
Abstract
Natural airway defenses currently impede the efficacy of viral vector-mediated airway gene therapy. Conditioning airways before vector delivery can disrupt these barriers, improving viral vector access to target receptors and airway stem cells. This study aimed to assess and quantify the in vivo histological and gene transfer effects of physical perturbation devices to identify effective conditioning approaches. A range of flexible wire baskets with varying configurations, a Brush, biopsy forceps, and a balloon catheter were examined. We first evaluated the histological effects of physical perturbation devices in rat tracheas that were excised 10 min after conditioning. Based on the histological findings, a selection of devices was used to condition rat tracheas in vivo before delivering a lentiviral vector containing the LacZ reporter gene. After 7 days, excised tracheas were X-gal processed and examined en face to quantify the area of LacZ staining. Histological observations 10 min after conditioning found that physical perturbation dislodged cells from the basement membrane to varying degrees, with some producing significant levels of epithelial cell removal. When a subset of devices was assessed for their ability to enhance gene transfer, only the NGage® wire basket (Cook Medical) produced a significant increase in the proportion of X-gal-stained area when compared with unconditioned tracheas (eightfold, p = 0.00025). These results suggest that a range of factors contribute to perturbation-enhanced gene transfer. Overall, this study supports existing evidence that physical perturbation can assist airway gene transfer and will help to identify the characteristics of an effective device for airway gene therapy.
INTRODUCTION
Gene addition therapy holds the potential to treat lung diseases, such as cystic fibrosis (CF), by inserting a correct gene copy into airway cells. Delivery vectors, such as the vesicular stomatitis virus G-pseudotyped (VSV-G) lentiviral (LV) vector, currently exhibit modest transduction levels in airways, 1 –3 regardless of the vector titer or volume delivered. 4 This is partly because natural barriers that protect airways against foreign particles also hinder vector access to target receptors. 5 Viral vectors do not efficiently penetrate viscous CF mucus, 6 –8 so trapped vector particles may be removed from the lungs by mucociliary clearance before transduction. 5 Tight junctions between airway epithelial cells also obstruct LV vector access to basolateral VSV-G receptors, 9 and basal cells. 10 It is advantageous to transduce basal cells, which function as airway stem cells, 11 because proliferation of basal cells containing an integrated transgene produces gene-corrected daughter cells for widespread, long-term gene expression. 12 Conditioning the airways before vector administration can help to overcome these epithelial surface barriers and make airway cells more susceptible to transduction.
Physical perturbation is a relatively underexplored conditioning method that uses a device to abrade or disrupt the airway epithelium before vector delivery. 13 The degree of perturbation from different devices ranges in severity from mild and intermittent disruption of epithelial integrity to complete stripping of cells from the basement membrane. 14,15 Early proof-of-principle mouse studies for gene vector delivery used forceps to externally compress, 16 or directly scrape tracheas 17 before adenoviral vector delivery, which led to increased transgene expression in the disrupted regions. Perturbation of rabbit tracheas using a bronchial cytology brush resulted in transduction of only 0.01% of epithelial cells using a retroviral vector, although this represented an increase over unconditioned tracheas. 18 Recently, abrasion of rat tracheas using a flexible wire basket (NCircle®; Cook Medical) increased the area of LV-LacZ reporter gene staining by 1,000-fold compared with unconditioned tracheas. 19
While physical perturbation has shown promising proof of principle, the nonuniform and varied transduction levels obtained in previous studies highlights the need to explore different devices and techniques to optimize the consistency and efficacy of the conditioning method for improved airway gene transfer. 19 Furthermore, the histological effects produced by different devices have not been characterized or compared; such studies could help to elucidate the relationship between epithelial disturbance and vector-mediated gene transfer levels.
This work compared the effects of different physical perturbation devices in vivo in rat tracheas, aiming to: (1) characterize the histological effects and quantify the degree of epithelial removal produced by different physical perturbation devices and (2) determine which device is the most effective for improving in vivo gene transfer by measuring the area of LacZ staining, as well as observing the extent of epithelium repair and basal cell transduction. We hypothesized that greater epithelium disruption with a physical perturbation device would increase gene transfer to epithelial cells and result in a greater area of LacZ staining over the unconditioned control. 19 This research contributes to the ongoing optimization of physical perturbation to improve the efficacy of viral vector-mediated gene therapy for lung disease.
MATERIALS AND METHODS
Animals
Animal studies were approved by the University of Adelaide Animal Ethics Committee under applications, M-2017-056 and M-2019-061 and complied with the National Health and Research Council (Australia) Code of Practice for Animal Care in Research and Training (2013).
Male and female Sprague Dawley rats were randomly assigned to experimental groups. Adult wild-type or heterozygous littermates >10 weeks age from our CF rat colony were used for histological assessment. 20 Wild-type rats >8 weeks of age were used for gene transfer assessment.
Histological assessment
We first considered whether conditioning methods resulted in notable histological effects using small sample sizes to provide proof of concept. Tracheas were nonintubated (n = 3, rats were neither intubated nor conditioned), unconditioned (n = 3, rats were intubated but not conditioned), or conditioned with a physical perturbation device after intubation (n = 3 per device) (Table 1).
