control of bacillary growth, followed by an increase of bacterial loads (Fig. 1B) that lead to lung damage (progressive pneumonia) (Fig. 1C). OPN is a cytokine that is expressed at low levels in lung tissue under homeostatic conditions 18 ; we confirmed this observation detecting minimal OPN immunostaining in lungs from noninfected BALB/c mice (Supplementary Fig. S2A) in few epithelial cells. However, OPN increases its expression in several inflammatory process, like allergies, cancer, and infectious diseases 19 ; in the last case, this molecule induces a pro-inflammatory cytokine pattern that confers protection against intracellular bacterial infection. 20 Thus, it was important to determine the kinetic of OPN expression in this experimental model of pulmonary MDR-TB.
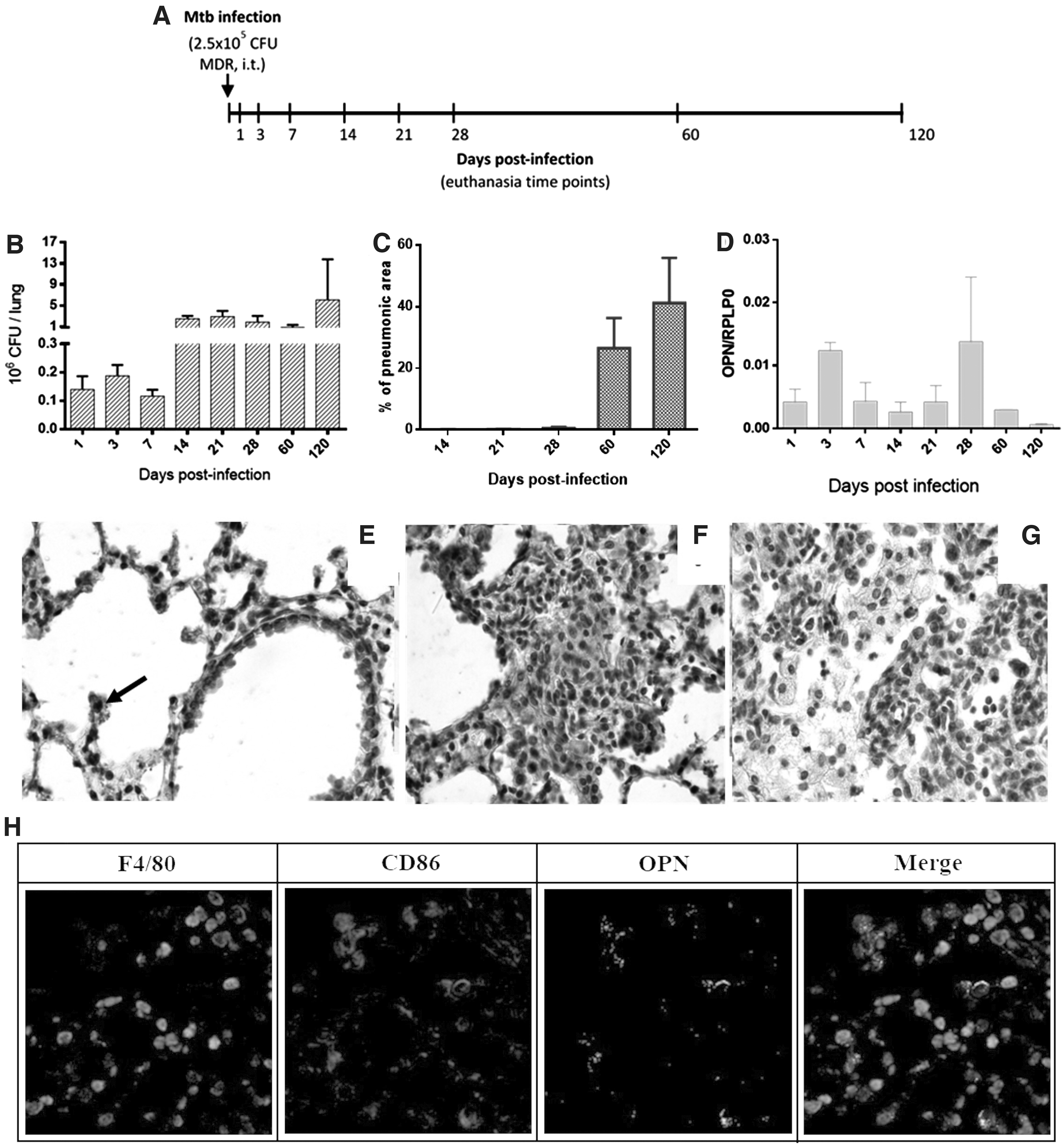
Kinetics of OPN expression during progressive pulmonary tuberculosis with an MDR-TB strain.
OPN gene expression was assessed by quantitative PCR (qPCR) and showed two peaks, at day 3 of infection when alveolar macrophages have been activated and at day 28 when pneumonia started its formation, followed by a decrease at days 60 and 120 after infection, being the later when the lowest expression was determined in coexistence with the highest number of CFU and affected pulmonary area with pneumonia (Fig. 1C, D).
