
Abstract
Chronic granulomatous disease (CGD) is a rare congenital immunodeficiency characterized by a defect in nicotinamide adenine dinucleotide phosphate oxidase required for phagocytosis. Hematopoietic stem cell (HSC) transplantation is currently the only curative treatment, but it is ladened with morbidities and mortality. Gene therapy is a promising treatment for CGD. However, if not properly designed, the gene therapy approach may not be successful. We engineered lentiviral vectors (LVs) carrying a universal promoter (EF1a) and two myeloid-specific promoters (miR223 and CD68) to drive the expression of green fluorescence protein (GFP) or CYBB, one of the key defective genes causing CGD. Tissue-specific LV expression was investigated in vitro and in a CGD mouse model. We compared GFP expression in both myeloid differentiated and undifferentiated HSCs. The CGD mice were transplanted with LV-modified mouse HSCs to investigate expression of CYBB and restoration of reactive oxygen species. The LV promoters were further compared under low and high-transgenic conditions to assess safety and therapeutic efficacy. A pneumonia disease model based on pathogenic Staphylococcus aureus challenge was established to assess the survival rate and body weight change. All three promoters demonstrated ectopic CYBB expression in vitro and in vivo. The EF1a promoter showed the highest expression of GFP or CYBB in transduced cells, including HSCs without cytotoxicity, whereas the LV-miR223 showed the highest transgene delivery efficiency with high myeloid specificity. Importantly, under low-transgenic condition, only the LV-EF1a-CYBB showed high antibacterial activity in vivo.
INTRODUCTION
Chronic granulomatous disease (CGD) is a primary immunodeficiency caused by the loss of function of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase in phagocytes. 1 NADPH oxidase consists of five subunits, and the loss of function of any subunit can cause CGD. Most patients (∼65%) carry a mutation in the CYBB gene on the X chromosome, while the others inherit an autosomal recessive mutation. 2 –6 Due to the NADPH oxidase deficiency, CGD patients cannot produce reactive oxygen species (ROS) to exert bactericidal effects, and infections occur repeatedly in various parts of the body. 5,6
Hematopoietic stem cell transplantation (HSCT) is a curative treatment for CGD and it requires myeloablative conditioning and an HLA-matched donor. Finding a suitable donor is difficult, and HSCT is associated with the risk of graft-versus-host disease and death. 7 –9 Furthermore, the success rate of HSCT in CGD patients is lower than in other primary immunodeficiencies. 7,8
Gene therapy is a promising alternative treatment. Clinical trials using gamma-retroviral vectors (RVs) have reported that the gene-modified cells do not persist in patients. 10 The RVs preferentially integrate near the promoter regions of pro-oncogenic loci and often display transgene silencing, a profile of poor safety and lack of persistence. On the other hand, lentiviral vector (LV)-mediated gene therapy has been successfully applied in treating monogenic diseases such as X-linked severe combined immunodeficiency and sickle cell disease. 10 –14 The safety and efficacy profile of the LV system has been largely proven in human trials. 15 –26
The ROS production can be restored in gene-modified CGD HSCs by ectopic expression of NADPH oxidase using non-tissue-specific promoters; however, overexpression of ROS may perturb the balance of quiescence, self-renewal, proliferation, and differentiation of HSCs. 27 Excessive ROS could lead to a significant exhaustion of stem cell pool by apoptosis or differentiation. 28 –32 The latter may be the reason for poor persistence of universal promoter transgene-mediated HSCT in previous trials.
LVs have been used to express gp91phox based on universal and chimeric myeloid-specific promoters without exemplified genotoxicity. 29,33 –38 CGD gene therapy using RV based on a strong spleen focus-forming virus promoter has reported the transactivation of the EVI1 proto-oncogene resulting in myeloid dysplasia syndrome. 39 Therefore, myeloid-specific promoters such as a miR223 promoter, a minimal gp91phox synthetic promoter, CD68 promoter, and CD11b promoter have been tested in CGD gene therapy. 33,40 –46 Brendel et al have reported restored E. coli killing activity in granulocytes in CGD mice using a miR223 promoter. 40
In a clinical trial, Kohn et al have investigated an LV chimeric cathepsin G and Cfes gene promoter/enhancer and reported stable vector copy number (VCN) (0.4–1.8 copies/neutrophil) and persistent oxidase-positive neutrophils (16–46%) in six of nine X-CGD patients after 12 months. 47 In this study, we compared a universal (EF1a) and two myeloid-specific promoters (miR223 and CD68) under low- and high-transgenic conditions both in vitro and in a CGD mouse model, and demonstrated restored ROS activities and in vivo control of pulmonary infection and inflammation after Staphylococcus aureus challenges.