Physical perturbation devices used as in vivo conditioning methods to determine their histological effects
Existing small endoscopy tools and a dental brush that were appropriate sizes to fit within adult rat tracheas were repurposed as perturbation devices for this exploratory investigation. Techniques were developed based on the characteristics and capabilities of individual devices. Scale bars = 5 mm.
PBS, phosphate-buffered saline.
Intubation and physical perturbation
Rats were anesthetized with a mixture of medetomidine (0.4 mg/kg; Ilium) and ketamine (60 mg/kg; Ceva) by intraperitoneal injection and intubated with a 16-gauge intravenous cannula to function as an endotracheal (ET) tube to maximize tracheal access. 19,21 Anesthetized rats were kept supine on a heating mat throughout all procedures. A device (Table 1) was inserted through the ET tube such that it extended 25–30 mm out of the cannula to reach the carina and was repeatedly moved along the trachea to and from the distal end of the ET tube (see detailed techniques in Table 1). 19 To ascertain device tolerability, intubated rats remained supine for 10 min after physical perturbation with breathing continually monitored. 19
Histology
At 10 min post-conditioning, ET tubes were removed, and anesthetized rats were humanely killed by intracardiac injection of 100 μL of sodium pentobarbitone (325 mg/mL; Virbac). Nonintubated rats were humanely killed by carbon dioxide asphyxiation. Tracheas were fixed in 10% neutral buffered formalin, embedded in paraffin wax, and sectioned at 5 μm before undergoing Hematoxylin and Eosin (H&E) staining. Slides were blinded before analysis. Images were captured using a Nikon Eclipse E400 microscope with a DS-Fi2-U3 camera, and NIS-elements D software (version 5.20.00; Nikon).
Quantification of epithelium removal by devices
Tracheal images were stitched together using Image Composite Editor (version 2.0.3; Microsoft). The length of the full trachea (dorsal and ventral sides) and sections of removed epithelium were measured from three representative H&E-stained trachea from each rat using ImageJ (version 1.53j) 22 and averaged. Epithelial removal was defined as sections that consisted of one cell layer of flattened cells as well as regions without any cells and was calculated as a proportion of the total trachea length. A subset of the perturbation devices was subsequently investigated for their ability to enhance LV-mediated gene transfer.
Gene transfer assessment
For the gene transfer assessment, a VSV-G pseudotyped LV-EF1α-3xFLAG-nlsLacZ-eGFP (LV-LacZ) vector was produced and titered by the Gene Silencing and Expression Facility at the Robinson Research Institute (The University of Adelaide). The Brush, NCircle®, NCompass®, NGage®, and 11582M Stone Extractor devices were tested, along with an unconditioned control. Each device was tested on n = 3–6 rats.
LV vector delivery
Rats were anesthetized and conditioned as above. Subsequent procedures were performed in a biosafety cabinet. At 10 min after physical perturbation, two 25 μL aliquots of LV-LacZ vector (5.2 × 108 TU/mL) were consecutively delivered through the ET tube through a gel-loading tip (Thermo Scientific™ QSP #010-Q) that was extended with plastic tubing so it protruded ∼2 mm past the ET tube tip into the trachea, as per our standard delivery procedure. 19 Unconditioned and intubated rats also received this vector dose. At 10 min after vector delivery, ET tubes were removed, and conditioned rats received Meloxicam (1 mg/kg; Ilium) through subcutaneous injection to relieve potential discomfort or pain caused by the perturbation procedure. Anesthesia was reversed for all rats through intraperitoneal injection of Atipamezole (1 mg/kg; Zoetis) and rats recovered under observation on a heating mat.
LacZ staining assessment
Animals were humanely killed by carbon dioxide asphyxiation 7 days post-vector delivery. Tracheas and lungs were harvested and fixed in 4% paraformaldehyde before X-gal staining for LacZ. 1 Tracheas were halved along the dorsal and ventral surfaces and held open with minutien pins on a SYLGARD® tray containing 70% ethanol for en face imaging using a Nikon SMZ1500 stereo microscope with a DigiLite 3.0MP Camera and TCapture software (Tucsen Photonics). Images were stitched together using Image Composite Editor before digitally quantifying the area of en face LacZ staining using a custom-written MATLAB script. 19 The LacZ-stained area was expressed as a proportion of the total tracheal area (from both halves). LacZ-stained tracheas were embedded in paraffin wax and sectioned at 5 μm. Slides were stained with H&E or used for basal cell immunohistochemistry using anti-cytokeratin 5 antibody (ab52635; Abcam) as previously described. 19
Statistical analyses
Statistical analyses were performed in R. 23 The length of epithelial removal was analyzed using a one-way analysis of variance followed by Dunnett's test to determine significance when compared with the unconditioned group; this control was selected as all conditioned rats were also intubated. As parametric assumptions were not met, Kruskal–Wallis and Dunn's test with the Bonferroni correction was used to determine statistical significance for en face LacZ-stained area data. Significance was defined as p < 0.05. Graphical and in-text results are expressed as mean ± standard error of the mean.