Many alveolar and some bronchial epithelial cells showed OPN immunostaining during early infection (Fig. 1E). Activated macrophages located in alveolar spaces and in granulomas were the most common and stronger OPN immunostained cells at days 3 and 28 of infection (Fig. 1E, F), suggesting that this kind of cells are the most important source of this cytokine in this model of pulmonary TB; this was confirmed by immunofluorescence at day 28 postinfection with correlation of macrophage markers F4/80 and CD86 with OPN detection (Fig. 1H). In contrast, at day 60 and 120 postinfection OPN immunostaining was scarce, occasional foamy macrophages located at pneumonic areas showed scant staining, as well as in some epithelial cells and macrophages with compact cytoplasm (Fig. 1G). Thus, it seems that the production of OPN is low during late progressive disease and it is possible that inducing an increase of OPN production during this phase could improve protective immunity and bacterial killing; this possibility was tested by the administration of AdOPN.
Biological activity of recombinant OPN from AdOPN infection
To determine the therapeutic efficiency of AdOPN, we first evaluated OPN production in the supernatants of infected HEK-293 cells with AdOPN. Figure 2A shows a clear well-defined 65 kDa band of OPN detection in these supernatants by western blot using specific polyclonal antibodies against murine OPN, which was similar to the band detection of recombinant OPN that was used as control.

Production of recombinant OPN from AdOPN infected cells and determination of its best dose in mice.
Then, the biological activity of this protein produced by the HEK-293 cells infected with AdOPN was assessed by OPN chemotaxis to macrophages, using an assay with mouse macrophages in which through α4 and α9 integrins as receptors OPN induces chemotaxis. 21 Migration of J744A.1 macrophages was stimulated after 8 h of incubation, either with OPN from the supernatants (SN) of HEK-293 infected cells (OPN/SN) or with recombinant M-CSF or OPN as controls. The number of cells induced to migrate by the supernatants from HEK-293 infected cells with AdOPN was similar (media of 17.1 cells/field or 16.75 cells/field with 5 or 10 μg of OPN, respectively) than those incubated with recombinant OPN used as a control (18.6 cells/field). In contrast, the supernatants from AdGFP infected cells showed significant lesser migrated macrophages (3.8 or 3.5 less than OPN/SN5μg and recombinant OPN, respectively) (Fig. 2B). These results confirm the efficient biological activity of the protein encoded by the adenoviral vector.
Determination of AdOPN dose in healthy mice
BALB/c mice were administered intratracheally three different doses of adenoviral vectors: low (5 × 106 PFU), medium (5 × 107 PFU), and high (5 × 108 PFU) of AdOPN or the control vector AdGFP. As expected AdGFP also induced OPN expression but in a minor quantity than AdOPN, due to that viral infections have been demonstrated to induce OPN expression like in influenza and respiratory syncytial virus, 22,23 besides that adenoviral vector even nonreplicative still contains viral proteins and mutated genome that can induce a minor response. 24 Adenoviral vectors have a natural tropism to respiratory airway in epithelium and alveolar macrophages; we corroborate this by immunohistochemistry to OPN (Supplementary Fig. S2B) showing epithelial cells and alveolar macrophages expressing this cytokine, also GFP expression (flag gene in adenoviral vectors) was analyzed by immunofluorescence showing expression in the same cell type.
In comparison with AdGFP, gene expression of OPN was higher at day 1 post-treatment with AdOPN with low and medium doses (6.41- and 7.4-fold higher). For the following days the expression of OPN was still higher with low and medium doses of AdOPN, been at day 7 post-treatment 6.61 and 4.85 times higher in low and medium dose, respectively, and 21 days after treatment 3.81 and 2.66 times higher in comparison to AdGFP. In contrast, the viral higher dose had the same effect of expression with both vectors (AdGFP and AdOPN) at days 1 and 21 post-treatment (Fig. 2C).
Histological analysis did not show significant tissue damage in lung with any of the three doses at 1, 7, or 21 days after recombinant adenoviruses (AdOPN or AdGFP) were administered; only slight inflammatory lymphocytes that infiltrate around some blood vessels and airways were observed (Fig. 2D and Supplementary Fig. S1). Taken together all results, low and medium doses seem similar except that OPN expression has a lower variation among the three animals with medium dose, so we choose this dose to perform the therapeutic experiments.
Effect of AdOPN administration in a murine model of progressive pulmonary TB
A single dose of AdOPN was administered (i.t.) at day 60 postinfection with Mtb (MDR-TB strain) in BALB/c mice, along with the control groups AdGFP and saline solution (Fig. 3A). Treatment with AdOPN permitted total survival, while the survival showed by the control groups AdGFP and saline solution (SS) was 83% and 75%, respectively (Fig. 3B). As expected from survival rates, bacterial loads in lung were diminished when treated with AdOPN at 14, 28, and 60 days after treatment (Fig. 3C), being 4.15, 5.96, and 1.85 times lesser compared to the control group (AdGFP), in coexistence with lesser lung tissue damage, showing <10% of pneumonia at days 14 and 28 post-treatment in comparison with >40% exhibited by the control groups AdGFP and SS (14 and 9 times lower, respectively). At day 60 post-treatment pneumonia slightly increased in mice treated with AdOPN, but yet remains 2.5 and 3 times lower than AdGFP and SS group, respectively (Fig. 3D, E).