MATERIALS AND METHODS
LV preparation
LVs were constructed, packaged, and concentrated using the NHP/TYF LV system as described previously. 48,49 The green fluorescence protein (GFP) gene mWasabi was a gift of Dr. Jiwu Wang (Allele Biotech). The mWasabi gene and the CYBB cDNA were cloned into pTYF behind the specific promoters and packaged in 293T cells as described. 50,51
Cell cultures
The HL60, HeLa, and EA-hy926 cell lines were obtained from ATCC (Manassas) and cultured in RPMI and DMEM (Hyclone, China), containing 10% FBS, and 1% penicillin and streptomycin. All cells were cultured at 37°C in 5% CO2.
LV transduction
HL60 were transduced with LV at MOI 50, and centrifuged at 100 g for 100 min. EA-hy926 and HeLa cells were transduced with LV at MOI 100. Transduction was performed with polybrene at 8 μg/mL (Sigma-Aldrich; Merck, Inc., China).
Determination of VCN
The LV VCN was determined using genomic DNA of transduced cells and standardized human and mouse reference cells based on SYBR green real-time quantitative PCR (qPCR) as previously described (Supplementary Materials and Supplementary Table S1). 52
Animal procedures
All mouse protocols were reviewed and approved by Institutional Animal Case and Use Committee (IACUC) of Shenzhen Geno-Immune Medical Institute (GIMI, Shenzhen, China). The X-CGD mice were purchased from Jackson Laboratory (X-CGD; B6.129S-Cybb tm1Din/J). Before HSCT, the 6–7-week-old male X-CGD mice were conditioned with 600 cGy using an X-ray irradiator (Faxitron).
HSCT and transplantation
Bone marrow was extracted from 12- to 16-week-old mice. HSCs were isolated using an EasySep™ mouse HSC isolation kit, and cultured in StemSpan-SFEM medium (Stemcell, China). The mouse cytokines were purchased from Peprotech, including SCF 50 ng/mL; Flt3-L 50 ng/mL; IL-6 10 ng/mL; and TPO 50 ng/mL. The HSCs were transduced with LVs at MOI 200. After transduction, the HSCs were infused through tail vein 4 days after irradiation.
Western blot analysis
Cells were collected and treated with RIPA buffer (Vazyme, China), and supplemented with a protease inhibitor (Boster Biological Technology). After electrophoresis and transfer, the membrane was blocked with 5% BSA (Biofroxx, Germany) for 1 h, and incubated with primary antibodies (anti-NOX2, Abcam; anti-GAPDH, Merck) at 4°C overnight. The membranes were incubated with horseradish peroxidase-conjugated secondary antibody (Cell Signaling Technology) for 1 h. The signals were detected with a chemiluminescence kit (ECL; Bio-Rad) and exposed and analyzed under the ChemiDoc Touch imaging system.
Dihydrorhodamine 123 assay
Blood cells were stimulated with phorbol-12-myristate-13-acetate (PMA, Sigma) and stained with Dihydrorhodamine 123 (DHR123; Thermo Fisher Scientific, China) as described. 29
S. aureus challenge
S. aureus was pelleted and prepared as inoculum of 2 × 1010 CFU/mL. The mouse was inoculated intranasally with 5 × 108 CFU twice within 30 min, 3 months after gene therapy. 53,54 The body weight, death time, and CFU counting in the lungs were recorded as described. 53,55,56
Cell preparation and flow cytometry
Cells were permeabilized using 0.1% Triton X-100 in PBS for 3 min, incubated with rat serum for 30 min, and stained with anti-NOX2 antibody (ab80508; Abcam) at 4°C for 30 min. A secondary antibody conjugated with AlexaFluor-488 (Invitrogen, Thermo Fisher) was added for 30 min. Analyses were performed using a NovoCyte Quanteon flow cytometer (ACEA Biosciences) and data were analyzed using the ACEA NovoExpress software.