RESULTS
Histological effects of physical perturbation on airway epithelium in vivo
In nonintubated and unconditioned but intubated rats, the epithelium of the distal trachea appeared histologically normal, and consisted of tightly packed ciliated and nonciliated pseudostratified columnar epithelial cells (Fig. 1A) with short sections of cuboidal epithelial cells primarily over the cartilage rings (not shown). Cells exhibiting basal cell morphology were attached to the basement membrane (Fig. 1A). Intermittent sections of flattened or removed epithelium were occasionally observed along the trachea in all controls, likely due to harvesting or sectioning damage (Figs. 1C, D, and Fig. 2).
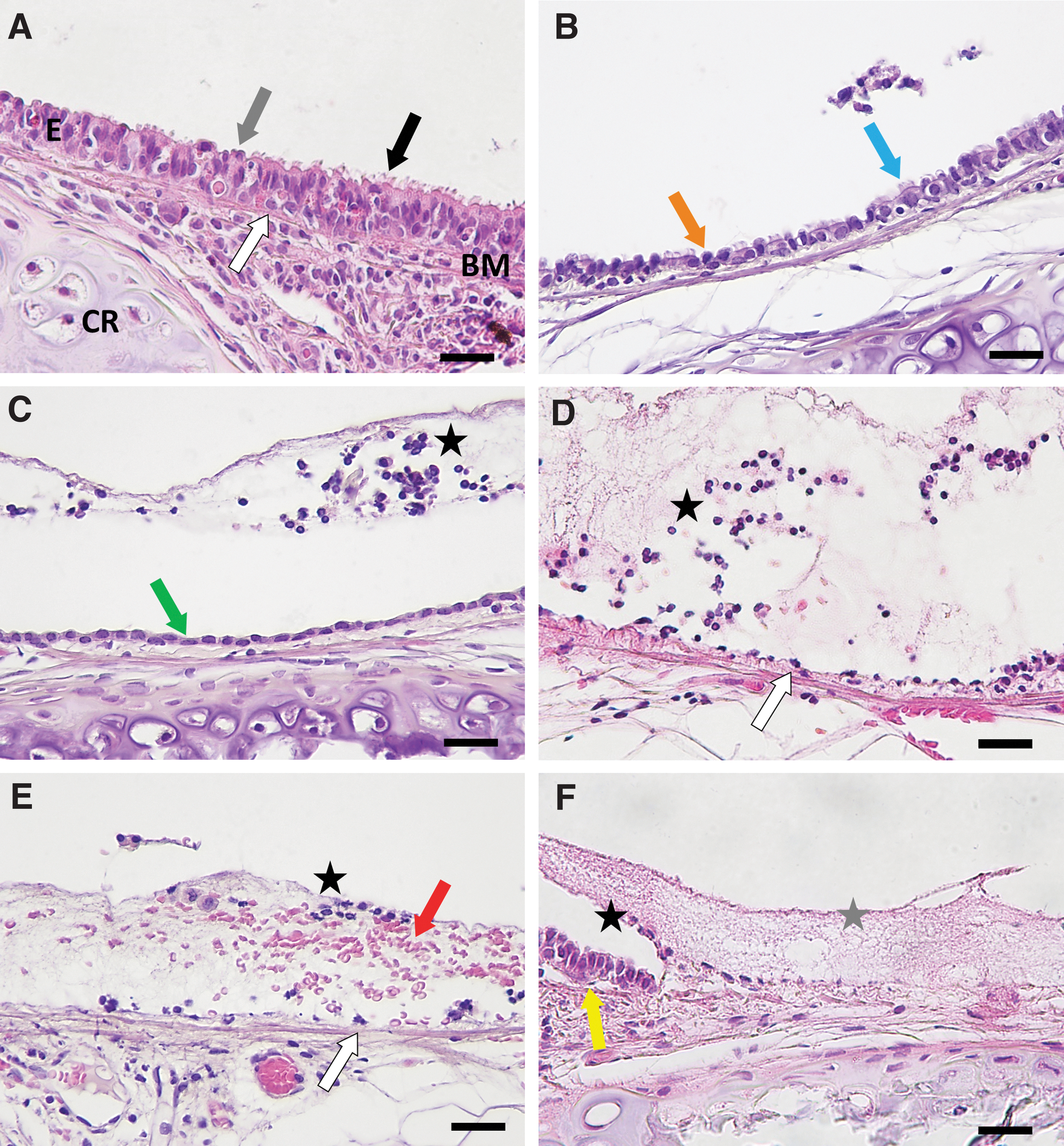
Representative examples of the histological effects of airway conditioning on rat tracheal epithelium. Tracheas were excised 10 min after conditioning with physical perturbation devices and sections stained with H&E. Images from selected animals showed that the unconditioned trachea

The proportion of removed epithelium across the length of the whole trachea after conditioning with physical perturbation devices. Three trachea slides were measured per rat (n = 3 rats per group), and the mean of the three measurements is plotted. The length of epithelial removal (μm) was adjusted with respect to total trachea length (μm). Mean ± SEM presented. An ANOVA and Dunnett's test determined statistical significance with all comparisons performed against the unconditioned group (*p < 0.05, **p < 0.01, ***p < 0.001). ANOVA, analysis of variance; SEM, standard error of the mean.