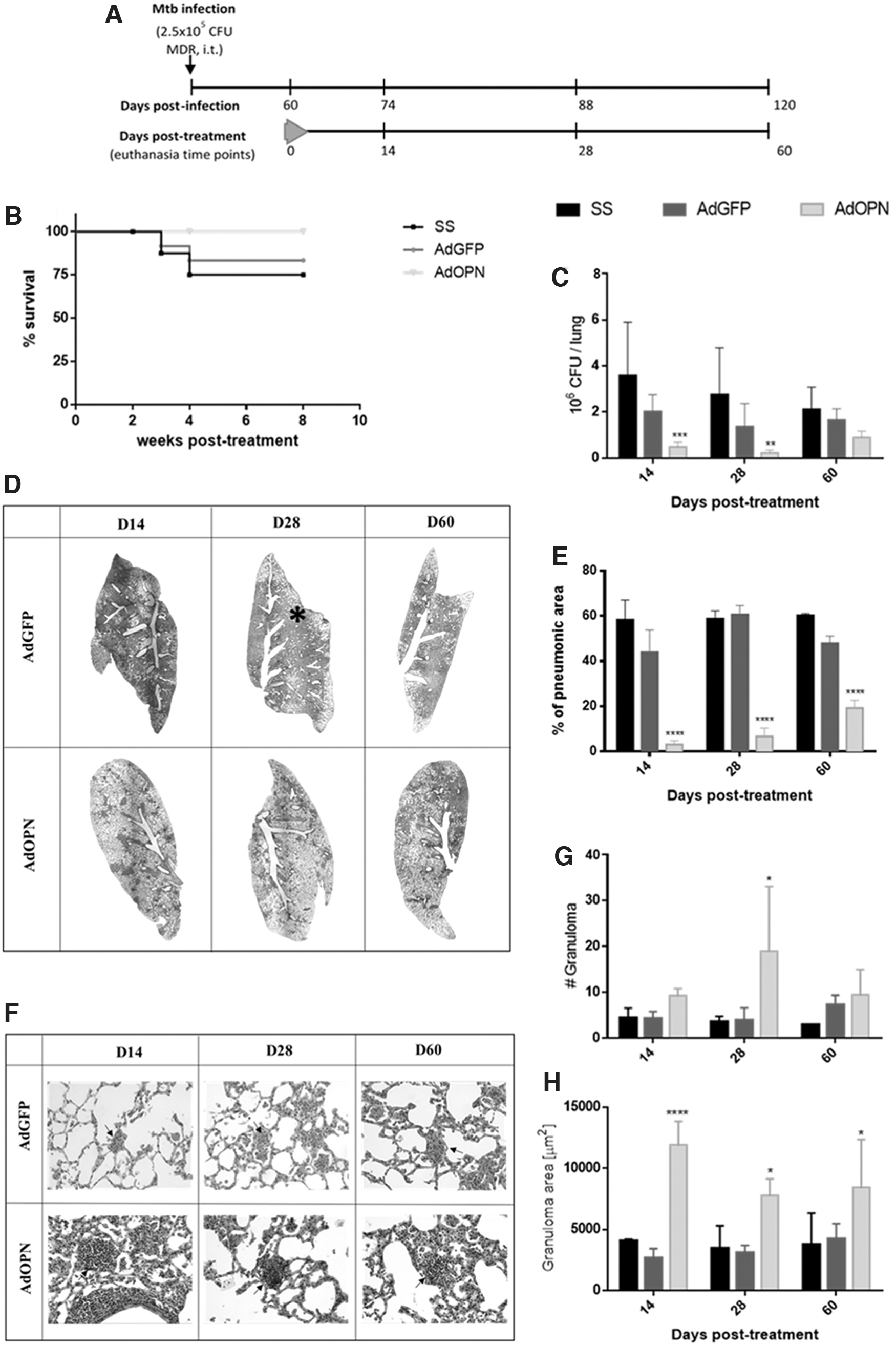
Therapeutic effect of one dose of AdOPN in pulmonary MDR-TB after 60 days of infection.
Immune regulation from gene therapy with OPN was observed in the number of granulomas (Fig. 3G); especially in day 28 post-treatment an average of 4 granulomas were counted per slide section in control group AdGFP, while 19 were observed in AdOPN group. These granulomas were also significantly bigger at the 3 evaluated time points, being 4.5 times bigger at day 14 and twice at 28 and 60 days post-treatment compared with control AdGFP. Thus, in comparison with SS control group, AdOPN induced an improved response of immune cells and containment of mycobacteria (Fig. 3F, H).
