
Abstract
Abnormal angiogenesis is associated with myriad human diseases, including proliferative diabetic retinopathy (PDR). Signaling transduction through phosphoinositide 3-kinases (PI3Ks) plays a critical role in angiogenesis. Herein, we showed that p110δ, the catalytic subunit of PI3Kδ, was highly expressed in pathological retinal vascular endothelial cells (ECs) in a mouse model of oxygen-induced retinopathy (OIR) and in fibrovascular membranes from patients with PDR. To explore novel intervention with PI3Kδ expression, we developed a recombinant dual adeno-associated viral (rAAV) system for delivering CRISPR/Cas9 in which Streptococcus pyogenes (Sp) Cas9 expression was driven by an endothelial specific promoter of the intercellular adhesion molecule 2 (pICAM2) to edit genomic Pik3cd, the gene encoding p110δ. We then demonstrated that infection of cultured mouse vascular ECs with the dual rAAV1s of rAAV1-pICAM2-SpCas9 and rAAV1-SpGuide targeting genomic Pik3cd resulted in 80% DNA insertion/deletion in the locus of genomic Pik3cd and 70% depletion of p110δ expression. Furthermore, we showed that in the mouse model of OIR editing retinal Pik3cd with the dual rAAV1s resulted in not only a significant decrease in p110δ expression, and Akt activation, but also a dramatic reduction in pathological retinal angiogenesis. These findings reveal that Pik3cd editing is a novel approach to treating abnormal retinal angiogenesis.
INTRODUCTION
Pathological angiogenesis,
The approval of gene therapy for RPE65-Leber congenital amaurosis (LCA2) 14 –17 has highlighted this strategy as a novel approach for the treatment of angiogenesis-associated eye diseases. Ocular diseases are particularly conducive to gene therapy, given the relative accessibility of the eye using established techniques that also allow noninvasive measurement of restoration of visual function to determine therapeutic efficacy. The eye is also immune-privileged, and adeno-associated virus (AAV)-mediated gene therapy of the retina has shown only limited local or systemic immune responses in the form of neutralizing antibodies to the AAV vector or T cell responses to the viral capsid. 17,18 The use of recombinant (r)AAV to express soluble Flt-1, the extracellular domain of VEGF receptor (VEGFR)1, has recently entered clinical trials 19,20 as a single injection approach to achieve long-term efficiency, in this case for the treatment of nAMD.
Targeted gene editing is possibly through induced double-stranded DNA breaks in the eukaryotic chromosome. By employing the microbial acquired immune system of CRISPR/Cas, the Cas (e.g., Cas9), a DNA (or RNA) endonuclease under the guidance of a single-guide RNA (sgRNA), binds to a DNA sequence complementary to the sgRNA spacer adjacent to a protospacer adjacent motif (PAM). Cas9 senses correct base-pairing, thus activating its RuvC and HNH nuclease domains to cleave target DNA strands, triggering genome editing. 18,21
Recently genome editing has been employed to deplete crucial genes to treat pathological conditions in a variety of animal models. For instance, VEGF depletion in retinal pigment epithelial (RPE) cells using rAAV-CRISPR/Cas9 has been explored to treat nAMD in a mouse model of laser-induced CNV. 22 Likewise, VEGFR2 depletion in vascular endothelial cells (ECs) has been harnessed to treat intraocular angiogenesis in mouse models of CNV and oxygen-induced retinopathy (OIR). 23 These two mouse models have been successfully used in the screening of currently used clinical anti-VEGF agents. 24,25 However, given that VEGF signaling is also required for neural cell survival, 23,26 an ideal approach would be to inhibit pathological angiogenesis without impacting on normal retinal function.
Phosphoinositide 3-kinases (PI3Ks) phosphorylate the 3′ position of the inositol head group of the phosphatidylinositol (PtdIns) membrane lipids, which serve as ligands and functional regulators of a range of proteins. One major downstream effector is the Akt serine/threonine kinase that plays a central part in cell survival, growth, metabolism, and angiogenesis. 27,28 PI3Ks include eight isoforms and can be divided into three classes, of which the class I PI3Ks are heterodimers consisting of a p110 catalytic subunit and a regulatory subunit. The class I PI3Ks engage in signaling downstream of tyrosine kinases, G protein-coupled receptors and monomeric small GTPases. Class I PI3K have four catalytic subunits in mammals, namely p110α, −β, −δ, and –γ encoded by the PIK3CA, −B, −D, and −G genes, respectively. 29
We have recently reported that PI3Kδ is highly expressed in vascular ECs in cultured human retinal microvasclar endothelial cells (HRECs), and that pharmacological or genetic inactivation of PI3Kδ suppresses pathological retinal angiogenesis in the mouse OIR model. 30 Given the hardly detectable expression of PI3Kδ in mouse photoreceptor cells, 30 it is likely that its activity in these cells is not essential. In addition, given that small molecular inhibitors for PI3Kδ (e.g., idelalisib) have a limited half-life in patients, and therefore most likely in vitreous, we thus applied a rAAV-based Pik3cd gene editing targeting to long-term dampen p110δ expression to experimentally treat the angiogenesis-related eye diseases, highlighting its potential as a novel approach for inhibiting angiogenesis-associated human diseases, including PDR.
METHODS
Reagents
Antibodies against p110α, p110β, p110δ, CD31, HIF1α, Akt, or phospho-Akt (S473) were from Cell Signaling Technology (Danvers, MA). The primary antibody against β-actin and secondary antibodies of horseradish peroxidase (HRP)-conjugated goat anti-rabbit immunoglobulin G (IgG) and anti-mouse IgG were from Santa Cruz Biotechnology (Santa Cruz, CA). Enhanced chemiluminescence substrate for detection of HRP was obtained from Thermo Fisher Scientific (Waltham, MA). Alexa fluorescence-594-conjugated mouse endothelial specific isolectin B4 (IB4) was from Life Technology (Grand Island, NY). High-fidelity Herculase II DNA polymerases were from Agilent Technologies (Santa Clara, CA).