Common histological findings were identified across conditioning methods (see Table 2 for detailed findings and Supplementary Fig. S1 for a representative image from each device). Conditioned and unconditioned but intubated animals often showed evidence of epithelial disturbance in the proximal trachea due to ET tube insertion. For all perturbation devices, except the Brush and Forceps, the disturbed epithelium was primarily limited to the area over cartilaginous regions, while the epithelium over the intercartilaginous regions consistently showed an intact pseudostratified epithelial cell layer, although some loss of cilia was evident for all conditioning groups.
Summary of the observed trachea histological effects produced by different conditioning methods
The Balloon Catheter and Forceps produced intermittent lengths of flattened and shortened epithelium and dislodged epithelial cells (Fig. 1B–D and Table 2). The Brush dislodged the epithelium along the whole trachea (Fig. 1D and Table 2). Wire baskets consistently resulted in a denuded epithelium and desquamated cells localized over the hyaline cartilage rings (Fig. 1D and Table 2). The Brush and wire baskets left intermittent ovoid cells with a basal cell morphology on the basement membrane in disturbed regions (Fig. 1D). The lumen of brush- and basket-conditioned tracheas contained desquamated cells and basement membrane-like material (Fig. 1D), although this was observed less frequently after the 11582M Stone Extractor procedure. Extravasated erythrocytes were visible in the lumen after conditioning with the Brush (Fig. 1E). Rarely, short sections of completely denuded epithelium, including basal cell dislodgement, were observed for all wire baskets and the Brush (Fig. 1F).
Proportional length of removed epithelium
The nonintubated tracheas did not exhibit significantly different proportions of epithelial removal when compared with the intubated, unconditioned tracheas (Fig. 2). The Brush (0.46 ± 0.11 μm/μm, p = 0.0051), NCircle (0.55 ± 0.06 μm/μm, p = 0.00053), NCompass (0.51 ± 0.06 μm/μm, p = 0.0016), and NGage (0.41 ± 0.05 μm/μm, p = 0.017) devices produced a significantly greater mean proportion of epithelium removal when compared with the unconditioned control group (0.11 ± 0.03 μm/μm).