To assess more about the immune response induced by gene therapy with AdOPN, we seek for the expression of relevant cytokines in OPN pathway and TB. Gene expression in whole lung of IFN-γ and IL-12 by qPCR did not show differences among the groups, except at day 60 post-treatment; when the expression increased 21 times more for IFN-γ and IL-12 in comparison with AdGFP (Fig. 4A, B), expression for OPN was also assessed observing a tendency to increase along time post-treatment (Fig. 4C). Treatment with AdOPN reduced bacterial load and pneumonia as described before and this could contribute to not seeing a difference in cytokine expression in the whole lung at 14 and 28 days post-treatment; since there are less infiltrates of immune cells, so we seek for production of the same cytokines plus IL-17 in areas of pneumonia by immunohistochemistry.
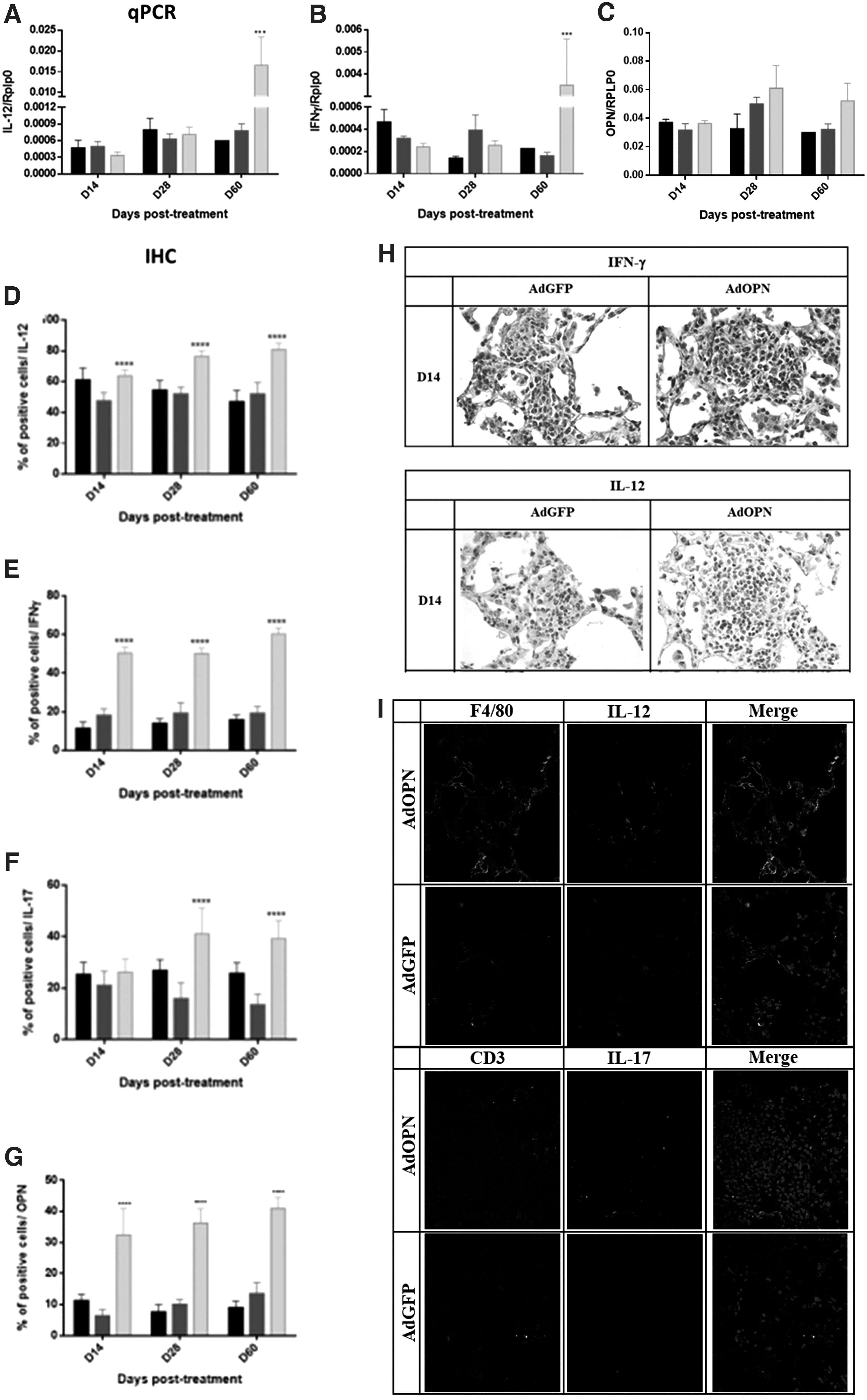
Cytokine expression induced by AdOPN gene therapy in pulmonary MDR-TB.
