
Abstract
Lentiviral transduction of human mesenchymal stem cells (MSCs) induces long-term transgene expression and holds great promise for multiple gene therapy applications. Polybrene is the most commonly used reagent to improve viral gene transfer efficiency in laboratory research; however, it is not approved for human use and has also been shown to impair MSC proliferation and differentiation. Therefore, there is a need for optimized transduction protocols that can also be adapted to clinical settings. LentiBOOST (LB) and protamine sulfate are alternative transduction enhancers (TEs) that can be manufactured to current Good Manufacturing Practice standards, are easily applied to existing protocols, and have been previously studied for the transduction of human CD34+ hematopoietic stem cells. In this study, we investigated these reagents for the enhancement of lentiviral transduction of adipose-derived MSCs. We found that the combination of LB and protamine sulfate could yield comparable or even superior transduction efficiency to polybrene, with no dose-dependent adverse effects on cell viability or stem cell characteristics. This combination of TEs represents a valuable clinically compatible alternative to polybrene with the potential to significantly improve the efficiency of lentiviral transduction of MSCs for gene therapy applications.
INTRODUCTION
Mesenchymal stem cells (MSCs) are used widely in the fields of tissue engineering and regenerative medicine for their multipotency, immunomodulatory properties, and ease of isolation. 1 –7 Ex vivo genetic modification of MSCs using viral transduction can expand their scope of clinical application by enhancing their therapeutic potential through the transfer and subsequent expression of specific target genes. For example, ex vivo transduction of MSCs using a lentiviral vector with the cDNA for the osteoinductive transgene BMP-2 has been associated with robust bone regeneration in preclinical, critical-sized bone defect models. 8 –10
Long-term transgene expression in transduced MSCs is desirable in many clinical contexts, including bone repair, where bone healing takes place over the course of several months. While viral and nonviral methods for gene transfer exist, lentiviral vector transduction is a well-established method specifically for prolonged transgene expression. 11 Thus, lentiviral transduction is an important gene therapy tool for clinical applications such as bone loss.
Standard transduction protocols use concentrated lentiviral vector preparations in the presence of various transduction-enhancing reagents to improve gene transfer rates. 12 Transduction enhancers (TEs) are small molecule polymers or peptides that typically serve to increase viral entry by altering cell surface charges or cell membrane permeability. 12 –16 Without TEs, lentiviral transduction of MSCs is low.
Polybrene, a polycationic polymer, is a widely used TE in research. 14,15 It is inexpensive, easy to use, and highly effective for a variety of cell types, including MSCs. 13,15,17 However, in MSCs, polybrene impairs proliferation and differentiation, which raises concerns about its use for translational research in scenarios where implanted MSCs may serve as progenitors for tissue regeneration. 18 For these reasons, there is a need to improve protocols for lentiviral transduction of MSCs that avoid polybrene and can be adapted for clinical applications.
Schott et al described the characteristics of an ideal TE as having (1) a positive influence on transduction, (2) a lack of cytotoxicity, (3) preservation of stem cell characteristics, (4) an ability to be manufactured to Good Manufacturing Practice (GMP) standards, and (5) easy application. 19 Previous research has identified an expanding list of lentiviral TEs meeting these criteria, but the vast majority of this work has been done in CD34+ hematopoietic stem and progenitor cells (HSPCs). 12,13,19 –23
The TEs, LentiBOOST (LB) and protamine sulfate (PS), have been previously reported to enhance transduction. Both LB and PS can be manufactured to GMP standards and are easily incorporated into the transduction milieu without the need for additional cell manipulation. 13,24 Additionally, there are additive effects when the combination of LB and PS is used for HSPC transduction. 19,20,22,23 However, LB has not been previously evaluated in MSCs and no studies have evaluated combinations of TEs in MSCs.
There are inherent differences between MSCs and HSPCs as well as differences in standard MSC and HSPC transduction protocols that may influence the effectiveness of TEs at a given concentration. 25 –30 Therefore, the purpose of this study was to evaluate the effects of LB and PS, both individually and in combination, on the enhancement of lentiviral transduction of adipose-derived MSCs (ASCs).