Neutrophil extracellular trap induction
A total 5 × 105 cells were seeded in 24-well plates for 60 min. The cells were stimulated with 500 ng/mL PMA (Sigma-Aldrich) and incubated at 37°C for 3 h. The cells were fixed with 600 μL 4% PFA for 15 min. After washing with PBS, the cells were counterstained with DAPI (Thermo Fisher) at 200 μg/mL. The neutrophil extracellular trap (NET) images were captured with Applied Imaging Software CytoVision.
Statistical analysis
Statistical analysis was performed based on the Wilcoxon matched-pairs signed-rank test using GraphPad Prism 8 software (GraphPad, Inc., La Jolla, CA). All data were presented as mean ± standard error of the mean. Significance of difference between groups was evaluated using Student's t-test or analysis of variance one-way test (Tukey), and specified as **p < 0.05; ***p < 0.03; and ****p < 0.01; n.s., no significant difference.
RESULTS
LV construction and packaging analysis
We constructed LVs carrying a universal (EF1a) or modified myeloid-specific, miR223 and CD68, promoters to drive the expression of GFP or CYBB. 48,49 The LV-miR223-CYBB was packaged at a titer of 5 × 109 IU/mL, about 1.56- and 1.89-fold higher efficiencies compared with LV-CD68-CYBB (p = 0.032) and LV-EF1a-CYBB (p = 0.029), respectively (Supplementary Fig. S1).
Comparison of universal versus myeloid-specific promoters in vitro
We first examined GFP reporter expression in mouse HSCs and in three human cell lines, EA-hy926 (endothelial), HL60 (myeloid), and HeLa (epithelial) cells. The cells were transduced with LVs at higher than 1 vector copy per cell as determined by qPCR (Supplementary Fig. S2). On day 5 after LV transduction, the HSCs were induced for myeloid differentiation with mG-CSF (25 ng/mL). The percentage of GFP-positive cells was determined after 14 days. We confirmed that the differentiated cells were CD11b positive (myeloid) and there was no detrimental effect on the HSC differentiation after LV-CYBB gene transfer (Supplementary Fig. S3A). The GFP signals in differentiated versus undifferentiated mHSCs were as following: EF1a 84.72% versus 85.35%, miR223 89.16% versus 26.42%, and CD68 77.49% versus 58.1% (Fig. 1A).
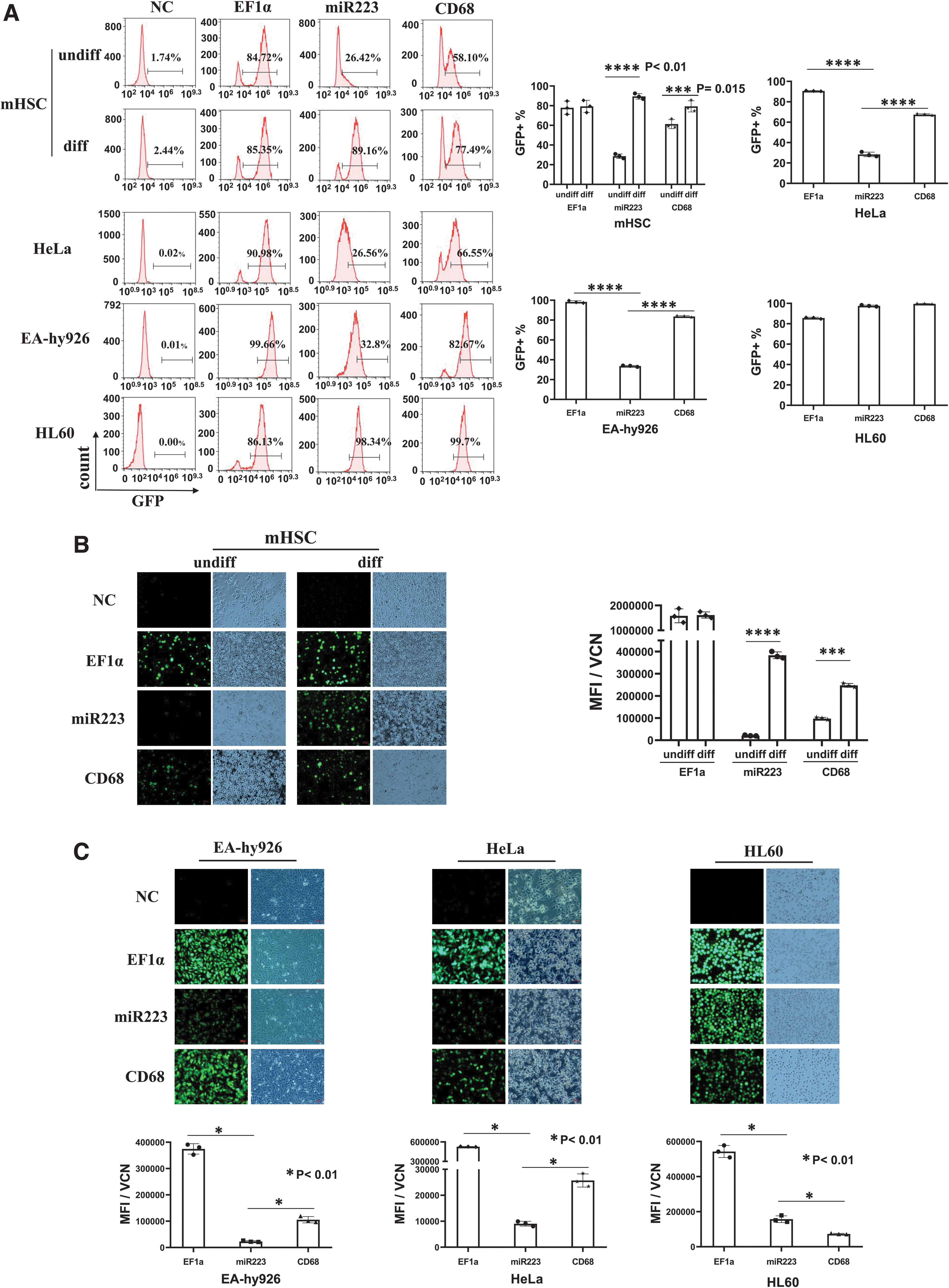
Analyses of LV reporter gene expression under universal (EF1a) or myeloid-specific promoters in mouse HSCs and in three human cell lines.