As shown in Fig. 4E, the percentage of cells in the pneumonic areas expressing IFN-γ was higher during the treatment with AdOPN, twice at 14 and 28 days post-treatment and threefold more at 60 days of treatment than the control group (AdGFP). Similar but lesser effect was seen in IL-12 expression (Fig. 4D). There was no difference in the percentage of IL-17 positive cells during the first 2 weeks of treatment, while at 28 and 60 days after treatment there was a higher percentage of IL-17 positive cells in areas of pneumonia in the group treated with AdOPN than in control AdGFP group (2.5 and 2.9 times higher) (Fig. 4F). As expected from AdOPN treatment, cells expressing OPN are higher than AdGFP and saline solution (Fig. 4G).
Taken together, immune regulation by gene therapy with AdOPN gave an effective activation and function of macrophages and lymphocytes at site of infection, as shown by the increase of IFN-γ, IL-12, and IL-17 immunostained cells in areas of pneumonia that prompts the formation of more and bigger granulomas with higher expression of IFN-γ and IL-12 (Fig. 4H) that contained and eliminated mycobacteria producing lesser bacterial load, reduced pneumonia, and in consequence showed higher mice survival. This higher production of protective cytokines by activated lymphocytes and macrophages was confirmed by immunofluorescence with confocal microscopy (Fig. 4I).
Administration of AdOPN with second line antibiotics in a murine model of progressive pulmonary TB
Immune regulation by gene therapy with AdOPN showed a better containment of mycobacterial infection in mice, so we further assess if combined therapy AdOPN in a single dose (i.t.) plus second line antibiotics could synergize and shorten chemotherapy (Fig. 5A).
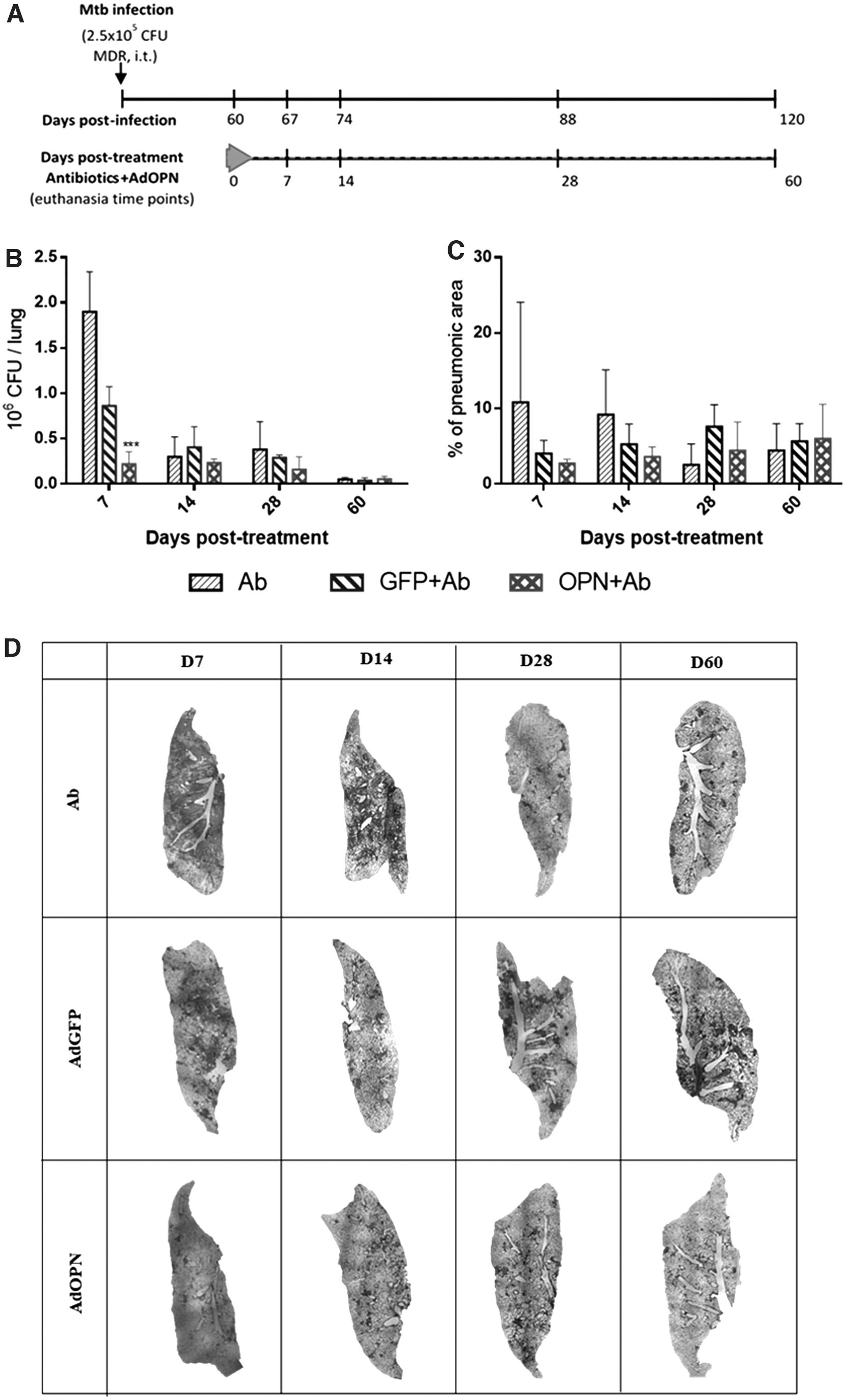
Effect of the combined therapy (second line antibiotics plus AdOPN gene therapy) in pulmonary MDR-TB.