We hypothesized that the combination of LB and PS would have an additive effect on transduction efficiency and reporter gene expression and that higher concentrations would be associated with increased transduction efficiency and transgene expression, with minimal toxicity.
MATERIALS and METHODS
Tissue isolation and cell culture
After Institutional Review Board approval, adipose tissue was collected from the infrapatellar fat pads of three healthy patients (two males and one female, mean age of 70 ± 8 years) undergoing primary total knee arthroplasty at our institution. This tissue is normally discarded after surgery. The samples were processed immediately after collection, and standard previously published protocols were used for isolation of the stromal vascular fraction and culture expansion. 2,31
After culture expansion, passage 3 cells were plated at a density of 0.2 × 106 cells/well in six-well plates, with a total of 18 × 106 cells plated for each donor. The following day, the medium was exchanged with 1 mL of fresh growth medium for viral transduction.
TEs and experimental groups
LB (SIRION Biotech, Planegg, Germany) was stored at −20°C, thawed before use, and then added directly to wells containing cells and cell media. We evaluated LB at concentrations of 0.1, 0.33, 1.0, 3.3, and 10 mg/mL. Concentrations were chosen based on previous reports and manufacturer recommendations. 13,20,32
PS (Fresenius Kabi, Lake Zurich, IL) was prepared in a stock solution of 10 mg/mL by diluting with cell culture grade water and stored as single-use aliquots at room temperature. Based on unpublished data from our laboratory evaluating the effects of PS on transduction of human ASCs, a concentration of 100 μg/mL PS was chosen for this study. This is consistent with PS concentrations reported in other studies for lentiviral MSC transduction. 24
Polybrene (Sigma-Aldrich, St. Louis, MO) was prepared in a stock solution of 8 mg/mL (1,000 × concentrated) with double distilled H2O and stored as single-use aliquots at −20°C. Thawed polybrene stock was added directly to media at a concentration of 8 μg/mL and filtered using a 0.45-μm filter before transduction.
A total of 15 experimental groups were evaluated for each donor and are described in Table 1. Two samples from each donor were analyzed separately for each experimental group.
Overview of experimental groups
LB dosing in mg/mL. Polybrene and PS dosing in μg/mL.
LB, LentiBOOST; PB, polybrene; PS, protamine sulfate; TE, transduction enhancer.
Lentiviral transduction of human ASCs
Cells were transduced overnight by adding vesicular stomatitis virus G-protein pseudotyped lentiviral vectors expressing enhanced green fluorescent protein (eGFP) under the control of the murine leukemia virus long terminal repeat (RhMLV) promoter (LV-GFP). 11 LV-GFP was added directly to the cells at a multiplicity of infection (MOI) of 3. Transduction was conducted either in the absence or presence of LB, PS, and polybrene at concentrations stated earlier (Table 1). The following day, the cells were washed twice with PBS and 3 mL of fresh growth media was added. Media were changed every 3–4 days for the duration of the experiment.
Analysis of the transduced cells included an evaluation of transduction efficiency, eGFP expression level, cytotoxicity, and differentiation potential. Analysis was performed for each of the 15 study groups for each donor. Our experimental design is depicted in Fig. 1 and detailed below.

Experimental scheme. Human adipose-derived MSCs were expanded in culture to passage 3 and then plated at a density of 0.2 × 106 cells/well in six-well plates, using two replicates per donor. MSCs were transduced with LV-GFP (MOI = 3) overnight in the presence or absence of TE(s). Cells were washed the following day and fresh media were added. Analysis included FACS for GFP-expressing cells, ddPCR to determine the vector copy number per cell, trypan blue live cell staining, and CFU-fibroblast assay (created with
Flow cytometry
Transduction efficiency and median fluorescence intensity (MFI) were evaluated at 2 and 7 days post-transduction by detection of GFP expression. Briefly, cells were harvested, washed, and centrifuged for 5 min at 200 g. Pellets were resuspended with flow cytometry (FACS) buffer (PBS with 2% FBS) containing DAPI at a working concentration of 1 μg/mL. Cells were recorded on an Attune NxT flow cytometer (Thermo Fisher) and analyzed using FlowJo software (Tree Star, Inc., Ashland, OR).