The miR223 promoter displayed significant myeloid specificity in mHSCs, and all three promoters showed high activities in HL60 by FACS (Fig. 1A). Based on MFI under equal VCN (MFI/VCN), the EF1a promoter showed similar levels of expression in both differentiated and undifferentiated cells, but for the two myeloid promoters, the differentiated cells showed increased expression, with the miR223 promoter displaying the highest myeloid specificity (miR223 18.56-fold increase, p = 0.0001, ****p < 0.01; and CD68 2.53-fold increase, p = 0.001, ***p < 0.03, Fig. 1B). The miR223 promoter displayed low GFP expression in HeLa and EA-hy926 cells, and all three promoters showed high activities in HL60 cells (Fig. 1C).
Ex vivo LV-CYBB expression and ROS analysis in differentiated CGD mouse HSCs
To examine LV-CYBB expression and functional restoration of ROS production, we performed ex vivo analyses using cells from wild-type (WT) and CGD mice. The HSCs were transduced with LV-CYBB to obtain VCN >1 copy/cell, and on day 5, the cells were induced for myeloid differentiation. After 14 days, the gp91-phox was detected by intracellular antibody staining and flow cytometry. After differentiation, gp91-phox was detected in CGD cells at 1.14%, in WT cells at 78.58%, and in CGD cells transduced with LV-CYBB carrying LV-EF1a at 72.38%, LV-miR223 at 75.4%, and LV-CD68 at 71.29% (Fig. 2A). Western blot analysis confirmed increased gp91-phox in cells transduced with LV-miR223 and LV-CD68 after myeloid differentiation; in contrast, the LV-EF1a showed no difference between undifferentiated and differentiated cells (Fig. 2B).

In vitro analyses of gp91-phox expression and ROS activity in differentiated CGD HSCs.
Next, we performed ex vivo functional assays using LV-CYBB-transduced CGD mHSCs. The DHR123 assay demonstrated restored ROS production near WT levels for all three promoters after LV-CYBB transduction as shown in Fig. 2C. We further performed the NET assay, which reflected ROS generation. Figure 2D showed that LV-CYBB-transduced CGD cells displayed markedly increased NET signals compared with the unmodified CGD cells. The LV-modified cells displayed markedly increased MFI of NETs after PMA treatment compared with control cells (Fig. 2D).