Patient samples
Fibrovascular membranes (FVMs) from patients with PDR were prepared as described previously. 30,31 An ethics approval was obtained before the initiation of this project from the Vancouver Hospital and University of British Columbia Clinical Research Ethics Board. The University of British Columbia Clinical Research Ethics Board policies comply with the Tri Council Policy and the Good Clinical Practice Guidelines, which have their origins in the ethical principles in the Declaration of Helsinki. Written informed consent was obtained from patients.
DNA constructs
Three 20 nt target DNA sequences preceding a 5′-NGG PAM sequence at exon 5 in the C57BL/6J mouse genomic Pik3cd locus (NC_000070.6)
32
were selected for generating sgRNA for SpCas9 targets using the CRISPR design website (
Cell culture
C57BL/6 mouse primary brain microvascular endothelial cells (MVECs) and human retinal vascular endothelial cells (HRECs) were purchased from Cell Biologics (Chicago, IL) and Cell Systems (Kirkland, WA). These vascular ECs were cultured in the endothelial cell medium with a kit (Cell Applications, San Diego, CA), and their tissue culture dishes were precoated with gelatin-based coating solution (Cell Biologics). A human RPE cell line, ARPE-19, was purchased from American Type Culture Collection (Manassas, VA) and cultured in Dulbecco's Modified Eagle Medium (DMEM)/F12 supplemented with 10% fetal bovine serum (FBS); human embryonic kidney (HEK) 293T cells (HEK 293 containing SV40 T-antigen) from ATCC were cultured in high-glucose (4.5 g/L) DMEM supplemented with 10% FBS. All cells were cultured at 37°C in a humidified 5% CO2 atmosphere. 23
Production of AAV
Recombinant AAV2/1 (rAAV1) vectors were produced as described previously 23,36 in the Gene Transfer Vector Core in Schepens Eye Research Institute of Massachusetts Eye and Ear (Boston, MA). Briefly, triple transfection of AAV package plasmid (AAV2/1), transgene plasmid (pAAV-pICAM2-SpCas9: AAV-SpCas9, 23 pAAV-U6-Pik3cd-CMV-GFP: AAV-Pik3cd or pAAV-U6-lacZ-CMV-GFP: AAV-lacZ) and adenovirus helper plasmid were performed in a 10-layer hyper flask containing confluent HEK 293 cells. On day 3 post-transfection, the cells and culture medium were harvested and enzymatically treated with Benzonase (EMD Millipore). After high-speed centrifugation and filtration, the cell debris was cleared. The viral solution was concentrated by running through tangential flow filtration, and then loaded onto an iodixional gradient column. After one round of ultracentrifugation, the pure vectors were separated and extracted, then ran through an Amicon Ultra-Centrifugal Filter device (EMD Millipore) for desalting.
Both vectors were titrated by TaqMan PCR amplification (Applied Biosystems 7500, Life Technologies), with the primers and probes detecting the transgene. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was performed to check the purity of the vectors, which were named rAAV1-SpCas9, rAAV1-Pik3cd, and rAAV1-lacZ. 23
Western blotting analysis
MVECs at 90% confluency in a 24-well plate were lysed in 1 × sample buffer, which was diluted with extraction buffer (10 mM Tris-HCl, pH 7.4, 5 mM EDTA, 50 mM NaCl, 50 mM NaF, 1% Triton X-100, 20 μg/mL aprotinin, 2 mM Na3VO4, and 1 mM phenylmethylsulfonyl fluoride) from the 5 × protein sample buffer (25 mM EDTA [pH 7.0], 10% SDS), 500 mM dithiothreitol, 50% sucrose, 500 mM Tris-HCl (pH 6.8), and 0.5% bromophenol blue.
Mouse retinas were grounded in the extraction buffer and their lysates were clarified by centrifugation for 10 min at 4°C. After quantification with a BCA Assay Kit (Thermo Fisher Scientific), these clarified lysates were mixed with the 5 × protein sample buffer.
The protein samples were boiled for 5 min and then centrifuged for 5 min at 13,000 g. Proteins from the centrifuged and heated samples were separated by 10% SDS-PAGE, transferred to polyvinylidene difluoride membranes, and subjected to western blot analyses using the appropriate antibodies. Experiments were repeated at least three times. Signal intensity was determined by densitometry using NIH ImageJ software. 30
DNA sequencing
Transduced cells were pelleted for genomic DNA extraction using the QuickExtract DNA Extraction Solution (Epicenter, Chicago, IL) following the manufacturer's protocol. In brief, the pelleted cells were resuspended in the QuickExtract solution, vortexed for 15 s, incubated at 65°C for 6 min, vortexed for 15 s, and then incubated at 98°C for 5 min. The genomic region of ∼300 bp around the PAM was PCR amplified with high-fidelity Herculase II DNA polymerases (Agilent Technologies). The PCR primers for MVECs and mouse retinas were P33F (forward 5′-GAAGGGCTGGATGTCGCAC-3′) and P33R (reverse 5′-CATGGCTAGTCTTCCGGTGT-3′). The PCR products were separated in 2% agarose gel and purified with a gel extraction kit (Thermo Fisher Scientific) for Sanger DNA sequencing and next-generation sequencing. DNA sequencing was performed by the MGH DNA core facility. 30,37
A mouse model of OIR
C57BL/6J litters on postnatal day (P) 7 were exposed to 75% O2 until P12 in an oxygen chamber (Biospherix). Oxygen concentration was monitored daily using an O2 sensor (Advanced Instruments, GPR-20F). 24 At P12, the pups were anesthetized by intraperitoneal injection of 50 mg/kg ketamine hydrochloride and 10 mg/kg xylazine. During intravitreal injections, the eyelids of P12 pups were separated by incision. Pupils were dilated using a drop of 1% tropicamide and the eyes were treated with topical proparacaine anesthesia. Intravitreous injections were performed under a microsurgical microscope using glass pipettes with a diameter of ∼150 μm at the tip after the eye were punctured at the upper nasal limbus using a BD insulin syringe with the BD ultrafine needle. One microliter of both of rAAV1-SpCas9 with rAAV1-Pik3cd or rAAV1-lacZ (1 μL, 3.0 × 1012 vg/mL) was injected.