Vector copy number determination
Vector copy number determination was performed in accordance with the method described by Masiuk et al, with minor modifications, which is described in detail in the Supplementary Data. 23
Cell viability
After overnight exposure with or without TEs, cell viability and proliferation were assessed at 7 and 14 days post-transduction. Separate wells were analyzed at each time point. Cells were harvested, washed, and centrifuged for 5 min at 200 g. Cells were counted before FACS at day 7 and before vector copy number (VCN) determination at day 14. Live cell count was determined using trypan blue and an automated cell counter.
Colony-forming unit assay
Colony-forming unit-fibroblast (CFU-F) assays were performed using standard techniques. Cells were aliquoted before FACS on day 2 post-transduction. Briefly, 100 transduced ASCs were plated in 10-cm culture plates in duplicate. Cells were maintained in culture with standard growth medium for 14 days with medium changes every 3–4 days. After 14 days in culture, the cells were washed and stained with crystal violet, and colonies were manually counted by two different readers (M.G. and S.C.).
Statistical analyses
Data were assessed for normality using histograms and Shapiro–Wilk tests and for equality of variances using Levene's test and by evaluating residual plots. Kruskal–Wallis tests with post hoc pairwise Wilcoxon rank-sum tests with Bonferroni correction were performed to evaluate the association between TE concentrations and outcomes of interest (transduction efficiency, MFI, VCN, cell counts, and CFUs); polybrene was set as the reference level for all comparisons. Next, linear mixed effects modeling was performed.
Two random intercepts were included in each model to account for variation within subjects and variation within samples nested within subjects. The concentration group was included as a fixed effect in all models. Ninety-five percent confidence intervals for all fixed effect estimates were calculated using restricted maximum likelihood. All tests were two-sided and a p-value ≤0.05 was considered statistically significant.
All analyses were performed using R software, version 4.1.1 (R Foundation for Statistical Computing, Vienna, Austria).
RESULTS
Single TEs for MSC transduction
In the TE(−) control group, FACS analysis at days 2 and 7 revealed low percentages of GFP-expressing cells at both time points (<23%) and an average VCN per cell of 1.03 (Fig. 2 and Supplementary Table S1). In the polybrene control group [PB(+)], transduction efficiency and VCN per cell were 49% and 7.9, respectively. In linear mixed effects modeling, the PB(+) group had significantly higher transduction efficiency at 2 and 7 days (p < 0.001 and p < 0.001, respectively), MFI (p < 0.001 and p < 0.001, respectively), and VCN per cell (p < 0.001) compared with the TE(−) group (Supplementary Table S1). The use of PS alone yielded transduction efficiency of 33% and 32% at 2 and 7 days, respectively, and a VCN per cell of 2.1.

Effects of TEs and TE combinations on lentiviral transduction of MSCs. Dose–response relationships were observed for transduction efficiency with the use of LB alone and in combination with PS
LB alone demonstrated improved transduction at all time points and at all concentrations tested compared with the TE(−) control group (Fig. 2). LB increased the percentage of GFP+ cells in a dose-dependent manner, with values ranging from 25% at 0.1 mg/mL to 42% at 10 mg/mL. Similarly, there was a positive association between LB concentration and VCN per cell.
At all concentrations of LB alone, values of transduction efficiency, MFI, and VCN per cell were lower than the corresponding values for the PB(+) group (Fig. 2). However, many of these differences were not statistically significant (Supplementary Table S2). There were no significant differences in transduction efficiency between the PB(+) group (49%) and LB at concentrations of 1, 3, and 10 mg/mL (34%, p = 0.11; 38%, p = 0.37; and 39%, p = 0.41, respectively).
For MFI, the PB(+) group had a significantly higher expression level compared with all concentrations of LB at 2 days, but these differences were no longer significant at 7 days. For VCN per cell, the PB(+) group had significantly higher mean values (∼8) compared with all concentrations of LB alone (range 1.5–3.2).