Functional analyses of LV-CYBB-modified HSCs in X-CGD mice
To investigate the different LV promoter activities, we performed HSCT of CGD mice by tail vein infusion of 1.5 × 106 LV-CYBB-transduced CGD HSCs after non-myeloablative radiation (n = 5, Fig. 3A). The expression and function of LV-CYBB were analyzed by gp91-phox staining and DHR123 assay 4 weeks post-transplantation. The blood cells were stained with anti-gp91-phox antibody together with antibodies for different lineage-specific markers, including T cells (CD3), B cells (B220), granulocytes (CD11b), and monocytes (Ly-6c).
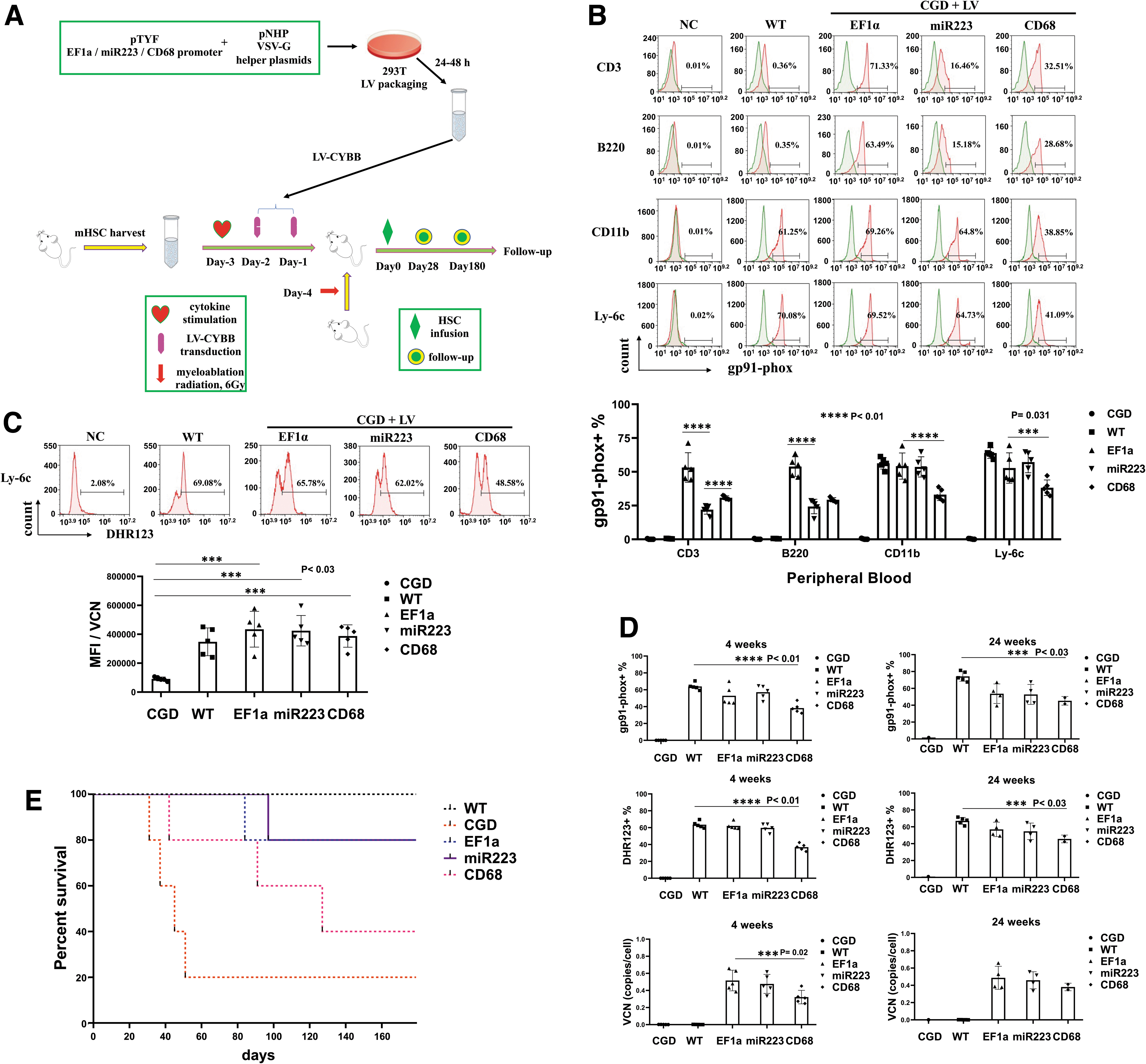
Functional analysis of LV-CYBB-modified HSCs in X-CGD mice.