Based on both the epithelial removal data and histological observations the NCircle, NCompass, NGage, Brush, and 11582M Stone Extractor were investigated for their ability to enhance LV vector-mediated gene transfer. The forceps and balloon catheter were not tested because they produced inconsistent histological effects and were difficult for the operator to employ.
LacZ staining following airway conditioning and LV-LacZ delivery
A statistically significant increase in the mean proportion of en face LacZ-stained area was found for NGage-conditioned tracheas (0.051 ± 0.011 mm2/mm2) when compared with the unconditioned control group (0.0062 ± 0.0026 mm2/mm2, p = 0.00025) (Fig. 3A). The NGage device produced an eightfold increase in the mean proportion of LacZ-stained area compared with unconditioned tracheas. Notably, there is one outlier in the NGage group that produced a substantially higher proportion of staining when compared with the other animals within the same group. No statistically significant differences were found for other conditioning groups (Fig. 3A).
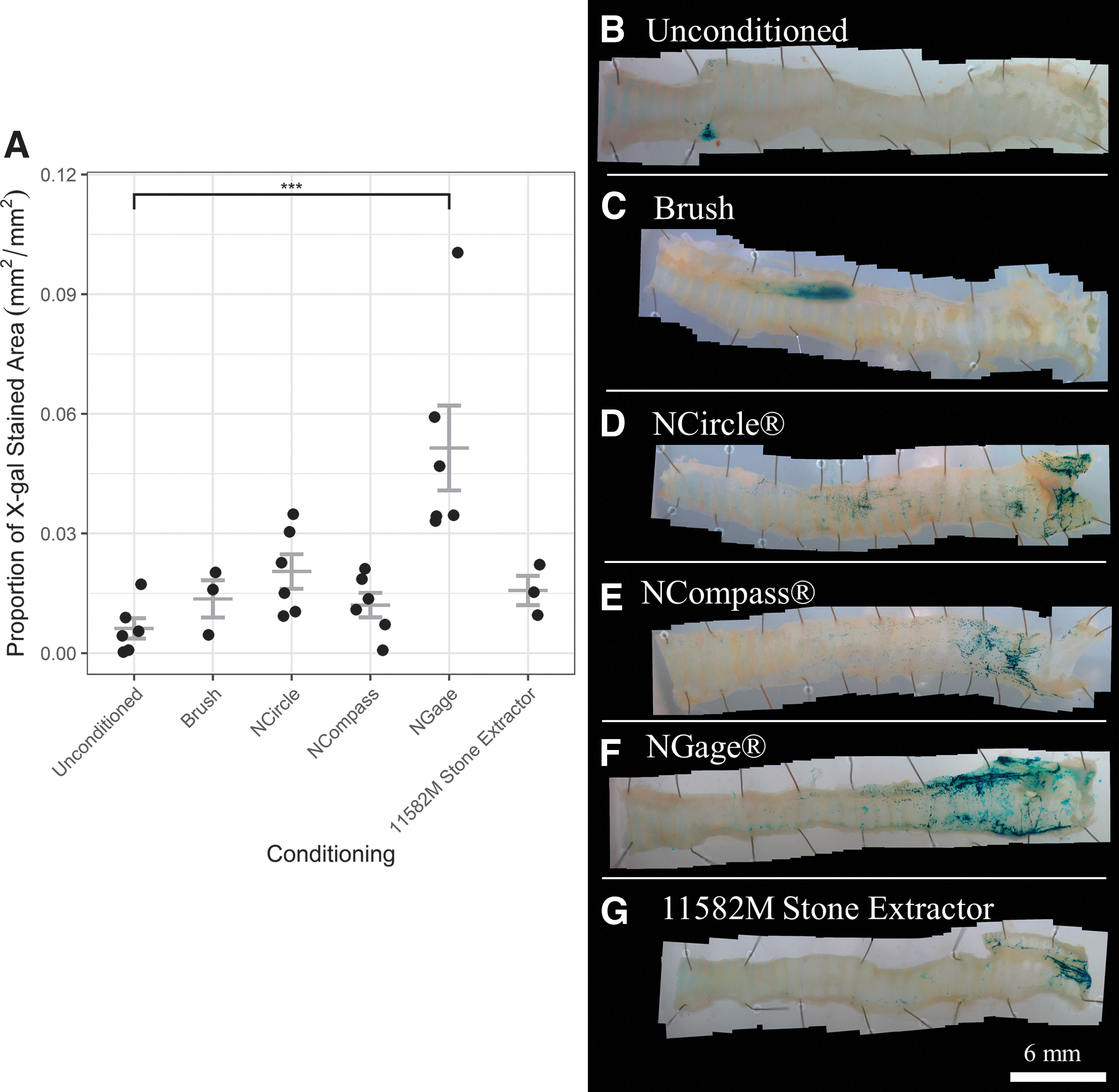
In unconditioned tracheas, en face LacZ staining was primarily observed in a cluster at the proximal end of the trachea around the location of the ET tube (Fig. 3B). Conditioned tracheas also exhibited some LacZ staining in the proximal region, but the majority of the staining was in the distal trachea and bronchi, regardless of device (Fig. 3C–G). LacZ staining was also observed in locations where the device was not applied, including the alveolar tissue. However, staining in those regions was not quantified as our aim was to assess the transduction in response to the physical perturbation that was only applied to the trachea. Conditioning with wire baskets resulted in LacZ staining in horizontal, striated lines over the cartilage rings, although it was not limited to these areas (Fig. 3D–G). Histological analysis confirmed the presence of LacZ-positive cells in the epithelium over the cartilage rings (Fig. 4).
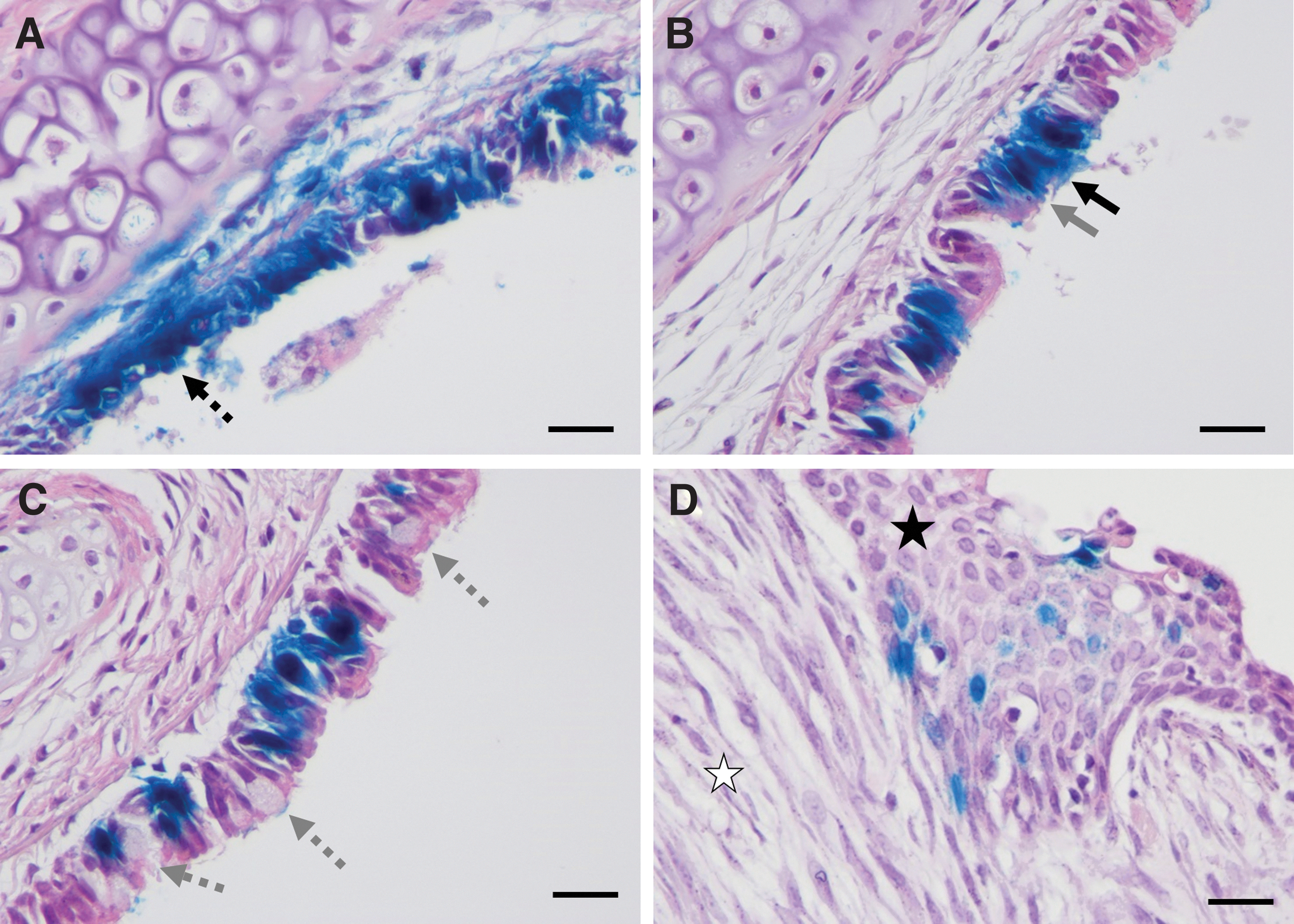
Histological observations of LacZ-positive cells and epithelium repair at 7 days post-gene transfer in H&E-stained tracheas. LacZ-positive cells are stained blue. LacZ-positive cells were