Combination of LB and protamine sulfate for MSC transduction
As shown in Fig. 2, addition of PS to LB resulted in substantial improvements in transduction efficiency, MFI, and VCN per cell, compared with LB alone. The combination with the lowest concentration of LB tested (0.1 mg/mL LB and PS) had a 2-day transduction efficiency of 37% compared with 33% for the PS-alone group and 26% for the 0.1 mg/mL LB group (Fig. 2 and Supplementary Table S1). The effects were even greater at higher concentrations of LB: the combination with the highest concentration of LB tested (10 mg/mL LB and PS) exhibited a 2-day transduction efficiency of 66% compared with 39% for 10 mg/mL LB alone (p = 0.001), and MFI and VCN per cell were also increased compared with LB alone.
In the linear mixed effects models, polybrene was chosen as the reference to evaluate whether combinations of LB and PS could serve as potential alternatives (Supplementary Table S2). TE combinations with LB concentration ≥1.0 mg/mL had higher mean values for transduction efficiency, MFI, and VCN per cell when compared with the PB(+) group (Fig. 2). However, our modeling showed that only the combination 10 mg/mL LB+PS group had significantly improved transduction efficiency (2 days, p = 0.033; and 7 days, p = 0.029) and MFI (2 days, p < 0.001; and 7 days, p < 0.001) compared with PB(+) (Supplementary Table S2).
The only other statistically significant difference between any combination TE group and the reference standard PB(+) was a significantly lower VCN per cell in the 0.1 mg/mL LB+PS group (2.63 vs. 7.87, p = 0.011). Combinations with 0.33, 1.0, and 3.3 mg/mL LB were not found to be statistically significantly different compared with PB(+) for transduction efficiency, MFI, or VCN per cell.
Proliferation and differentiation of TE-treated MSCs
At both 7 and 14 days post-transduction, we did not observe any significant differences in the number of live cells between the 15 study groups (p = 0.52 and p = 0.99, respectively), demonstrating an apparent lack of cytotoxic effects (Fig. 3 and Supplementary Table S3). The PB(+) group tended to have lower cell counts at both time points compared with all other treatment groups, but these differences were not significant (Supplementary Table S2).
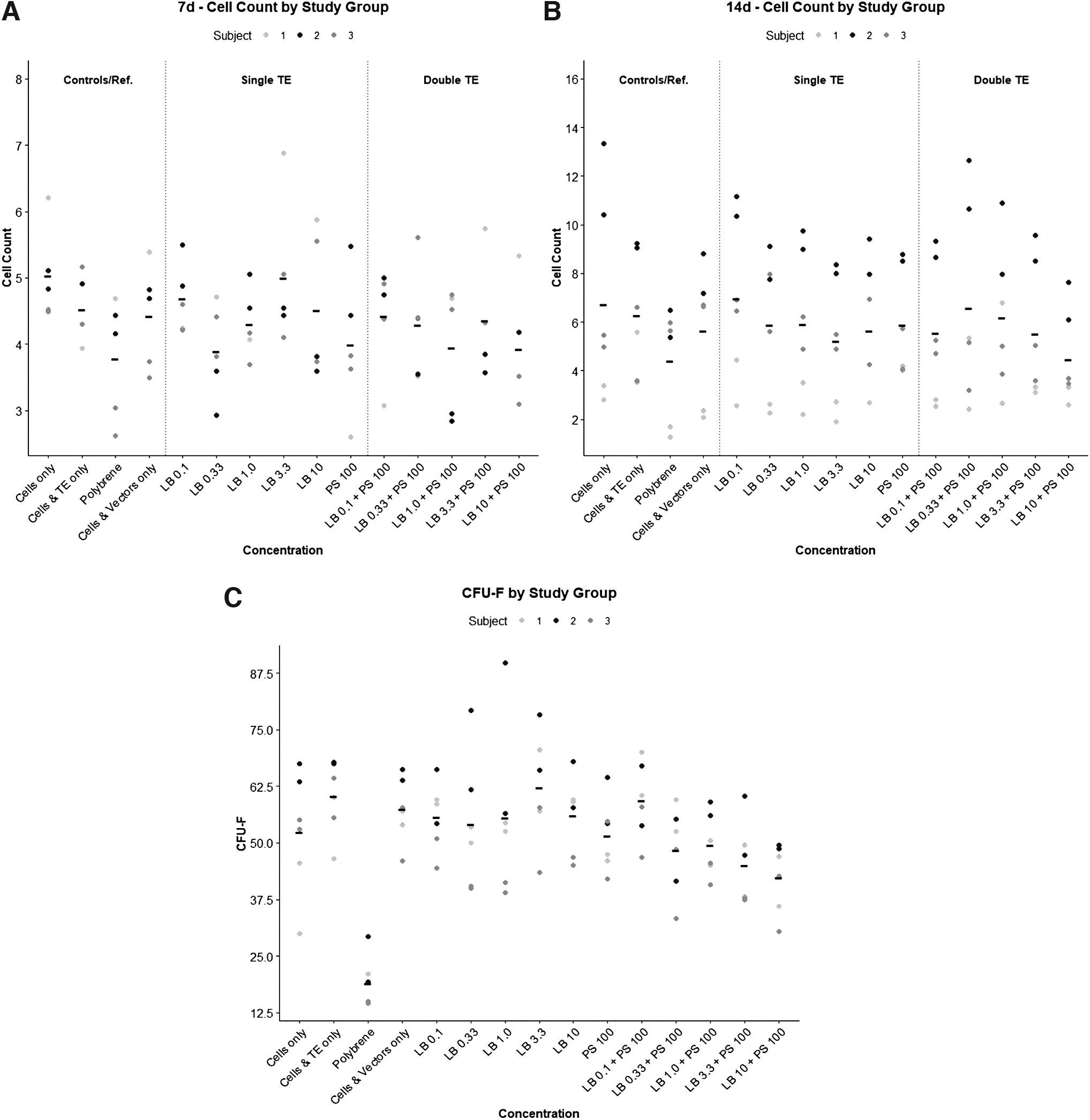