Results showed that the EF1a promoter exhibited widespread expression of gp91-phox in all lineages, whereas the expression varied for miR223 and CD68 promoters, with the highest expression in granulocytes and monocytes, with the miR223 promoter (Fig. 3B). The EF1a promoter showed high activities in all cells, but miR223 and CD68 promoters displayed myeloid specificity (Fig. 3B). DHR123 analysis of Ly-6c-positive monocytes illustrated restored ROS activities close to WT levels in the CGD mice after LV-CYBB gene transfer for all three promoters (Fig. 3C), which was above the reported 20% ROS levels to prevent infections. 57 Further analyses for the gp91-phox expression and DHR123 assay indicated no significant difference between the WT mice and the LV-EF1a- or LV-miR223-treated CGD mice (p = 0.53), but a significant difference was observed with the LV-CD68-treated mice (p = 0.02, ***p < 0.05, Fig. 3D).
In continued follow-up, we found steady-state VCNs for all LV constructs 4 and 24 weeks after transplantation (VCN per cell, Fig. 3D). Survival analysis was performed after 160 days, and the results showed 20% survival for the untreated CGD mice, 40% for the CD68 promoter-treated group, and 80% for the EF1a and miR223 groups (Fig. 3E). We found that the dead mice, not the survival mice, showed extensive skin lesions due to the high-dose radiation. Evidently, fighting among the male mice exacerbated their injuries, which might be the cause of death of few LV-EF1a- and LV-miR223-treated mice.
Assessment of low versus high LV gene transfer conditions in correcting CGD phenotype
The gene transfer efficiency is a key factor to the success of gene therapy applications. 8,10,47,57,58 As gene transfer efficiencies might be different for different promoters, we designed an experiment to test the effects on CGD phenotype correction based on low versus high LV transgene levels, and analyzed the ROS recovery ability and resistance to pulmonary infection. We set the low functional transgenic threshold at 5% based on previous report that the DHR123+ cells below 10% are highly associated with infections. 57