After the intravitreal injection, the eyes were treated with a triple antibiotic (Neo/Poly/Bac) ointment and kept in room air (21% O2). At P17, the mice were euthanized and retinas were carefully removed for western blot analysis or fixed in 3.7% paraformaldehyde.
Mice under 6 g of total body weight were excluded from the experiments. Each experiment was at least repeated three times in this OIR model. Retinal whole mounts were stained overnight at 4°C with the murine-specific EC marker IB4-Alexa 594 (red). 23 The images were taken with an EVOS FL Auto microscope.
All the animal experiments followed the guidelines of the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research. Investigators who conducted analysis were masked as to the treatment groups. All the mice were cared for by following the Animal Care and Use Committee (ACUC) protocol approved by the Institutional Animal Care and Use Committee at Schepens Eye Research Institute, and at Jinan University.
RNA sequencing
Retinas carefully isolated from mice intravitreally injected the dual rAAV1 of rAAV1-pICAM2-SpCas9 with rAAV1-Pik3cd or rAAV1-lacZ (1 μL, 3.0 × 1012 vg/mL) were sent to Novogene 38,39 for RNA sequencing analysis, whose report was summarized in the Supplemental Data. Briefly, RNA-sequencing was performed on the Illumina HiSeq2500 with the sequencing reagents and flow cells that provided up to 300 Gb of sequence information per flow cell. The quality of the total RNA was estimated using the Agilent 2100 Bioanalyzer. For messenger RNA (mRNA)-seq, 2 rounds of polyA+ selection were performed. We used the Stranded mRNA Library Generation Kits as per the manufacturer's instructions (Agilent Technologies). The complementary DNA (cDNA) libraries were quantitated using qPCR in a Roche LightCycler 480 with the Kapa Biosystems Kit for library quantitation (Kapa Biosystems, Woburn, MA) before cluster generation. Clusters were produced to yield ∼725–825 K clusters per mm2. Cluster density and quality were determined during the run after the first base addition parameters were assessed. Around 2 × 50 bp sequencing runs were used to align the cDNA sequences to the reference genome.
Quantification of vaso-obliteration and neovascularization
This was performed as described previously. 23 Briefly, retinal image was imported into Adobe Photoshop CS4, and the Polygonal Lasso tool was used to trace the vascular area of the entire retina. Once the vascular area was highlighted, the number of pixels was obtained. After selecting total retinal area, the lasso tool and the “subtract from selection” icon was used to selectively remove the vascularized retina leaving behind only the avascular area. Once the avascular region was selected, click the refresh icon again to obtain the number of pixels in the avascular area.
When analyzing neovascularization (NV), the original image was reopened. The magic wand tool was selected from the side tool panel on the left side of the screen. On the top tool panel, the tolerance to a level that will pick up NV was set while excluding normal vessels (beginning at 50). Regions of NV were selected by clicking on them with the magic wand tool. The areas of NV fluoresced more intensely than surrounding normal vessels. When neovessels were selected, the area of interest was zoomed in by holding the “Alt” key on the keyboard and scrolling up. When all NVs were selected and checked, the icon was refreshed and the total number of pixels were recorded by clicking in the NV area. 23
Immunofluorescence
Embedded frozen mouse eyeballs were prepared as described previously. 30,40 The eyeballs were embedded in O.C.T. on dry ice and sectioned. The slides were fixed in 3.7% formaldehyde/phosphate-buffered saline (PBS) for 10 min. Subsequently, the tissues and sections were preincubated with 5% normal goat serum in 0.3% Triton X-100/PBS for 20 min, incubated with IB4 (1:100 dilution) for 1 h. Following three washes with PBS, the slides were mounted with a mount medium containing 4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories Inc., Burlingame, CA) and photographed using an EVOS FL Auto microscope. 30
FVMs and eyeball sections on slides were fixed in 3.7% formaldehyde/PBS for 10 min. Subsequently, the sections and FVMs were preincubated with 5% normal goat serum in 0.3% Triton X-100/PBS for 30 min, incubated with primary antibodies against p110δ (1:200, rabbit) and CD31 (1:200, mouse) or a normal rabbit (or mouse) IgG overnight at 4°C. After three washes with 0.3% Triton X-100/PBS, the tissues and sections were incubated with fluorescently labeled secondary antibodies DyLight 549 (Vector Laboratories, Inc.) and DyLight 480 (1:300 dilution in blocking buffer) for 30 min. Following 3 washes with 0.3% Triton X-100/PBS, the slides were mounted with a mount medium containing DAPI and photographed under a fluorescence microscope. 23,40,41
Data preparation and associations of PIK3CD with Hif-1A
The transcriptome profiles and corresponding clinical information on PDR patients were downloaded from the Genomic Data of GEO (
Statistics
The data from three independent experiments were analyzed using an unpaired t-test between two groups, and one-way analysis of variance among more than two groups. For animal experiments the data from at least six mice were used for the statistical analysis. All data were analyzed using a masked procedure. p-Values <0.05 were considered statistically significant.
RESULTS
Hypoxia heightens p110δ expression in the mouse retinas
p110δ is expressed at low levels in cells other than leukocytes such as ECs, but its expression can be induced in ECs by tumor necrosis factor α. 43,44 Our previous findings have demonstrated that high glucose induces p110δ expression in cultured HRECs. 30 Based on the notion that diabetic hyperglycemia can cause retinal ischemia, leading to hypoxia, 45 we hypothesized that hypoxia might also be able to enhance p110δ expression in the retina. Indeed, western blot analysis of retinal lysates from P17 mice with or without experiencing OIR showed that hypoxia significantly promoted p110δ expression in the mouse retina without altering expression of p110α and p110β (Fig. 1A, B).

Hypoxia enhances p110δ expression in mouse retinas.