LB and protamine sulfate do not affect viability or colony-forming potential of MSCs. No significant differences were noted between LB concentration (either alone or in combination with PS) and cell proliferation, as determined by cell counts at 7 and 14 days post-transduction
As shown in Fig. 3, colony formation in cells treated with LB, PS, or both was similar to nontransduced TE(−) controls. Colony formation in the PB(+) group was significantly lower compared with all other treatment groups (p < 0.001).
DISCUSSION
Our study found that the combination of LB and protamine sulfate markedly enhances lentiviral MSC transduction and can achieve comparable or superior results to polybrene with negligible toxicity. We observed that the use of LB alone had a positive influence on transduction and resulted in average transduction efficiency of 42% at an MOI of 3 at the highest concentration tested (Fig. 2). However, all concentrations of LB alone were associated with lower transduction efficiency, VCN per cell, and MFI compared with the PB(+) group (Supplementary Table S2), although these differences did not reach statistical significance.
Our findings on LB-mediated transduction of MSCs stand in contrast to prior studies in HSPCs, which have shown transduction efficiency of >80%. 19,20,23 Additionally, Hauber et al investigated the effects of LB on LV-mediated transduction of HSPCs and did not observe the same dose-dependent relationship between LB concentration and transduction efficiency and MFI that we observed in MSCs. 20 Their findings may be explained by vector saturation, as they used an MOI of 10 (≥10 is typical for HSPCs), and transduction efficiency was >80% even at the lowest LB concentration tested (0.5 mg/mL). 20
In the same study, the authors found that transduction efficiency was about 50% in the absence of TEs at an MOI of 3, which is more than twice that seen in the present study. These differences underscore the need to interpret studies on TEs with caution due to the multitude of factors that influence transduction, including cell type and MOI.
With respect to PS alone, we found that PS at a dose of 100 μg/mL improved transduction efficiency, but tended to be inferior to the PB(+) group (7-day efficiency: 32.4% vs. 48.4%, p = 0.047; and VCN per cell: 2.1 vs. 7.9, p = 0.005) (Fig. 2 and Supplementary Table S2). These results suggest that a high dose of PS may not be a comparable alternative to polybrene.