Low-transgenic CGD mice based on 7–10% LV-modified HSCs were established using the three promoters (EF1a, miR223, and CD68, n = 10). The ROS recovery was confirmed at levels less than 10% after 3 months. For comparison, high-transgenic CGD mice were also established (n = 5). To assess for resistance to infection, each mouse was intranasally inoculated with 5 × 108 CFU S. aureus 3 months after gene therapy as illustrated in Fig. 4A. 53,54,56
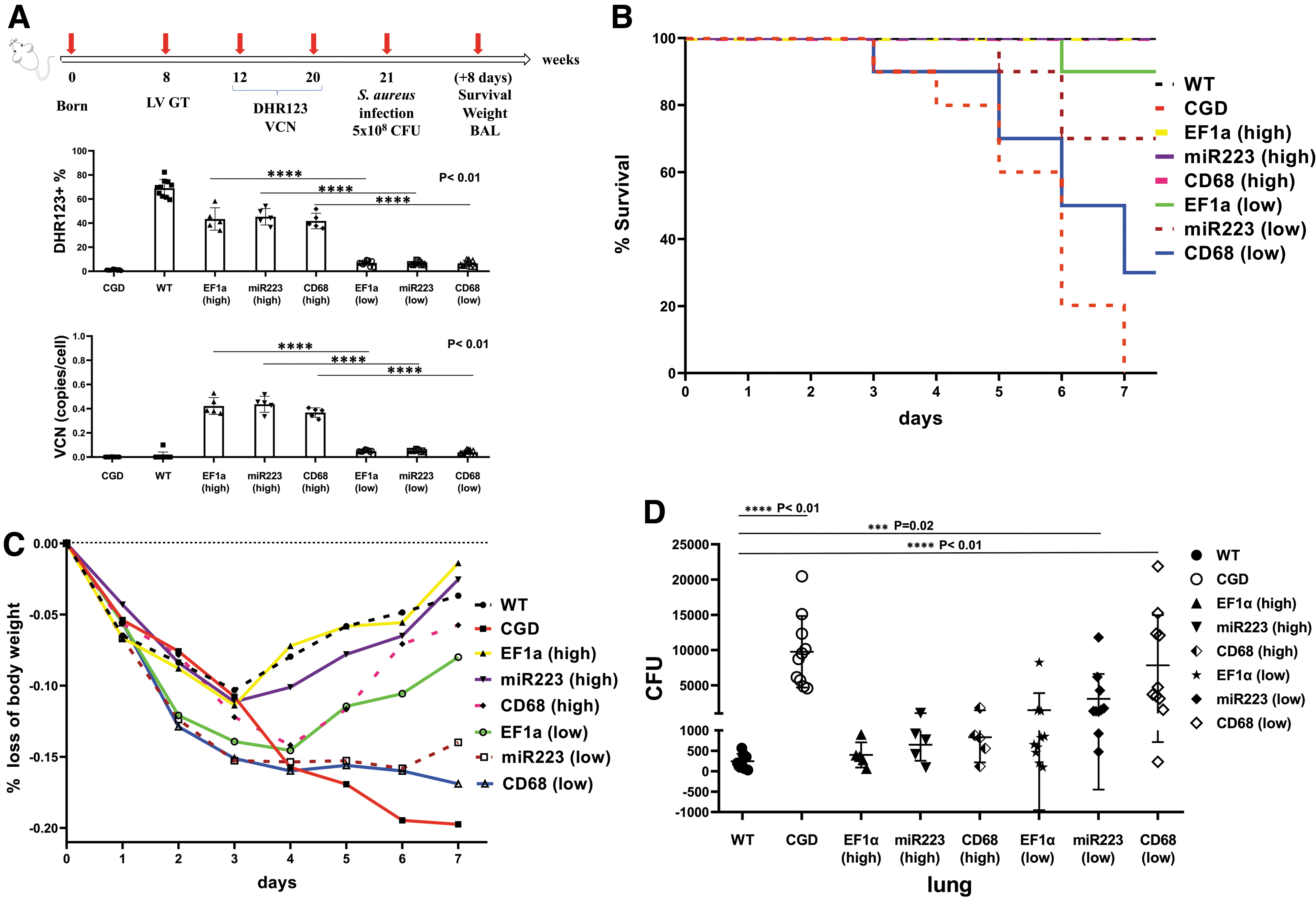
Assessment of LV promoter activities under low- and high-transgenic conditions in CGD mice.
The results showed that high-transgenic CGD mice were resistant to S. aureus challenge in 7–8 days, with an overall weight loss below 15%, similar to the challenged WT mice. The maximal weight loss in the high-transgenic EF1a and miR223 groups, and in the WT mice, was around 10% after 3 to 4 days, and the maximal weight loss in the high-transgenic CD68 group approached 15% on day 4 (Supplementary Table S2). Some of the low-transgenic CGD mice died on days 3 to 7.
On day 7, the low-transgenic EF1a, miR223, and CD68 displayed 10%, 33.3%, and 70% mortality, respectively (Fig. 4B). The maximal weight loss in the low-transgenic group approached ∼15% on day 4. Nevertheless, the low-transgenic groups began to gain weight on day 5; the miR223 low-transgenic mice regained weight slowly, but gradually, whereas the CD68 low-transgenic mice lost more than 15% of body weight, which reached the threshold to death (Fig. 4C). Thus, the high-transgenic condition helped restore the antibacterial ability in CGD mice, as quantified in the lungs by S. aureus CFUs (Fig. 4D), and the recovery of ROS production under the two tissue-specific promoters was slower compared with the universal EF1a promoter after gene therapy.
DISCUSSION
Previous gene therapy trials for CGD have reported detection of DHR123+ granulocytes from 0.004% to 0.05% in 3–6 weeks; however, no gene-corrected granulocyte was present after 1 year. 8,59 Several RV-mediated gene therapy trials after fludarabine and busulfan conditioning achieved long-term gene marking of only 0.08–0.5% of neutrophils. 10 In later trials, insertional activation of proto-oncogenes in HSCs has raised serious concerns with the RV. 39,40 Another study based on LV gene therapy has reported high initial VCN and DHR123+ neutrophils, but the latter decreased in some patients over time. 47 Overall, the engraftment of LV-modified cells was poor in pediatric patients, and the reason was unclear.