In addition, p110δ was expressed at low levels in mouse retinal cone cells and high levels in mouse macrophages (Fig. 1A, B), in agreement with our previous findings. 30 To reveal if p110δ was expressed in pathological ECs in vivo, we examined p110δ expression in mouse retinas of the OIR model at P17, a time when pathological angiogenesis reaches a peak in this model. 24 High p110δ expression was confirmed by immunofluorescence in pathological ECs (Fig. 2A–F); notably, its expression was hardly detectable in normal mouse retinal ECs (Supplementary Fig. S1).
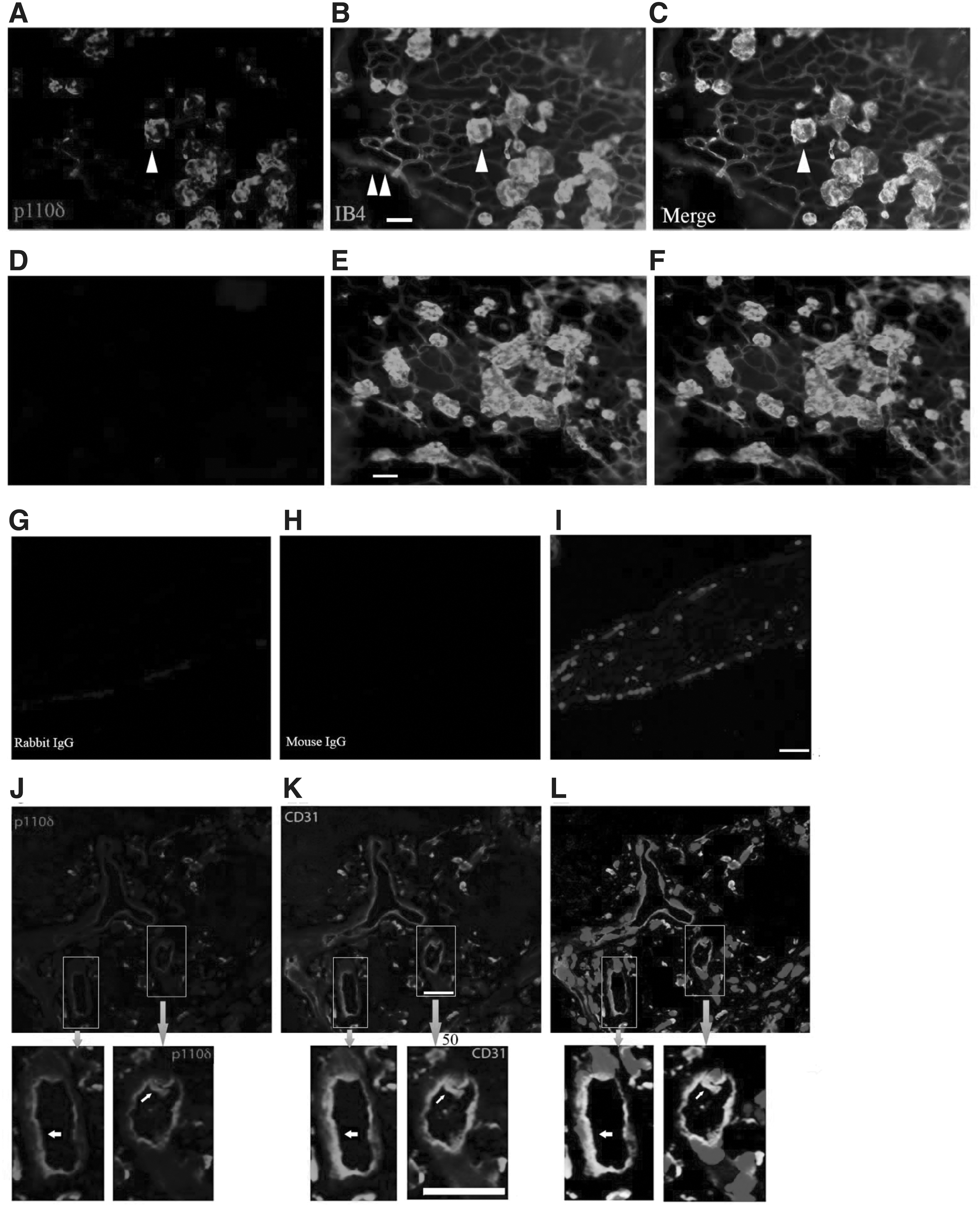
Expression of p110δ in pathological retinal ECs.
Furthermore, p110δ was also highly expressed in ECs of FVMs from patients with PDR (Fig. 2G–L), consistent with our previous findings. 30 Notably, pharmacological and genetic inactivation of PI3Kδ suppresses retinal pathological angiogenesis, 30 suggesting Pik3cd editing to impede p110δ expression as a novel approach for hindering pathological angiogenesis.