In contrast, Lin et al recently concluded that a high dose of PS (100 μg/mL) was a viable alternative to polybrene for MSC transduction. 24 However, this group used a higher MOI and bone marrow-derived MSCs, whereas our study used adipose-derived MSCs. It is possible that the MSC tissue source has an effect on the utility of various TEs and TE combinations for enhancement of cell transduction.
We observed additive effects of LB and PS when used in combination. A possible explanation is that LB and PS work by different mechanisms. LB contains Poloxamer 388/Pluronic F108, which belongs to a class of amphiphilic nonionic copolymers, which interact with biological membranes and are thought to increase viral fusion and transmembrane transport. 13,33 –35 PS, such as polybrene, is a polycationic polymer and is thought to enhance viral entry by reducing charge repulsion between the vector and the cell membrane. 14 –16
When used in combination, these mechanisms may complement each other to enhance transduction. Many of the combination groups achieve comparable or higher transduction efficiency and MFI when compared with polybrene (Fig. 2 and Supplementary Table S2). Our data suggest that PS with LB doses between 0.33 and 1.0 mg/mL may provide an effective alternative to polybrene. PS combined with higher concentrations of LB exhibited further increases of all transduction parameters, but VCN per cell was higher than the FDA-recommended VCN value of <5 for LV-based products. 36
In our study, average VCN exceeded this level only at LB+PS concentrations higher than 0.33 mg/mL LB. At this concentration, transduction efficiency and MFI were comparable with the reference standard polybrene (Supplementary Tables S1 and S2), and at no concentration of LB+PS was VCN statistically significantly higher than the reference standard polybrene, suggesting that LB+PS may present a viable, FDA-compliant, clinically useful TE preparation.
Live cell staining demonstrated that PS and LB had no adverse effects on cell viability and proliferation at 7 and 14 days post-transduction. The CFU-F assay demonstrated significantly decreased colony-forming potential in polybrene-treated cells when compared with PS or LB. These findings are in agreement with prior work from Lin et al, which demonstrate polybrene-associated cell toxicity and impaired differentiation. 24
There are limitations associated with this study. First, one of the three subjects was an outlier with respect to transduction efficiency and VCN per cell (Fig. 2). Transduction efficiency for this subject did not exceed 25% for any of the TEs or TE combinations tested. Subsequently, transduction efficiency was significantly lower, on average, than previously reported. In our analysis, we sought to limit this source of variance by performing linear mixed effects modeling to account for intersubject variation.
Second, we assessed adipose-derived stem cells in this study and the results should not be generalized to other cell types.
In conclusion, the combination of LB and PS as TEs for lentiviral transduction of MSCs presents a promising alternative to polybrene. We observed synergistic effects when combining LB and PS, with LB dose-dependent increases in transduction efficiency, MFI, and VCN, with minimal cytotoxicity.
Collectively, our results provide evidence for a clinically compatible combination of TEs for lentiviral transduction of adipose-derived stem cells with the potential to improve the efficient use of viral vector and cells for MSC-based gene therapy research.
Footnotes
ACKNOWLEDGMENTS
The authors would like to acknowledge and thank Jeffrey Boyd and Bernadette Masinsin of the USC Flow Cytometry Core for their guidance and expertise, as well as critical resource provision for this project.
AUTHORs' CONTRIBUTIONS
K.C. was involved in conceptualization, methodology, investigation, copy editing, and supervision; M.C.G. was involved in conceptualization, methodology, formal analysis, investigation, and writing; J.A.B. was involved in writing, copy editing, supervision, and formal analysis; S.W.C. was involved in conceptualization, methodology, investigation, visualization, and copy editing; J.C.S.R. was involved in formal analysis; O.S. was involved in methodology, investigation, and validation; D.B.K. was involved in conceptualization, methodology, and copy editing; and J.R.L. was involved in conceptualization, methodology, resources, copy editing, supervision, and funding acquisition.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by a National Institutes of Health grant to J.R.L., [R01AR057076].
SUPPLEMENTARY MATERIAL
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.