As expected, total survival was seen in all treated groups (data not shown), since second line antibiotics are quite effective in reducing pulmonary bacterial loads as seen in Fig. 5B, particularly in the group that received only antibiotics (Ab) compared with the group without treatment (SS) (Fig. 3C). In the group treated with AdOPN and antibiotics (OPN+Ab), the bacterial load was four times lower at day 7 post-treatment compared to the control vector plus antibiotics (GFP+Ab). Then, a similar trend was observed at days 14, 28, and 60 post-treatment, although this was not statistically significant (Fig. 5B).
In comparison to Ab group, decreased bacterial load at day 7 post-treatment is higher in the combined AdOPN+Ab treated group (8.7 times lower). This more rapid bacterial elimination produced by the combined treatment suggested that it can shorten chemotherapy. Interestingly, AdGFP+Ab also showed higher bacterial elimination than the group treated only with antibiotics at this time point (two times lower bacterial load), suggesting that the production of cytokines induced by control AdGFP also contribute to an efficient bacterial elimination, but it was lower than induced by AdOPN (Fig. 3C).
Lung damage in all groups was below 10% (except for the group of Ab at day 7) during the whole course of the experiment; a nonsignificant trend to a lower percentage of pneumonia was observed (D7, D14, and D28 post-treatment) in the group of combined therapy with OPN+Ab (Fig. 5C), compared to GFP+Ab group. This trend is more notorious compared to the Ab group. Representative low power micrographs of the whole lung show this trend of better tissue architecture improvement in the AdOPN+Ab group (Fig. 5D).
DISCUSSION
TB is 1 of the 10 principal causes of death in developing countries and the leading cause of death by a single bacterial infectious agent in 2019. An estimate of 10 million people fell ill with TB and ∼1.2 million died worldwide. 1
Standard treatment for pulmonary TB involves 2 months of a quadruple antibiotic therapy with isoniazid (INH), rifampicin (RMP), ethambutol (EMB), and pyrazinamide (PZA) followed by 4 months of RMP and INH. 25 Due to this long time of treatment and adverse effects it is frequently abandoned, which commonly leads to mycobacterial drug resistance to one or several antibiotics of the standard treatment.
Drug resistance has become a growing problem for the treatment of TB, with an annual incidence of half million cases resistant to RMP (RR-TB), from which 78% were MDR-TB (resistant to RMP and INH). MDR-TB treatment is longer, taking from 18 to 20 months with the use of a fluoroquinolone like moxifloxacin or levofloxacin and an injectable drug such as amikacin, presenting more toxicity and high cost. 1,25 Hence, it is important to find new therapies that can reduce the time of treatment and toxicity to prevent drug resistance and treatment abandonment. In our study we seek to improve the immune response with the use of an adenoviral vector (Ad5) codifying OPN in an experimental mice model of pulmonary TB, in combination with MDR-TB antibiotic scheme with the aim to get a better and most rapid mycobacteria elimination.
OPN is a complex cytokine that was first discovered as a sialoprotein crucial in bone homeostasis. 26 However, several roles on the immune response have been associated to this molecule, such as chemotaxis of macrophages, neutrophils, NK cells, T cells, and dendritic cells through integrin receptors α4, α9, αv, α3 and CD44 and the production of MCP-1. 9,27,28 Maturation of dendritic cells is induced by OPN, driving to activation of T cells expressing IFN-g and IL-12 prompting a Th1 or Th17 polarization. 12,29 Interestingly, these immune profiles are decreased in TB late progressive disease that is associated with an increase of regulatory and tissue-repairing molecules, such as IL-4 produced by Th-2 cells and TGF-b and IL-10 produced by T regulatory cells and M2 macrophages in the site of infection. 4,30
Our results show that OPN encoded by adenoviruses in mice suffering late active TB by MDR-TB evoked an efficient control of disease progression, in comparison with tuberculosis mice that received the control adenovirus that codifies the GFP protein (AdGFP), and the effect of AdOPN was even better than mice that received only the vehicle saline solution. AdOPN treated mice showed the lesser bacterial load and pneumonia after 14 and 28 days of its administration. Notoriously, 60 days after treatment the effect of AdOPN started to decay with an increase of both bacterial load and pneumonia; this response was expected since only a single AdOPN dose was administrated and adenoviral vector Ad5 is a nonreplicative and nonintegrating virus, so upregulated expression of cytokine correlates with a slight reactivation of mycobacteria infection.