In vitro genomic Pik3cd editing
We previously established a dual rAAV system for Streptococcus pyogenes (Sp) Cas9 expression specifically in ECs. In this system, expression of SpCas9 is driven by the endothelial-specific promoter of the intercellular adhesion molecule 2 (pICAM2), namely pAAV-pICAM2-SpCas9, and in the SpGuide there is expression of green fluorescence protein (GFP) driven by a cytomegalovirus (CMV) promoter. 23 To edit mouse Pik3cd, we selected three gRNAs from mouse genomic loci and cloned them into the AAV-SpGuide vector by SapI, respectively, as shown previously. 23,33
We first analyzed the editing efficiency of the dual AAVs in cultured ECs. Primary C57BL/6 mouse brain MVECs were electroporated with plasmids of rAAV-pICAM2-SpCas9 in combination with plasmids of rAAV-mPK1, -mPK2, -mPK3 or -lacZ as a control. The mPK2-sgRNA to target mouse Pik3cd was found able to guide SpCas9 to cleave the genomic Pik3cd locus in MVECs, resulting in DNA mutations around the PAM (Fig. 3A, B) and depletion of p110δ in about 70% of the cells transduced with mPK2-sgRNA compared with those with the control lacZ-sgRNA (Fig. 3C, D). For in vivo studies, we next used the plasmids of rAAV-mPK2-sgRNA, rAAV-lacZ-sgRNA, and rAAV-pICAM2-SpCas9 to produce their rAAV1 viruses, respectively, given that rAAV1 transduces ECs with high efficiency. 23
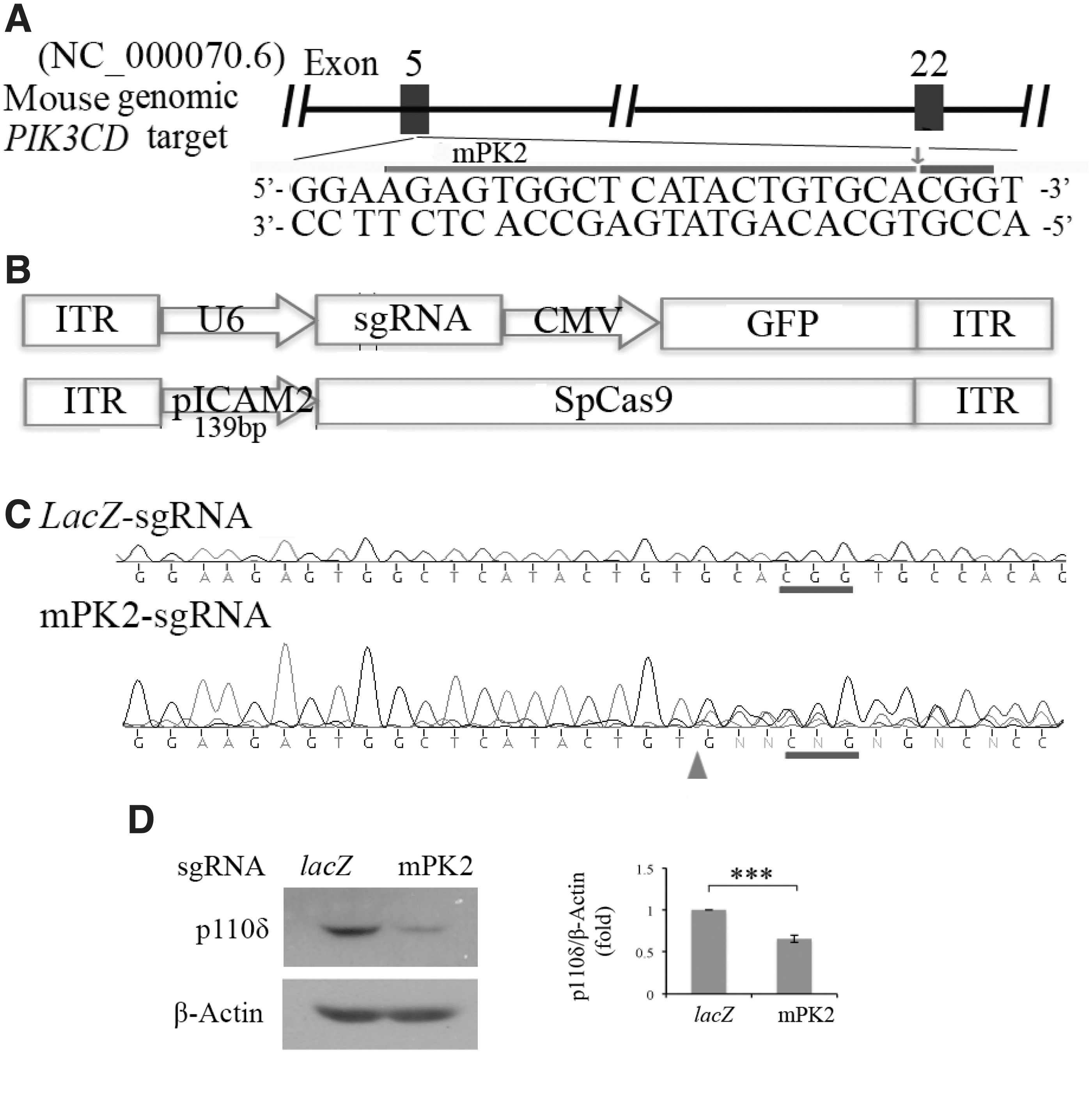
AAV-CRISPR/Cas9-mediated depletion of p110δ in vitro.
Editing Pik3cd suppresses abnormal retinal angiogenesis in a mouse model of OIR