The X-linked CGD female carriers also experience CGD-related infections due to lionization. These carriers have median DHR123+ level ∼8% (from 0.06% to 48%). 8,57,58 Statistics indicate that the percentage of DHR123+ granulocytes may drop to very low levels in CGD patients and carriers. The premise of obtaining long-term and stable gene therapy efficacy in CGD may lie in the selection of an appropriate gene delivery vector, including novel promoter designs, to maintain a relatively high percentage of DHR123+ neutrophils.
In this report, we evaluated the properties of different LV promoters under high and low functional transgenic conditions. The hsa-miR223 gene and CD68 gene have been reported to be highly myeloid specific.40,60 –63 These two tissue-specific promoters could effectively limit the range of gene expression in targeted cells and thus reduce possible HSC toxicity. We observed that the universal EF1a and the myeloid miR223 and CD68 promoters could effectively drive the expression of GFP and CYBB in relevant cell types, and the miR223 promoter displayed a higher myeloid specificity than the CD68 promoter. The EF1a promoter was much stronger than the two tissue-specific promoters in driving transgene expression in all cell types.
All three LV promoters expressed CYBB and restored ROS production in in vivo analyses, and we observed >40% of ROS+ expression in 4 weeks. Doering et al. have reported that an LV CD68 promoter restored ∼20% of normal level of human F8 clotting factor in mice in 15 weeks. 63 We detected high gp91-phox and ROS 24 weeks after gene therapy similar to those reported by Brendel et al and Hernandez et al. 26,33,34 Nevertheless, the CGD mice treated with the LV-CD68 promoter started to die after 6 weeks, conceivably due to insufficient gp91-phox and ROS activities.
It has been reported that the engraftment of LV gene-marked cells is poor in pediatric CGD patients 47 ; thus, it is particularly important to investigate the effect of different promoters in controlling infections under low-transgenic conditions. Earlier studies have indicated that below 10% of DHR123+ neutrophils is highly associated with infections. 57 Therefore, we defined the low-transgenic threshold at around 5% to compare the different promoter activities in a pneumonia model. All three promoters restored the antibacterial activities at high-transgenic condition in the CGD mice.
Under low-transgenic condition, the EF1a universal promoter exhibited the best antibacterial activities, and the miR223 promoter performed better than the CD68 promoter. This direct comparison demonstrated that the universal EF1a promoter generated high levels of ROS that killed bacteria under a low-transgenic condition. Furthermore, the LV-EF1a-CYBB showed no evident adverse effect on the differentiation ability of HSCs, and we did not observe any toxicity in the LV-CYBB-treated CGD mice in 66 weeks.
In conclusion, our study suggests that under low-transgenic condition, the strong universal EF1a promoter is a better choice than the two myeloid-specific promoters. However, at high-transgenic condition, the miR223 promoter was as effective as the EF1a promoter in CGD phenotype correction. Thus, it may be necessary to restore the function of both myeloid and nonmyeloid immune effector cells to obtain optimal effects.
Footnotes
ACKNOWLEDGMENTS
We thank Jie Gong for caring for the mice, Yue Qin and Yuchen Liu for the LV preparation, Liu Xu for qPCR analysis, Dr. Ruiqin Fang and Dr. Lixia Tang for the kind gift of bacterial strains, and Dr. Biljana Horn and Dr. Yoshiyuki Takahashi for reading and advice on the article.
AUTHORs' CONTRIBUTIONS
Engineered the LV constructs and finalized the article: L.J.C. Designed the study: L.J.C. and H.Y. Performed the laboratory work: H.Y., X.W., and H.L. Performed the data analysis: H.Y. and L.J.C. Drafted and revised the article: H.Y. and L.J.C. All authors read and approved the final article.
DATA AND MATERIALS AVAILABILITY
All raw data used for figure generation in this study can be accessed by contacting the corresponding author.
AUTHOR DISCLOSURE
The authors declare that they have no financial conflict of interest.
FUNDING INFORMATION
This work was supported by grants from the Foundation for Introduction and Training of Outstanding Scholarship, The National “985” Project (A1098531023601102); and The Fundamental Research Funds for the Central Universities (ZYGX2016Z009).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.