Immune cells expressing protective cytokines IL-12, IL-17, and IFN-γ during the treatment with AdOPN were strikingly upregulated. This observation was well demonstrated by the study of the production of protective cytokines in the pneumonic areas by immunohistochemistry and immunofluorescence, especially for IFN-γ and IL-12, which confirmed the efficient activity of OPN that increased protective immunity during late active MDR-TB, prompting a Th1 and Th17 profile in lung that favors the control and reduction of infection.
Another effect from gene therapy with OPN was in granuloma formation, a key structure in controlling and eliminating mycobacteria; in our murine TB model the size of granulomas is related with the control of the disease, larger granulomas are related with protection, while small granulomas are present during disease progression. 31 OPN gene therapy prompted bigger and more numerous granulomas, reflecting the role of OPN and its induced cytokines in the formation of these significant structures. Interestingly, OPN has been associated in previous reports with the formation of granulomas not only in TB but also with other granulomatous diseases like sarcoidosis and silicosis. 20,32,33
Antibiotics against TB is a long treatment that frequently produces gastrointestinal disturbs, neuritis, exanthema, and severe hepatic toxicity. 25,34 Thus, shortening the long treatment of chemotherapy could be an efficient way to prevent toxicity and lack of compliance. Immunotherapy based in recombinant adenoviruses can contribute to this, at least at the experimental level, as has been explored before with BALB/c mice infected with H37Rv strain, which is a reference drug-sensible mycobacterium strain. Gene therapy based in adenoviruses that encoded the pro-inflammatory cytokines IFN-γ, administrated at late progressive disease or IL-12 supplied 1 day before Mtb infection, produced significant bacilli elimination. IL-12 therapy also prevented transmission of Mtb to cohousing convenient healthy mice, and AdIFN-γ showed synergism with conventional chemotherapy that produced lower bacterial load when combined with first line antibiotics. 15,35
In a latent TB infection mice model, AdTNF-α therapy led to bigger granulomas and a better containment of mycobacteria after glucocorticoid reactivation. 36 Other molecules like antimicrobial peptides have also been tested by gene therapy or natural products with a reduction of bacterial loads and higher survival, in some cases improving antibiotic treatment. 17,37 Thus, the possibilities for a combined therapy have been explored before since immunotherapy by itself is not capable of eliminating the bacilli completely.
In the present study an MDR-TB mice model was used to explore a combined therapy as an option to improve time of treatment in cases of MDR mycobacteria infection, since the current treatment is much more complex and larger than chemotherapy for drug sensible TB. OPN immunotherapy plus a scheme of second line antibiotics (amikacin, moxifloxacin, ethionamide, and pyrazinamide) was capable of reducing bacterial load more efficiently than antibiotics alone during the first week of treatment, indicating a better elimination of bacteria at shorter times of treatment with a trend to a faster recovery of lung damage and pneumonia. Then, bacterial loads in later time points did not show a difference between groups, due to that second line antibiotics treatment is quite efficient.
Adenoviral vectors for gene therapy have been used widely, not only for immunotherapy in infection diseases but also for the treatment of cancer, allergies, some genetic diseases, and even for vaccination. 38 –41 Along time, different generations of adenoviral vectors have been produced to generate a lower humoral immune response to the vector, with a larger and safer expression of the protein of interest or even by changing cell targets and tropism of the vector. 42 –44 In the present work an Ad5 vector of first generation was used for OPN immunotherapy.
Interestingly, AdGFP as control group also improved the immune response in mice infected with MDR-TB, probably due to an immune recognition of adenoviral proteins and genes present in this vector. Indeed, adenoviral vectors have many advantages, such as high security (nonreplicating virus); they have low production cost and have a natural tropism to respiratory airway in bronchial epithelium and alveolar macrophages through the known receptor CAR (Coxsackievirus–adenovirus receptor), 39,43 being excellent vectors for gene expression in the lung and do not require many doses as in the case of recombinant protein administration. This low and few dose administration of recombinant adenovirus are also necessary to reduce the inflammatory response and the neutralizing effect of antibodies induced by its repetitive administration.
In summary, our results showed that a single dose of AdOPN was able to reactivate the immune protection in mice infected with MDR mycobacteria suffering late progressive disease, by increasing IL-12, IFN-g, and IL-17 expression leading to a higher response of Th1 and Th17 profile, which resulted in an increase of the number and size of granulomas that efficiently contained and eliminated mycobacteria, producing a significant decrease of bacterial load and tissue damage that permit total survival (Fig. 6). AdOPN plus second line antibiotics reduced the bacterial load more efficiently than antibiotics alone during the first week of treatment, suggesting a better elimination of bacteria at shorter times of treatment that could contribute to shorten the long treatment of this significant infectious disease.