Before testing the editing efficiency of the dual recombinant (r)AAV1s in vivo, we examined if the promoter of pICAM2 was specific for vascular ECs in vitro. As shown in Figure 4A, B, GFP expression driven by pICAM2 was hardly detectable in ARPE-19 cells, but it was detected in HRECs; in addition, GFP expression driven by the CMV promoter was very strong in ARPE-19 cells (Fig. 4C), demonstrating pICAM2 was specific for vascular ECs. We next evaluated whether rAAV1 could transduce ECs by intravitreal injection of rAAV1-CMV-GFP into mouse eyes at P12 experiencing the OIR model, followed by IB4 staining of frozen eye sections at P17 (Fig. 4D–G). These analyses showed that rAAV1 was able to transduce vascular ECs of neovascularization induced by hypoxia in the OIR, in agreement with our previous findings. 23
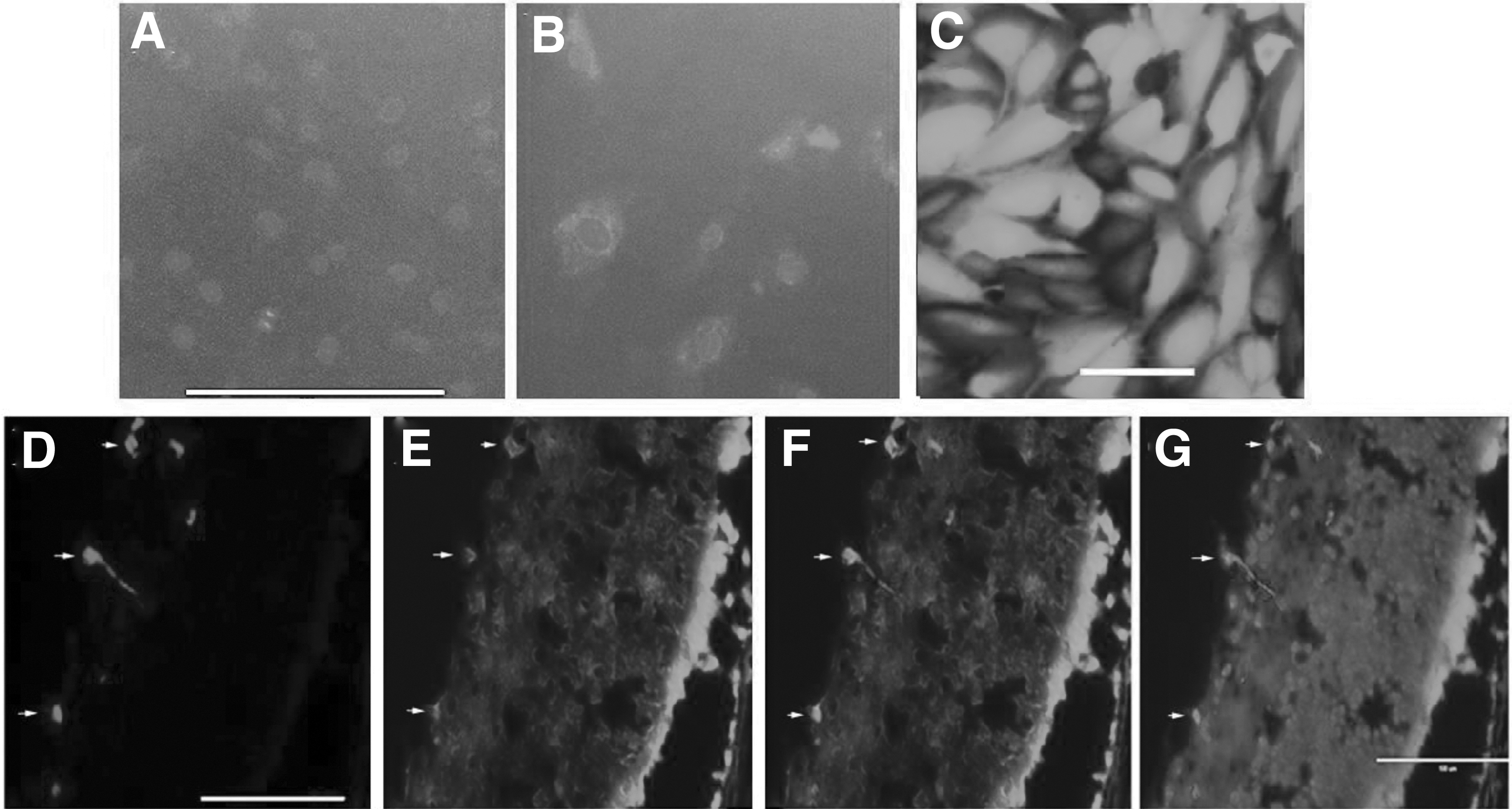
Transduction of ECs with rAAV1 in vitro and in vivo.
Subsequently, we intravitreally injected rAAV1-pICAM2-SpCas9 together with an equal amount of rAAV1-Pik3cd-sgRNA or lacZ-sgRNA (1 μL, 3.0 × 1012 vg/mL) into P12 mice in the OIR model, followed by analysis of the retinas at P17. IB4 staining of whole-mount retinas revealed a significant decrease in the number of preretinal tufts (reflective of pathological neovascularization) 24 (Fig. 5A, B; Supplementary Fig. S3) in the retinas from mice injected with Pik3cd-sgRNA compared with those injected with control lacZ-sgRNA, but the avascular areas between these groups were not significantly changed (Fig. 5A, C). Gene editing efficiency tested by next-generation sequencing of retinas