One trachea conditioned with the Brush had only clusters of LacZ staining around the carina, however, the other two tracheas exhibited abnormal tissue growth protruding into the lumen and out of the posterior side of the trachea near the trachealis muscle. Histological assessment of the tissue masses revealed hyperplasia of the tracheal epithelium and fibroplasia in the lamina propria, along with scattered LacZ-expressing cells (Fig. 4D). Two NGage-conditioned tracheas exhibited similar abnormal growths, although they were notably smaller in size (not shown).
At 7 days post-gene transfer, the epithelium contained regions of tightly packed pseudostratified epithelial cells, while other sections consisted of cuboidal cells (Fig. 4A). LacZ-positive ciliated and nonciliated epithelial cells were identified (Fig. 4B). Mild goblet cell hyperplasia was visible in the tracheal epithelium of an NCircle-conditioned trachea (Fig. 4C). In rare instances, connective tissue below the basement membrane and cells in the lamina propria were LacZ-stained (Fig. 4A). Basal cells that were positive for LacZ and cytokeratin 5, a basal cell marker, were identified in tracheal sections from all conditioning groups (Fig. 5 and Supplementary Fig. S2). For the unconditioned control group, LacZ-positive basal cells were only identified in the proximal trachea, where epithelium damage occurred during intubation (Supplementary Fig. S2).
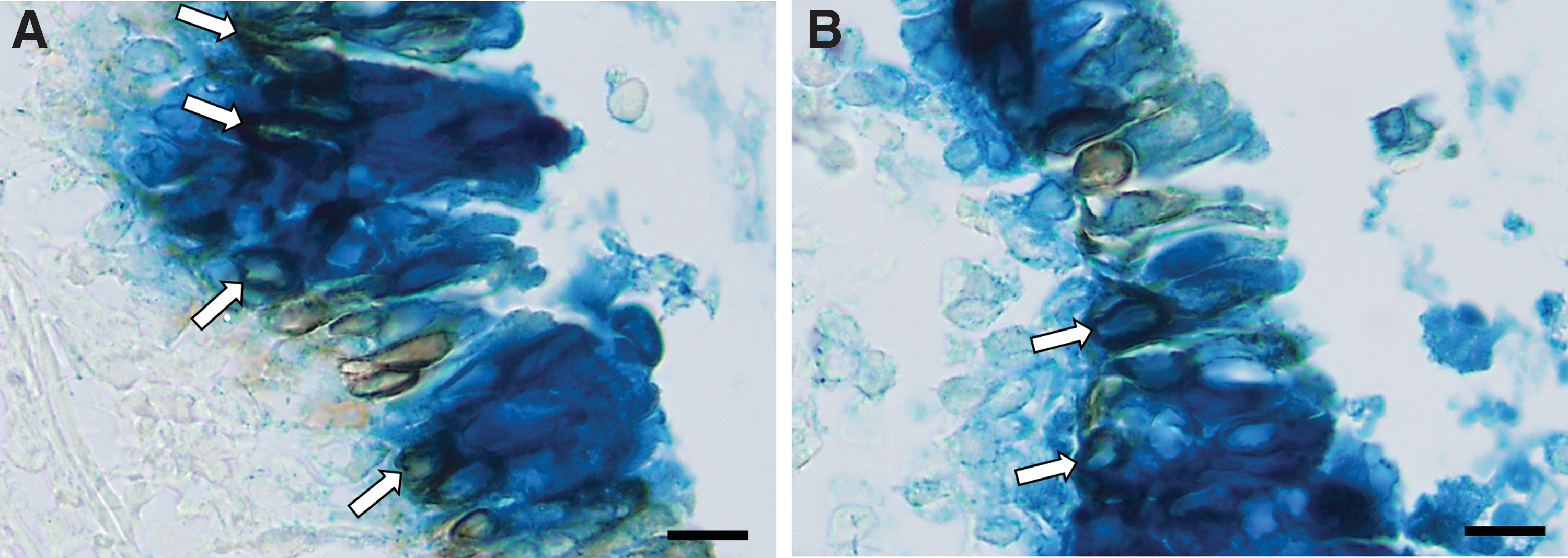
Examples of LacZ-positive basal cells at 7 days posttransduction (white arrows) using
DISCUSSION
A growing body of knowledge indicates that physically disrupting the integrity of the airway epithelium facilitates vector access to the basolateral surface and improves gene transfer. 16 –19 However, the unique effects produced by different perturbation devices on airway epithelium in vivo, and how these effects may aid gene transduction has not yet been examined. This study was the first to assess and quantify the histological and LV gene transfer effects of physical conditioning devices on the tracheal epithelium in rats. Our results demonstrated that a range of perturbation devices could be employed to disturb the airway epithelium, and this study found the NGage was the most effective device to enhance gene transfer when compared with an unconditioned control.