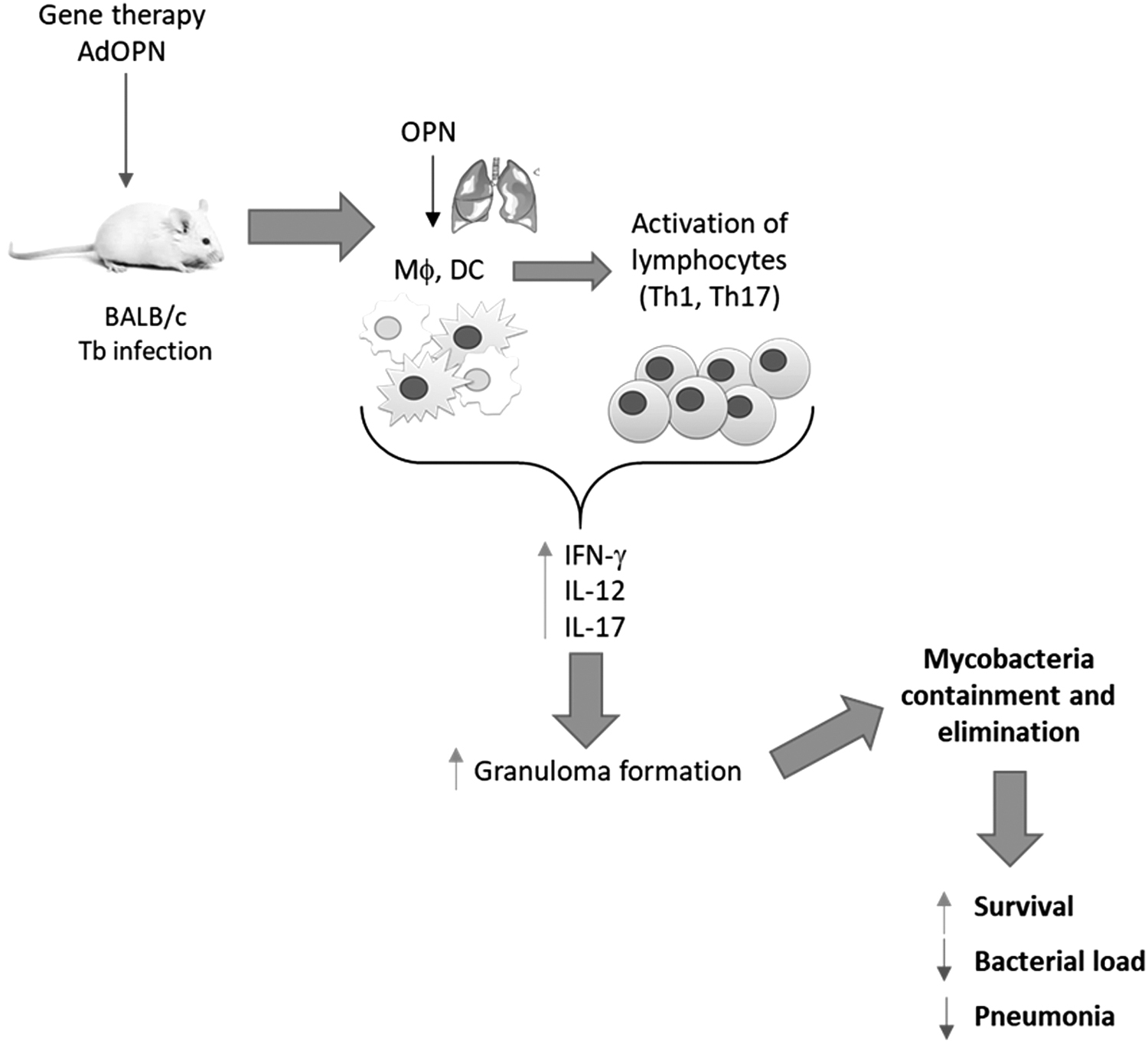
The intratracheal administration of a single dose of AdOPN at day 60 postinfection in mice with late active pulmonary MDR-TB produced a significant reactivation of protective immunity mediated by Th-1 and Th-17 cells, which induced more and bigger granulomas that improved the containment and elimination of mycobacteria producing higher survival and lesser tissue damage.
Footnotes
ACKNOWLEDGMENTS
S.H.B. is a doctoral student from Programa de Maestría y Doctorado en Ciencias Bioquímicas, Universidad Nacional Autónoma de México (UNAM) and received fellowship 385001 from CONACyT. Unidad de Aplicaciones Avanzadas en Microscopía (ADMiRA), INCan, RAI, UNAM, collaborate with confocal microscopy equipment.
AUTHOR DISCLOSURE
S.H.B., D.M.E., and R.H.P. contributed to the background work. S.H.B. and R.H.P. conceived experiments. S.H.B. performed, organized, and analyzed the results. S.H.B., O.R.E., and V.L. performed the experiments. D.M.E., J.B.P., and B.M. contributed with mice infection procedures. F.L.C. and D.M.E. designed the adenoviral vectors.
FUNDING INFORMATION
S.H.-B. is a PhD student from Programa de Doctorado en Ciencias Bioquimicas, Universidad Nacional Autónoma de México (UNAM) and received fellowship 385001 from Consejo Nacional de Ciencia y Tecnología (CONACyT). This study was supported by CONACyT project Fon Inst./58/2016.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.