showed a 2.05% insertion and 0.68% deletion with Pik3cd-sgRNA (Fig. 5D). Western blot analysis indicated there was 30 ± 0.5% reduction in p110δ expression in the retinas from mice injected with rAAV1-Pik3cd-sgRNA compared with those with rAAV1-lacZ-sgRNA with no significant changes in p110α and p110β expression (Fig. 5E, F).
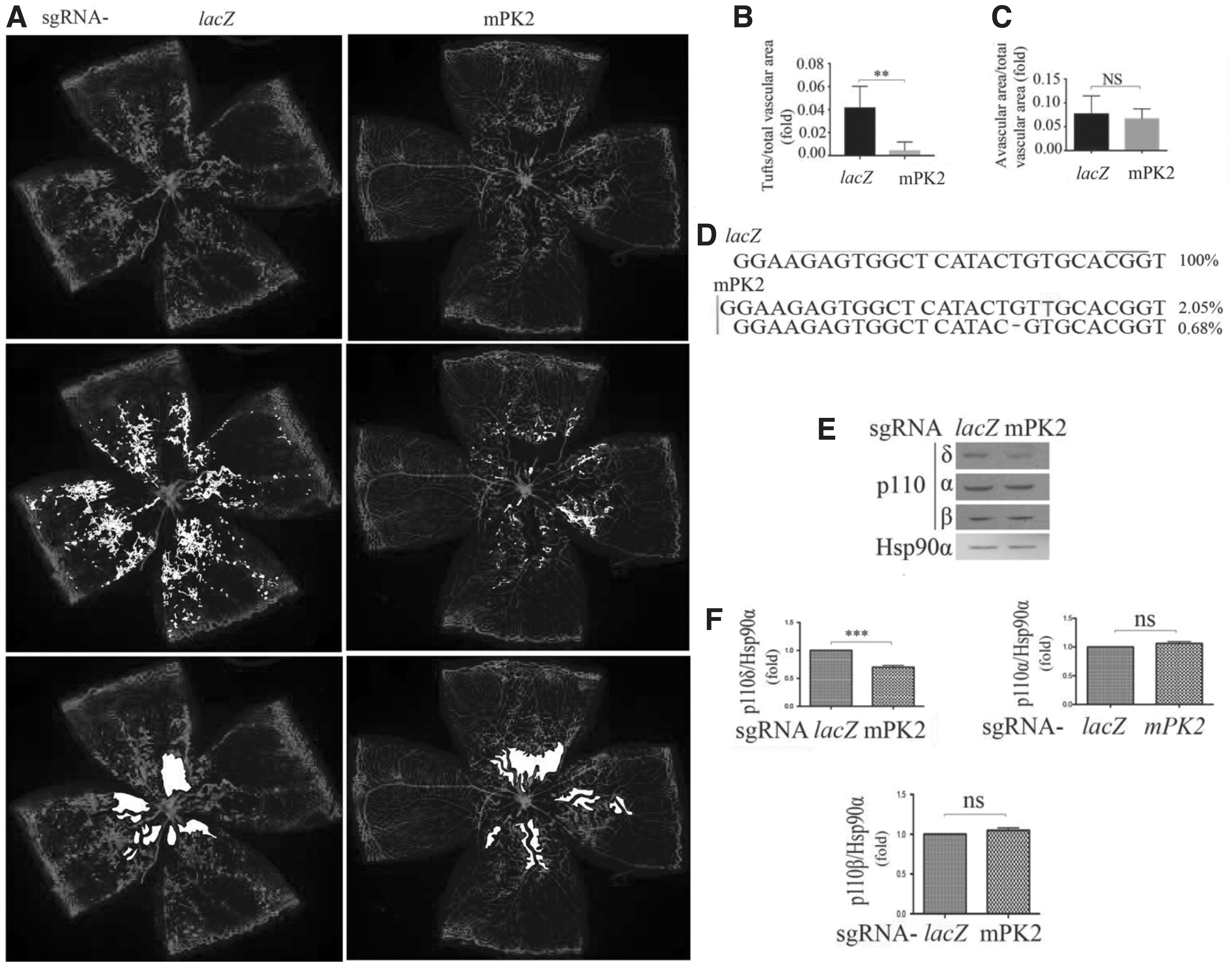
Editing genomic Pik3cd attenuates hypoxia-induced angiogenesis.
Taken together, these data in Figure 5 demonstrate that Pik3cd editing in vascular ECs reduced p110δ expression and suppressed hypoxia-induced pathological neovascularization in the mouse model of OIR.
Next, RNA sequencing was used to examine which signaling pathways were impacted after genomic editing of endothelial p110δ in the retinas from P17 mice experiencing OIR. As shown in Figure 6A and Supplementary Figures S4–S9, Pik3cd editing affected 20 signaling pathways, of which PI3K/Akt signaling was downregulated as expected, but we also found that HIF1 signaling was the strongest downregulated candidate among these signaling pathways (Fig. 6A). Western blot analysis confirmed that there was a significant decrease in Akt activation and HIF1α expression after Pik3cd editing in retinal vascular ECs in the OIR mice (Fig. 6B), suggesting that Pik3cd editing resulted in the suppression of pathological retinal angiogenesis by reducing HIF1α expression through blocking hypoxia-induced Akt activation.
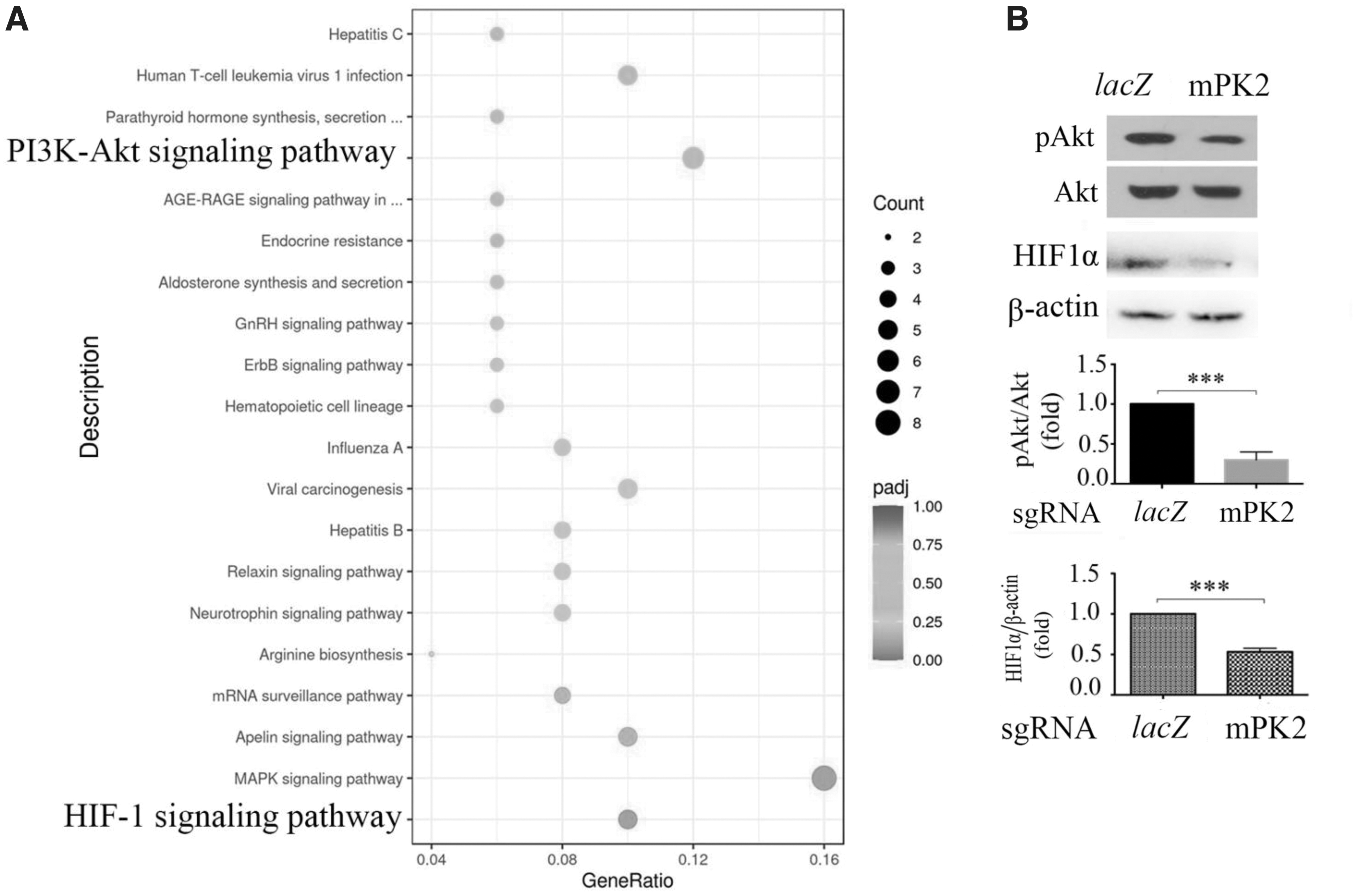
Editing genomic Pik3cd lessens hypoxia-induced Akt activation and HIF1αexpression.