All conditioning devices resulted in epithelial cell removal, but the location, severity, and affected cells varied between groups, likely influencing the relative effectiveness of devices for conditioning. Epithelium disruption was more severe over the cartilaginous regions, likely because cartilage rings protrude further into the lumen than the intercartilaginous regions, 24 leading to greater contact with the devices. Devices dislodged epithelial cells to varying degrees, with the Brush and wire baskets leaving intermittent cells on the basement membrane, consistent with previous reports. 14,25 The Forceps and the Balloon Catheter produced nonhomogenous and often minimal disruption, potentially because the device had limited points of contact with the airway epithelium or exerted less pressure on the epithelium over a wider area, respectively; these devices were not pursued further here.
The Brush, NCircle, NCompass, and NGage devices removed a greater proportion of the epithelium compared with unconditioned tracheas because these devices likely had the most points of contact with the epithelium. Epithelial removal was not significantly different between the nonintubated and unconditioned (but intubated) tracheas, however, epithelial disturbance identified in the proximal trachea of unconditioned rats was likely due to ET insertion. 26
Physical perturbation using a variety of devices, including a brush and the NCircle basket, has previously improved gene transfer to the tracheal epithelium. 17 –19 Based on visual assessment of LacZ staining, all devices trialed in this study appeared to have enhanced gene transfer to varying degrees and therefore may be candidates for further investigation. Evaluation of en face images also revealed distinct differences in the location and quantity of LacZ staining between conditioned and unconditioned tracheas. Upon statistical comparison of the proportion of LacZ-stained area, the novel NGage wire basket was the only device that resulted in a significant increase over the unconditioned group.
Physical perturbation may increase gene transfer through a number of potential mechanisms, including: (1) removing the protective mucus layer or disrupting mucociliary transport to increase vector access to cells, (2) exposing basolateral VSV-G receptors on epithelial and basal cells, 27 or (3) removing poorly transducible epithelial cells to facilitate access to more readily transduced cells such as basal cells or remaining epithelial cells in disturbed regions. 16,17 In support of these mechanisms, greater gene transfer appeared to occur in disturbed regions. LacZ-expressing cells at 7 days post-transduction were predominantly observed in the epithelium over the cartilage rings, where dislodged cells and exposed basal cells had been observed histologically 10 min postperturbation. However, the possibility that LacZ-positive cells migrated to the damaged regions during epithelial repair cannot be excluded. 14,28
To provide further support for the proposed mechanisms, cytokeratin-5 antibody staining confirmed that all devices tested resulted in LacZ-positive basal cells, often in regions of disturbance identified in the initial histological assessment. Importantly, the ability to transduce basal cells suggests the potential for physical perturbation to facilitate long-term transgene expression for lasting gene therapy. 12
In a previous study, the NCircle significantly increased the area of LacZ reporter gene staining in rat tracheas by 1,000-fold over unconditioned controls 7 days after LV vector transduction. 19 Contradicting these findings, the present study found the LacZ-stained area after NCircle conditioning was not significantly different from the unconditioned control. This may be due to a difference in operator and/or technique; the technique used in the present study twisted the wire basket in the distal trachea, possibly causing it to collapse and reduce the area or severity of perturbation. Furthermore, the previous study used a LacZ reporter gene under the control of a MPSV viral promoter, 19 rather than EF1ɑ, which is thought to be a weaker promoter and may have generated less transgene expression, possibly below detection thresholds. 29 –32
For the first time, this study explored the relationship between the degree or type of epithelial disturbance and the amount of gene transfer produced by different devices and revealed a nuanced relationship. The en face images indicate that epithelial cell disturbance is beneficial for conditioning, as all physical perturbation devices conferred some gene transfer enhancement when compared with the unconditioned control group. However, while most devices significantly increased epithelial removal, only the NGage device produced significant enhancement in the area of LacZ staining. This suggests that epithelial cell removal may not be the most informative predictor of the efficacy of a conditioning method. Other factors may be of greater importance when predicting the ability of a device to enhance gene transfer, such as the location and type of disturbance, and the identity and viability of the remaining cells following perturbation.
Although not tested in this study, it is possible that a greater number of viable basal cells may have remained on the basement membrane following perturbation with the NGage wire basket, leading to an increase in LacZ-stained area. Cell viability assays, 33 markers to identify cell types, 28 or a grading scale to examine less severe histological effects, 26 including epithelial shortening and deciliation, could be used in future studies to provide a more comprehensive understanding of the histological effects and their relationship to gene transfer.