DISCUSSION
In this study, we report that Pik3cd editing with a dual rAAV1 system to deliver SpCas9 and its gRNA into mouse ECs suppresses p110δ expression and attenuates pathological angiogenesis in the mouse model of OIR. These data are in line with a previous study demonstrating that inactivation of PI3Kδ using pharmacological or genetic approaches inhibits pathological angiogenesis in the OIR model. 30 Importantly, in this study we uncovered that hypoxia enhances p110δ expression in vascular ECs (Figs. 1 and 2), and the purpose of this research was to deplete expression of p110δ in these ECs to prevent abnormal neovascularization.
In Figure 5, there are only about 2.7% of insertions and deletions around the PAM, which still led to significant suppression of hypoxia-induced retinal pathological angiogenesis, consistent with our similar findings for VEGFR2. 23 This could be explained by the possibility that rAAV1 preferentially infects pathological ECs, which constitute only a small fraction of whole retinal genomic DNA; however, the specific receptor(s) that rAAV146 prefer to engage for rAAV1 to enter pathological ECs remain elusive and this question is worth further investigation. This treatment was also found to give rise to biological effects already only 5 days after rAAV1 injection. This could be explained by our observations (Supplementary Fig. 2) that it is possible to detect AAV1 infection in cultured ECs within 6 h in vitro and vascular ECs within 24 h in vivo.
Upon intravitreal injection of AAV1 at P12, that is, when the mice just came out of the oxygen chamber, the abnormal ECs had not grown yet, but when these began to expand the next day, the rAAV1s were already present for infection, allowing to induce a decrease in expression of p110δ to suppress hypoxia-induced Akt activation and pathological ECs expansion.
Given the instability of HIF1α, which is regulated primarily by stabilization of the expressed protein, the western blot indicates HIF1α protein level (Fig. 6) may not be a direct indicator of HIF1α expression. Intriguingly, analysis of the transcriptome data, which were downloaded from the Genomic Data of GEO (

Diagram of a potential mechanism of PI3Kδ in pathological angiogenesis.
PI3Ks are major intracellular lipid enzymes, which transduce extracellular signals to the inside of cells. 29 Akt is a major effector of PI3Ks and plays a central role in angiogenesis. 27 Our previous findings demonstrated that PI3Kδ in vascular ECs mediates Akt activation downstream of multiple angiogenesis-enhancing factors, including VEGF, bFGF, and EGF. Hypoxia can induce production of these proangiogenic factors. 30 In line with these data, Pik3cd editing can suppress activation of Akt induced by multiple growth factors in response to hypoxia. In addition, PI3Kδ expression is extremely low in mouse photoreceptors compared with ECs. 30 In this way, anti-PI3Kδ is superior to anti-VEGF to only inhibit pathological angiogenesis. Taken together, editing Pik3cd is a novel approach with high potential to treat pathological angiogenesis, such as PDR, ROP, and nAMD.
A major disadvantage of this genome editing strategy is the generation of genomic alterations such as insertions and deletions. Generation of a kinase-inactive mutant of p110δD910A by prime editing 48 might therefore be a better approach. In addition, kinase-dead Cas9 49 could also be used to target and functionally disable the Pik3cd promoter. These potential strategies are currently being actively explored in our laboratory.
AAV-based gene therapy is expected to be most advantageous in the treatment of inherited retinal diseases, including inherited retinal degenerative disease. 17,18,20 In this study, we used this AAV approach to treat acquired angiogenesis-related ocular disease, aiming to decrease the frequency of intravitreal administration of anti-VEGF agents and to treat those patients who suffer from these eye diseases but do not respond to the anti-VEGF therapy. Our dual AAVs with an EC-specific promoter pICAM2 to drive SpCas9 expression is not expected to affect other cells, resulting in less potential side effects in the eye. However, off-target effects associated with prolonged Cas9 expression will need to be monitored. Overall, our results provide evidence for Pik3cd editing as a novel approach to prevent abnormal retinal angiogenesis.
Footnotes
ACKNOWLEDGMENTS
Dr. Hetian Lei is the guarantor of this work, and has full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis. Authors thank Dr. Joanne A Matsubara of the University of British Columbia, Canada for providing samples of FVMs from patients with PDR.
AUTHORs' CONTRIBUTIONS
W.W., G.M., H.Q., L.D., F.C., Y.W., X.G., and X.M. performed the experiments and analyzed the results. J.C. and J.A.M. provided patient samples. B.V., X.Y., and G.Z. revised the article. S.Z. and H.L. conceived the experiments, analyzed the data, and wrote the article.
DATA AND RESOURCE AVAILABILITY
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request. The [RESOURCE] generated during and/or analyzed during the current study is available from the corresponding author upon reasonable request.
AUTHOR DISCLOSURE
H.L. has a patient application of the dual AAV vectors related to this article, and B.V. is a consultant for iOnctura (Geneva, Switzerland), Venthera (Palo Alto. USA), Olema Pharmaceutical (San Francisco, USA), as well as has received speaker fees from Gilead (Foster City, USA). All other authors declare no conflicts of interest.
SUPPLEMENTARY MATERIAL
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.