Repair of the pseudostratified epithelium was ongoing at 7 days post-gene transfer as sections of poorly differentiated cuboidal epithelium persisted, consistent with previous observations that epithelium repair following perturbation can take more than 1 week in rats. 19,28 A common response to airway injury is transient epithelial hyperplasia, which acts to rapidly regenerate the epithelium. 25,34 Brush- and NGage-conditioned tracheas exhibited epithelial hyperplasia and fibroplasia of the lamina propria at 7 days post-gene transfer. This persistent and extensive hyperplasia has not been reported after airway abrasion. The Brush and the NGage basket was less pliable than other devices and possibly exerted a stronger mechanical force on the epithelium and basement membrane, causing severe damage and stimulating this prolonged, abnormal hyperplasia.
Our finding of transduced cells within the lamina propria demonstrated that severe perturbation could damage basement membrane integrity, as viral vectors cannot penetrate an intact basement membrane. 16 Brushing airways has previously extravasated erythrocytes into the lumen, 34 an effect again observed here, which is suggestive of undesirable damage to the airway mucosal layer. Targeted epithelial disturbance will be necessary to ensure the safe and effective use of physical perturbation approaches as a conditioning method.
This novel investigation of epithelium disturbance and gene transfer aids the identification of device features and histological effects that facilitate the enhancement of gene transfer. A flexible device that conforms to airways (avoiding the severe damage produced by the rigid Brush and NGage) but still with many points of contact (like the collapsible wire baskets) may be optimal to disturb extensive sections of epithelial cells to predefined degrees. Controlled perturbation that leaves a layer of viable basal cells may maximize gene transfer to this cell type, which can clonally expand after airway injury to produce widespread LacZ-expressing cells. 12,35
Some experimental groups in this study had small sample sizes, however, these results still provide informative proof-of-principle data. Currently, in vivo physical perturbation is an exploratory technique, but the promising results of this study justify further experiments to optimize the perturbation technique and devices in the lower conducting airways. In rats, direct airway visualization through a bronchoscope will likely be essential for future controlled and targeted application. 36 In addition, application of these methods to larger animal models and potentially to humans appears straightforward using standard bronchoscopic equipment and techniques. Furthermore, while physical perturbation appeared to be safe and effective in normal rats, future studies should investigate these methods in a CF model, including assessment in the presence of bacterial infection.
Several refinements to perturbation should now be explored. One avenue involves testing the effect of different intervals between conditioning and vector delivery on gene transfer effectiveness, as our preliminary unpublished data suggest that delivery immediately following epithelium disturbance is also effective to enhance gene transfer. Future quantification of LacZ-positive basal cell numbers produced by different devices would also provide valuable information on the comparative ability of these devices to facilitate stem cell transduction, an important target for sustained gene therapy. Although this study has revealed beneficial associations between epithelial disturbance and gene transfer, future studies should explore the mechanisms of action and verify causative relationships. Quantification and assessment of histological parameters other than epithelial removal may prove valuable to further understand which effects confer gene transfer enhancement.
CONCLUSION
The novel NGage device was the most effective of the conditioning methods tested, significantly increasing the proportion of LacZ-stained area eightfold when compared with unconditioned tracheas. Interestingly, other devices also produced significant epithelial cell removal, but these did not significantly improve gene transfer. This indicates a complex relationship between epithelium disturbance and gene transfer and suggests that factors other than epithelial removal play a role in conferring gene transfer enhancement. The promising results of this study support the continued investigation of physical perturbation as an airway conditioning method for LV vectors and potentially other gene delivery vehicles. Enhanced gene transfer facilitated by perturbation has the potential to improve the efficacy of LV vector-mediated gene therapy to reach the requisite levels for treatment of lung diseases.
Footnotes
ACKNOWLEDGMENTS
The authors thank Prof. John Finnie, Senior Comparative Pathologist, University of Adelaide, for assistance with interpretation of histological findings; Dr. Emma Knight for advice on statistics; Dr. Jason Gummow for producing and titering the vector at the Gene Silencing and Expression Facility; and Dr. Juliette Delhove for designing and generating the LV construct. Assistance was also provided by Laboratory Animal Services for animal husbandry and monitoring, and Histology Services at the University of Adelaide.
AUTHORs' CONTRIBUTIONS
V.D. contributed to study design, data collection, data analysis and interpretation, and article drafting. P.C. contributed to study design, data collection, provision of knowledge, and article drafting. M.D. and D.P. contributed to study conception, data analysis and interpretation, provision of knowledge, and article drafting and editing. N.R. contributed to study design, data collection, provision of knowledge, and article drafting. A.M. contributed to study conception and design, data collection, data analysis and interpretation, provision of knowledge, and article drafting. All authors gave final approval of the article.
AUTHOR DISCLOSURE
A.M., D.P., and M.D. are inventors on a Patent Application No. PCT/AU2021/051164 entitled “Methods and Devices for Delivering Agents to the Respiratory System.” The other authors have no competing financial interests to declare.
FUNDING INFORMATION
Funding for this project was provided by the National Health and Medical Research Council (GNT1160011) and the Cystic Fibrosis Foundation (DONNEL21GO).
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.